
谷氨酸棒杆菌中基于5′UTR及其下游序列的增强型表达载体构建
2021-10-25 08:53方求武孙曼曼刘秀霞杨艳坤白仲虎
生物学杂志 2021年5期
方求武,高 雄,孙曼曼,刘秀霞,李 业,杨艳坤,白仲虎
(1.江南大学 粮食发酵工艺与技术国家工程实验室,无锡 214122;2.江南大学 工业生物技术教育部重点实验室,无锡 214122;3.江南大学 生物工程学院,无锡 214122)
谷氨酸棒杆菌(Corynebacteriumglutamicum)是革兰氏阳性细菌,广泛用于氨基酸发酵,如谷氨酸[1]和赖氨酸[2]的生产。近年来,由于其易于分泌目标蛋白[3],细胞外水解酶活低和无内毒素[4]等特性,已发展成为表达外源蛋白的理想宿主[5]。然而,高强度表达载体的缺乏限制了谷氨酸棒状杆菌的应用。因此,探索和开发一些具有高表达能力的载体具有重要意义,也在一定程度上促进谷氨酸棒杆菌的商业应用。
通过筛选具有高表达能力的启动子和优化核糖体结合位点(ribosome binding site,RBS)序列是目前提高谷氨酸棒杆菌蛋白表达的主要方法。其中,Ptac启动子和乳糖操纵子结合被广泛应用于谷氨酸棒杆菌的各种载体表达[6]。在此基础上,PtacM启动子的修饰会进一步增强蛋白的表达[7]。此外,谷氨酸棒杆菌的几种天然启动子,包括组成型启动子Ptuf、PcspB和Psod和诱导型启动子PprpD2已被应用于工业蛋白的表达[6-9]。另外几种合成启动子PH36、PI16和PL26也被应用于构建蛋白表达载体[10-13]。RBS序列也称SD序列,我们前期研究证实在谷氨酸棒杆菌中Paph启动子与不同RBS序列结合后呈现出不同的蛋白表达能力[14]。作为基因表达的重要部分,RBS序列的优化也已应用于谷氨酸棒杆菌的代谢工程。通过构建RBS文库和调节信号通路基因aroG、aroB、aroD和aroE的RBS强度,建立了几个遗传模块来提高谷氨酸棒杆菌中草酸的产量[15]。实际上,启动子下游的一小段转录非编码区(5′UTR)不仅包含更多的RBS序列,而且还包含可能影响启动子活性的特殊序列[16]。与仅有几个碱基的RBS相比,5′UTR包含大量特殊序列,使其具有更高的修饰潜力。
本实验室的前期工作中已经证明了在谷氨酸棒杆菌中,与单顺反子载体(monocistronic expression model,MEM)相比双顺反子表达载体(bicistronic expression model,BEM)能够显著提高载体的表达能力[5]。研究利用不同的5′UTR及其下游序列与不同启动子组成新的表达组件,实现强表达载体筛选,对扩展谷氨酸棒杆菌的工业应用具有重要意义。
1 材料与方法
1.1 菌株、生长条件和质粒
大肠杆菌DH5α和谷氨酸棒杆菌CGMCC1.15647由本实验室保存。大肠杆菌DH5α用于构建表达载体;谷氨酸棒杆菌作为外源蛋白表达宿主,同时为5′UTR序列的PCR提供基因组模板。氯霉素的终浓度在大肠杆菌中为30 mg/L,在谷氨酸棒杆菌中为10 mg/L。VHH蛋白基因及分泌信号肽T1514片段从本实验室保存的p19-T1514-VHH载体扩增得到。pH36-EGFP是基于本实验室保藏的pE-0载体构建携带PH36启动子和报告基因EGFP的质粒,pTac-EGFP是基于pXMJ19构建携带Ptac启动子和EGFP报告基因EGFP的质粒[17]。

表1 5′RACE所用的引物Table 1 Primers in 5′RACE experiment
1.2 cDNA 5′末端快速扩增实验(5′RACE)
cDNA末端快速扩增实验(rapid amplification of cDNA ends,RACE)是基于PCR从低丰度转录本中通过快速扩增获得cDNA 完整5′和3′末端的有效方法。根据谷氨酸棒杆菌中蛋白质表达的丰度,列出了12个基因[18](表2),并且使用5′RACE分析法鉴定了不同基因的5′UTR序列及其下游部分序列。如图1(a)所示,在谷氨酸棒状杆菌过夜培养后,收集处于对数生长期的细胞,并利用细菌总RNA提取试剂盒(TakaRa,中国)提取谷氨酸棒状杆菌的总RNA。然后使用基因特异性引物(gen-specific primer,GSP1)在有逆转录酶(GoldenstarTM RT6 cDNA Synthesis Kit)的作用下使基因片段内部合成cDNA的第一条链,同时用RNAse混合物降解模板mRNA,并纯化合成cDNA,通过末端转移酶将dCTP或dGTP连续添加到cDNA链的3′末端,以形成寡核苷酸(dG或dC)尾巴。最后使用基因片段内部寡聚锚定引物(anchor primer,AP)和另一种基因特异性引物(GSP2)进行巢氏PCR扩增,得到PCR产物进行测序,测序得到连续寡核苷酸(dG或dC)尾巴下游即为转录起始位点(TSP),通过与序列库比对以获得5′UTR及其基因的下游部分序列。
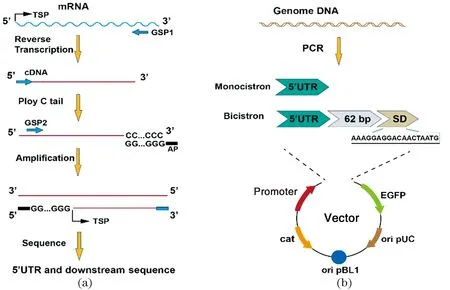
TSP:转录起始位点。图1 利用5′RACE鉴定5′UTR及其下游序列(a);5′UTR及其下游序列与启动子组成新的表达载体的构建(b)Figure 1 Identification of 5′UTR and its downstream sequence by 5′RACE(a);construction new expression vectors by combining promoter with different 5′UTR and its downstream sequence(b)
1.3 5′UTR表达载体的构建和插入
如图1(b)所示,在单顺反子表达模型MEM中,将从基因组中克隆的5′UTR利用同源重组的方式插入到pH36-EGFP载体中。由不同5′UTR序列和H36启动子组成的新表达载体根据基因名分别命名为pH36-M2673/0935/2501/2473/2328/2826/0572/0533/1316UTR-EGFP。而在双顺反子表达模型BEM中,将不同基因的5′UTR及其下游62 bp和保守SD(AAAGGAGGACAACTAATG)序列分别重组到pH36-EGFP及pXMJ19-EGFP载体上,构成新的双顺反子表达载体,分别命名为pH36-B2673/0935/2501/2473/2328/2826/0572/0533/1316UTR-EGFP和pTacB2673/0935/2501/2473/2328/2826/0572/0533/1316UTR-EGFP。pH36-EGFP和pTac-EGFP分别作为阴性对照pH36-Negative和pTac-Negative,而将保守SD序列插入pH36-EGFP和pTac-EGFP分别为阳性对照载体pH36-Positive和pTac-Positive。
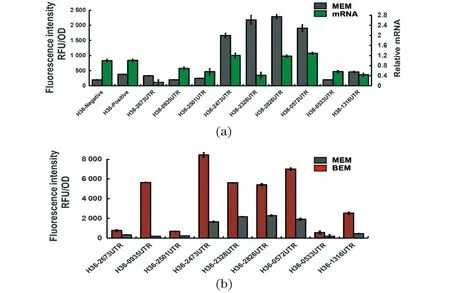
图 2 5′UTR序列与PH36启动子组成新的单顺反子载体的荧光强度以及5′UTR对启动子转录水平的影响(a);5′UTR及其下游序列与PH36启动子组成的双顺反子载体egfp蛋白的表达(b)Figure 2 Fluorescence intensity of monocistronic expression element consisting of PH36 promotor and 5′UTR and relative mRNA of EGFP gene for each part(a);egfp protein expression of different expression elements in BEM compared with MEM(b)
1.4 荧光强度测定
用报告蛋白EGFP检测不同5′UTR的表达强度。将成功构建的具有不同5′UTR表达载体转化至谷氨酸棒状杆菌中,分别从平板上选取3个转化子将其转染至24孔板中(每孔含2 mL LBB培养基)。30 ℃以220 r/min培养约12 h后,将谷氨酸棒杆菌转接至新的24孔板(每孔含2 mL LBB培养基)中,初始OD600为0.05,二次活化培养3 h后,在含有诱导型启动子tac的每个孔中添加1 mmol/L诱导剂IPTG,添加IPTG继续培养24 h后测量荧光强度,组成型启动子PH36不需要添加诱导剂。
用培养基将酶标仪进行标零,取培养后的菌液加入培养基进行充分稀释,使稀释终浓度OD600为0.4~0.7,利用酶标仪测定菌液的OD600浓度并记录,同时利用荧光计测定稀释后菌液的荧光强度(激发波长:488 nm,发射波长:507 nm),计算单位OD600的荧光强度以指示不同5′UTR的表达强度。每种菌液取3次样测量作为技术重复,每个载体转化后选取3个转化子作为生物学重复。
1.5 实时定量PCR(qRT-PCR)
为验证5′UTR是否会影响启动子的转录强度,分析单顺反子载体中EGFP基因的mRNA转录水平。取摇瓶培养过夜菌液,如方法1.2所述,进行谷氨酸棒杆菌总RNA提取和cDNA合成。qRT-PCR程序为95 ℃预变性30 s;95 ℃变性5 s,64 ℃退火30 s,72 ℃延伸20 s;95 ℃变性15 s,65 ℃退火30 s和95 ℃持续退火15 s,以16S rRNA为标准,使用2-△△Ct方法分析相对EGFP的转录水平[19]。每种样品进行3次实验重复。
1.6 VHH表达载体的构建
为分析新表达载体在谷氨酸棒状杆菌中生产重组蛋白的实用性,选择3株高强度表达载体pTac-B2826/2328/2473UTR对重组蛋白VHH进行分泌表达。从p19-T1514-VHH载体中扩增出带有T1514信号肽的VHH基因片段,并利用重组的方法将其克隆到3个载体中,从而构建了pTac-B2826/2328/2473UTR-VHH 3种蛋白表达载体。并将6×His标签添加到VHH蛋白质的C末端,以简化目标蛋白质的检测和纯化。
1.7 VHH-His的纯化和分析
将带有表达效果最好的载体pTac-B2826-VHH的谷氨酸棒杆菌菌株在LBB培养基中于30 ℃过夜培养,然后以1∶50的比例转移到新鲜培养基中,培养3 h后当OD600为0.6时加入1 mmol/L诱导剂IPTG。培养24 h后,通过离心和过滤收集培养上清液。然后,用AKATa系统(GE Healthcare,Uppsala,瑞典)中的HisTrap HP色谱柱纯化VHH-His蛋白。纯化后蛋白质应用双辛可宁酸(BCA)、SDS-PAGE以及Western Bolt测定。
2 结果与分析
2.1 从高蛋白表达基因中筛选和鉴定5′UTR序列
在谷氨酸棒杆菌中筛选表达量最高的12个基因[18]。通过5′RACE实验,成功鉴定5′UTR及其基因下游的部分序列(表2)。这12个基因中有9个具有5′UTR序列,而其中3个基因启动子直接与基因起始密码子ATG连接而缺少5′UTR序列。
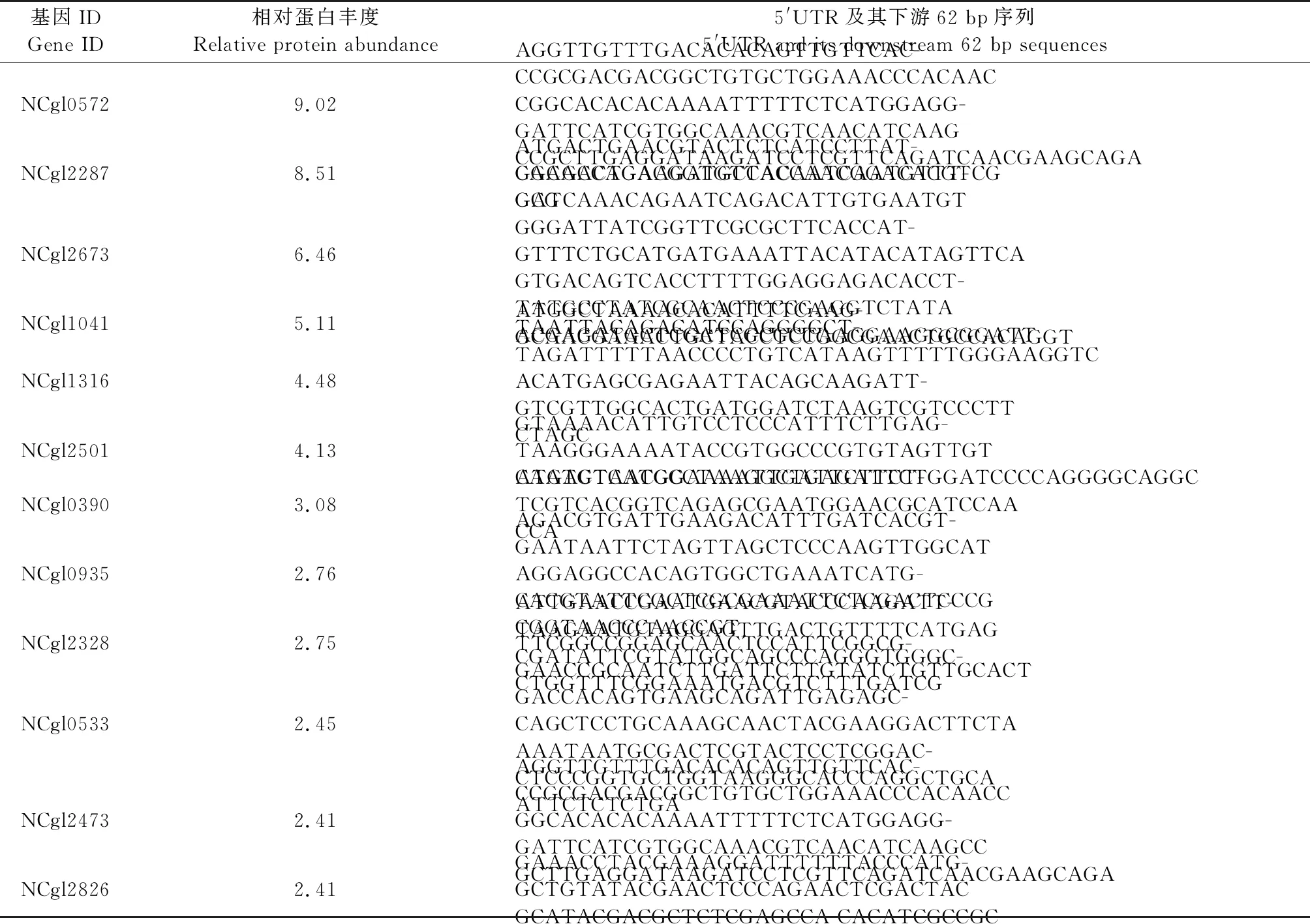
表2 在谷氨酸棒杆菌中筛选蛋白表达丰度最高的12个基因和其5′UTR及下游部分序列Table 2 Twelve genes with the highest protein abundance in Corynebacterium glutamicum and 5′UTR and its downstream sequences
2.2 不同5′UTR序列对蛋白表达的影响
首先测量单顺反子表达模型(MEM)中的荧光强度。如图2(a)所示,不同的5′UTR对蛋白表达水平的影响不同,然而单顺反子表达模型中仅含PH36启动子的表达元件并未显示出文献中的高表达强度[10],这可能是由于实验中菌株种类的不同造成的差异。然而,含有带有5′UTR序列的PH36的表达元件显著提高了egfp蛋白的表达,这证明内源表达元件5′UTR确实在谷氨酸棒杆菌中对蛋白的表达中起着至关重要的作用。与阳性对照pH36-Positive相比,具有不同5′UTR序列的表达载体实现了egfp蛋白的荧光强度从1.03倍增加到11.65倍。为进一步探讨5′UTR影响基因表达的方式,通过qRT-PCR分析,以检测表达过程中不同表达载体中EGFP基因的相对转录水平。这些结果表明,不同5′UTR序列能够显著影响着启动子的转录水平,这也说明5′UTR序列不仅影响蛋白的翻译效率,同时也对基因的转录有着重要影响。
2.3 5′UTR及其下游序列在双顺反子表达模型中的作用
双顺反子表达模型(BEM)中的荧光强度如图2(b)所示,与单顺反子模型相比,双顺反子模型中5′UTR与启动子PH36组成的表达载体的表达能力有了进一步提高,egfp蛋白的荧光强度分别提高了1.21~27.82倍。同样还测量5′UTR及其下游序列与Ptac启动子组合形成的表达载体对egfp蛋白表达的影响(图3),其中载体pTac-B2826UTR表达egfp蛋白的能力最高,其荧光强度比阳性对照pTac-Positive提高了 3.57倍。
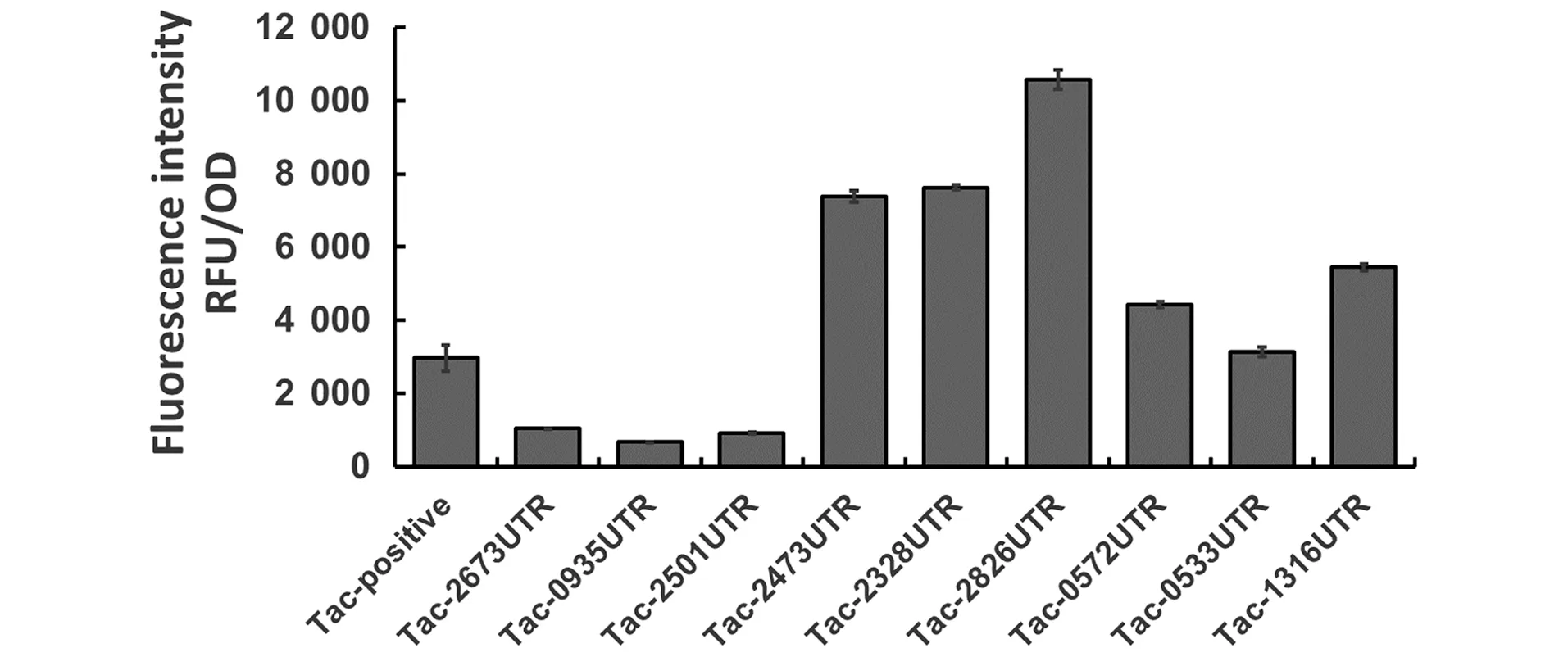
图3 5′UTR及其下游序列与Ptac启动子组成新的载体表达egfp蛋白的荧光强度Figure 3 Fluorescence intensity of egfp protein by new vectors constituted by 5′UTRs and their downstream sequences with Ptac promoter
2.4 重组蛋白VHH的分泌表达
为进一步证明筛选出的高表达载体对外源蛋白的表达能力,用VHH替换EGFP蛋白进行验证。选用3柱表达强度最高载体(pTac-B2826/2328/2473UTR)来表达蛋白VHH(15.7 ku),SDS-PAGE电泳实验结果[图4(a)]表明,这3株新型表达载体均能实现外源蛋白在谷氨酸棒杆菌中分泌表达。发酵结束后收集上清液,并成功通过镍柱纯化出具有His标签的VHH蛋白,如图4(b)。目标蛋白VHH下面杂带经WB分析,可能是由于SDS电泳时温度升高导致VHH蛋白出现部分降解所致。通过BCA蛋白浓度分析,在摇瓶中发酵24 h,VHH蛋白分泌浓度达到85.4 mg/L。
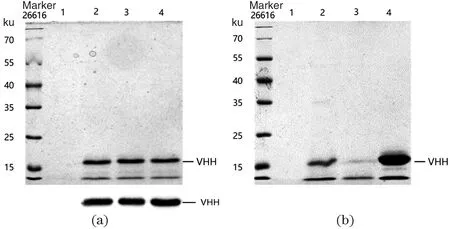
(a)泳道1为阴性对照(泳道2~4分别为pTac-B2826/2328/2473UTR-VHH载体表达VHH蛋白);(b)泳道1~4分别为阴性对照、原液、流穿液和洗脱液。图4 SDS-PAGE和WB分析VHH蛋白的表达(a)和纯化后VHH的SDS-PAGE分析(b)Figure 4 SDS-PAGE and WB analysis of VHH protein expression (a)and SDS-PAGE analysis of purified VHH(b)
3 结语
筛选谷氨酸棒杆菌中12个蛋白表达丰度最高的基因,通过5′RACE实验成功鉴定这12个基因的5′UTR及其下游序列,通过分子克隆技术,以EGFP作为报告基因验证5′UTR及其下游序列对不同的强启动子PH36和Ptac的表达能力均有显著的提升作用,并利用表达能力最强的载体成功在谷氨酸棒杆菌实现外源蛋白VHH的分泌表达。
猜你喜欢
华人时刊(2022年9期)2022-09-06
华人时刊(2020年15期)2020-12-14
甘肃教育(2020年2期)2020-09-11
甘肃教育(2020年4期)2020-09-11
作文成功之路·小学版(2020年7期)2020-08-24
甘肃教育(2020年20期)2020-04-13
中成药(2017年6期)2017-06-13
中国洗涤用品工业(2017年2期)2017-04-16
中国比较医学杂志(2017年5期)2017-01-17
医学研究杂志(2015年12期)2015-06-10
