
PVP处理对黑皮油松外植体酚类物质形成及酶活性的影响
2021-12-15 03:32赵雪莹刘德强
林业科学 2021年10期
梁 艳 赵雪莹 白 雪 刘德强 张 妍 潘 朋
(齐齐哈尔大学生命科学与农林学院 齐齐哈尔 161006)
黑皮油松(Pinustabulaeformisvar.mukdensis)为油松的变种,抗逆性强,具有耐干旱和瘠薄土壤、耐寒、防风、抑菌能力强等优良特性,是我国北方兼具经济价值与生态价值的造林树种(Wangetal., 2019)。黑皮油松树形俊美、枝虬若龙,尤其是冬态更能充分体现北方地域性的植物景观特色,已被广泛应用于我国北方园林绿化中(陈新宇等, 2019)。由于黑皮油松生命周期较长,且存在种子结实率低、子代优良性状降低等问题(Huietal., 2016),导致嫁接与种子的传统良种繁育方式难以满足市场需求,而离体快繁技术能显著加速树种品质改良并提高良种苗木的生产能力(高洁等, 2019; 宋跃朋等, 2019)。课题组前期研究发现,与其他木本植物类似(Bongaetal., 2010),以黑皮油松的休眠芽作为外植体进行离体培养褐变较严重,类似的褐变问题在欧洲赤松(Pinussylvestris)等松属(Pinus)植物的离体培养中也普遍存在(Andersoneetal., 2005),极大地影响了其离体培养的成功率,制约了其优质组培苗的快速繁殖与推广应用。因此如何有效抑制黑皮油松外植体褐变是组培生产中亟待解决的关键问题之一。
大量研究表明,培养基中添加聚乙烯吡咯烷酮(polyvinyl pyrrolidone,PVP)、维生素C(VC)、硝酸银(AgNO3)和活性炭(active carbon,AC)等抗氧化剂以及酚类物质的吸附剂可达到抑制外植体褐化的目的。在葡萄(Vitisvinifera)(饶慧云等, 2015)、天女木兰(Magnoliasieboldii)(高红兵等, 2017)、水曲柳(Fraxinusmandshurica)(彭春雪等, 2019)等材料的研究中发现,培养基中添加适宜浓度的PVP对抑制外植体褐化效果显著。组培中褐变多由酶促褐变引起,酚类化合物是褐变发生的主要底物,酚类物质再经多种氧化酶催化后形成醌类物质,醌类物质通过聚合产生褐色物质,酶、底物和氧气是引起外植体褐变的3个基本条件。酚类物质的含量与种类直接影响着褐化的程度,相关研究已发现与褐化有关的酚类物质主要包括咖啡酸、阿魏酸、香豆酸、绿原酸、儿茶酚和对羟基苯甲酸等(Babbaretal., 1986; Cosetengetal., 2010)。相对于草本植物,酚类物质含量较髙的木本植物的褐化问题尤为严重,成为制约其离体培养及快速繁殖成功与否的重要因素(Xuetal., 2011; Wangetal., 2014)。此外,多酚氧化酶(polyphenol oxidase,PPO)、过氧化物酶(peroxidase,POD)、苯丙氨酸解氨酶(phenylalanine ammonia-lyase,PAL)等也与褐化过程相关(Wangetal., 2014)。
目前黑皮油松愈伤组织抗褐化研究已有相关报道(付双彬等, 2017),但对于其休眠芽外植体离体培养中的褐化现象及抗褐化处理过程中相关酶活性与酚酸类物质含量变化的研究尚未见报道。鉴于此,本研究以成年黑皮油松的休眠芽为外植体,分析抗褐化剂PVP处理对其褐变及萌发的影响,筛选适宜的PVP处理浓度,在此基础上分析抗褐化过程酚酸类物质及PPO、POD、PAL酶活性的动态变化,确定引起褐化现象的主要酚酸类物质,探讨PVP处理过程中酚酸类物质及相关酶活性与其褐化的相关性,为黑皮油松组织培养过程中褐变这一瓶颈问题的解决提供理论依据和技术支撑,进而为黑皮油松高效植株再生体系的构建奠定基础。
1 材料和方法
1.1 试验材料和培养方法 2018年3月10日从齐齐哈尔大学校园的成年黑皮油松上采集生长健壮、无病虫害、芽体饱满的休眠叶芽作为外植体。叶芽在洗洁精溶液中清洗干净后流水下冲洗2 h,超净工作台上采用0.1%的HgCl2浸泡6 min,无菌水冲洗5次,每次5 min(带有振荡); 无菌器械剥去芽鱗(图1A),将叶芽横切成0.5~0.7 cm厚的薄片,按照形态学方向竖直接种到培养基上(图1B)。

图1 PVP处理对黑皮油松外植体褐化的影响
1.2 PVP处理试验 DCR基本培养基(Guptaetal., 1985)中分别添加不同质量浓度的PVP(100、300、500、800、1 000、1 500、2 000 mg·L-1),分别记为P1、P2、P3、P4、P5、P6、P7,以不添加任何抗褐化剂为对照(CK)。其余添加成分相同,分别添加30 g·L-1蔗糖、7 g·L-1琼脂、2.0 mg·L-16-BA,pH5.8。培养条件: 温度(25 ± 2)℃,光照每天16 h,光照强度1 500~2 000 lx。每个处理培养30个外植体,重复3次。接种后每隔4天观察外植体的褐化情况,培养32天统计外植体的褐化率及休眠芽萌发率。
采用目视观察确定褐化程度,将褐化等级划分为3级: “+” 外植体轻度褐化,周围培养基色泽浅; “++” 外植体中度褐化,周围培养基颜色加深; “+++” 外植体褐化严重,周围培养基颜色深。
褐化率=(褐化的外植体数 /接种的外植体总数)× 100%;
萌发率=(萌发的外植体数 /接种的外植体总数)× 100%。
1.3 酶活性和酚酸含量的测定 1)取样 分别于接种后0、4、8、12、16、20、24、28、32天选取1 500 mg·L-1PVP处理与CK处理下典型性的黑皮油松外植体,液氮速冻后-80 ℃保存用于酶活性与酚酸含量的测定。
2)酶活性的测定 PPO活性测定参照朱广廉等(1990)方法,PAL活性测定参照Solecka和Kacperska(2003)方法,POD活性测定参照李合生(2003)的愈创木酚法,3种方法均略作修改。
3)酚酸含量的测定 参考Alonso-Salces等(2005)方法略加改动。称取冷冻样品1.0 g,加入10 mL 50%甲醇水溶液,研磨制备匀浆液,50 ℃ 40 kHz超声40 min,重复提取2次,合并提取液; 将提取液8 000 r·min-1离心10 min,收集上清液,剩余上清液用旋转蒸发仪40 ℃减压浓缩,2 mL甲醇润洗残渣,50%甲醇定容至5 mL,上机测试。测试条件: 超高效液相色谱仪(型号 1290,美国安捷伦公司生产); 二极管阵列检测器(型号1290 Infinity II); 色谱柱为Eclipse Plus C18柱(50 mm × 2.1 mm,1.8 μm)。使用A和B双泵系统,流速0.5 mL·min-1,柱温30 ℃,波长280 nm,进样量1 μL。流动相A为2%(体积分数)乙酸水溶液,流动相B为甲醇溶液。采用梯度洗脱: 0~2 min,A100%~100%,B0~0; 2~3 min,A100%~95%,B0~5%; 3~4 min,A95%~85%,B5%~15%; 4~5 min,A85%~75%,B15%~25%; 5~7 min,A75%~65%,B25%~35%; 7~9 min,A65%~50%,B35%~50%; 9~12 min,A50%~100%,B50%~0。甲醇和乙酸均为色谱纯,没食子酸、绿原酸、咖啡酸、儿茶酚、香豆酸、对羟基苯甲酸和阿魏酸7种标准品纯度≥99%(Sigma公司生产)。定量方法参照孙洪圳等(2012),并略作改动。每个样品重复3次进样测定,测定结果取平均值。
1.4 数据统计与处理 试验数据采用Microsoft Excel 2007、SPSS 17.0、Origin 8.5软件进行分析处理,邓肯氏新复极差法(Duncan)进行显著性检验,图中数据均为3次重复平均值。
2 结果与分析
2.1 抗褐化剂PVP对黑皮油松休眠芽外植体褐化和萌发的影响 以未添加抗褐化剂的外植体为对照(CK),在培养基中添加不同质量浓度的抗褐化剂PVP,于处理32天统计外植体褐化率与萌发率(表1)。结果表明,随着PVP处理浓度的增加,褐化率与褐化程度均呈现先降后增的规律性变化,萌发率则先增后降,说明PVP处理可以有效抑制黑皮油松休眠芽外植体的褐化,并促进其萌发。当PVP浓度为1 500 mg·L-1(P6)时,褐化程度最轻,褐化率最低(25.56%),萌发率也最高(37.78%),整体来看显著优于其他处理; 当PVP增至2 000 mg·L-1时,其抑制褐化与促进萌发效果再次降低。综合来看,300~1 500 mg·L-1PVP处理可以有效抑制黑皮油松休眠芽外植体的褐化,其中1 500 mg·L-1效果最好。培养32天结束时,对照休眠芽由上至下整体变为深褐色,几乎全部死亡(图1C),而PVP处理的外植体褐化程度明显轻于对照组,长势也明显优于对照组(图1D)。
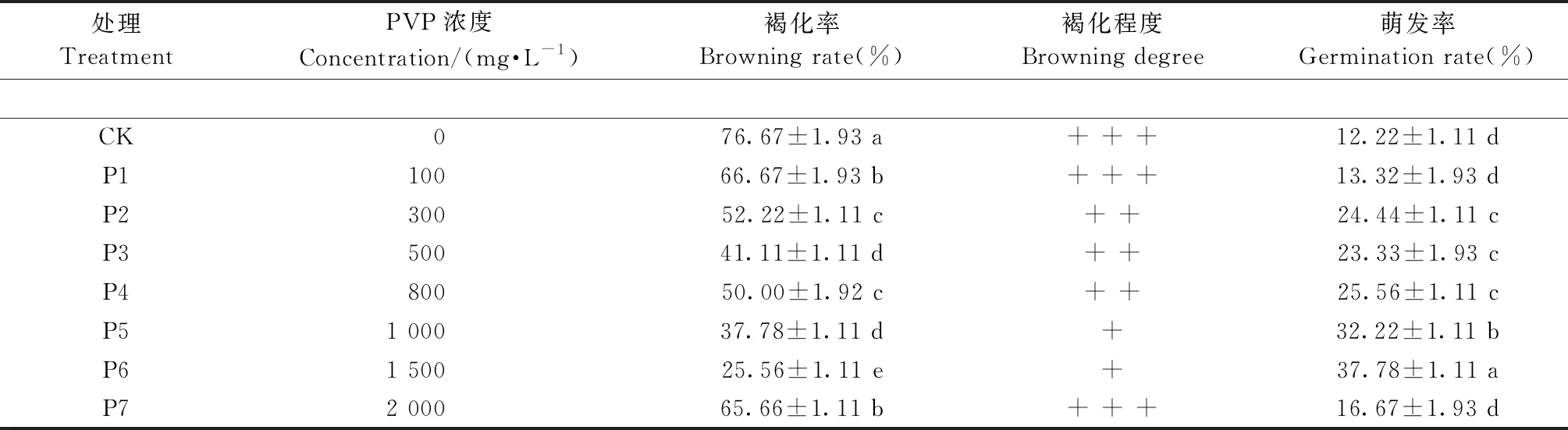
表1 不同浓度PVP处理对外植体褐化和芽萌动的影响①
为进一步明确PVP抑制黑皮油松外植体褐化的时间效应,对PVP处理不同时间节点外植体的褐化情况进行统计分析(图2)。随着处理时间的增加,对照与所有PVP处理外植体的褐化率均呈现先增加后趋于稳定的变化趋势。具体来看,外植体均在培养4天时开始出现褐化,随后随着培养时间的增加,褐变的外植体数量逐渐增加。培养8天时,除P7处理外,其余PVP处理的褐化率均显著低于对照,表明PVP(100~1 500 mg·L-1)处理从第8天开始表现出明显的抑制黑皮油松外植体褐化的作用; 8~12天所有处理外植体褐变愈发严重,褐化率增加幅度大,其中800 mg·L-1PVP处理的褐化率从14.44%急剧增加到47.78%; 除P2、P6处理外,对照与其他5个PVP处理的外植体均在20~24天时褐化率达到峰值,随后趋于稳定。
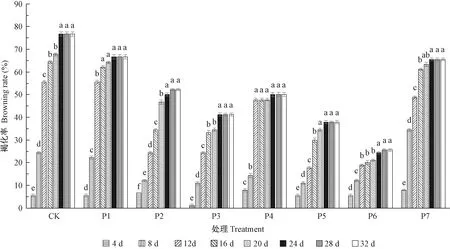
图2 PVP处理不同时间对黑皮油松外植体褐化的影响
2.2 PVP处理过程相关酶活性的动态变化 组培褐变是一个复杂的生理代谢过程,为了更清楚地了解抗褐化剂PVP减缓黑皮油松离体外植体褐化的机理,进一步以1 500 mg·L-1PVP处理(最佳处理)不同时间节点的黑皮油松外植体为材料,以不添加PVP的外植体为对照,分析PPO、POD、PAL 3种抗氧化酶活性的变化特征(图3)。
1) PPO活性的动态变化 随着培养时间的延长,对照组PPO活性呈现“升—降—升—降”的交替式变化,0~8天 PPO活性有小幅增加,8~12天急剧增加,12天出现峰值,活性为8.75 U·g-1min-1,显著高于其他处理天数(P< 0.05),为PVP同样处理天数下的3.5倍; 12~20天PPO活性呈现递减的变化趋势, 20~24天再次增加,24天活性为5.83 U·g-1min-1,随后再次降低。对照组PPO活性在8~12天和20~24天急剧增加,这2个时间点恰好是外植体褐变愈发严重的时期。与对照相比,PVP处理的外植体PPO活性在整个培养周期内始终处于相对较低的水平(1.25~2.92 U·g-1min-1),从处理后4 天开始PPO活性显著低于对照(图3A)。上述结果表明PPO活性变化与黑皮油松外植体褐化密切相关,PVP处理可抑制PPO活性的升高。
2) PAL活性的动态变化 在0~32天内,CK处理PAL活性呈现“升—降—升—降—升—降”的交替式变化,0~12天PAL活性小幅增加,12天活性52.08 U·g-1min-1,12~16 天大幅降低,16~20天急剧增加, 20天达到峰值(148.33 U·g-1min-1),显著高于其他处理天数(P< 0.05), 20~32天PAL活性再次出现多次波动,28天出现小高峰(活性为65.42 U·g-1min-1)。与对照相比,PVP处理PAL活性在整个处理周期内一直处于相对较低的水平,活性为4.58~31.67 U·g-1min-1, 20天时活性最高(31.67 U·g-1min-1),但仍显著低于对照20天的活性(P< 0.05)(图3B)。由此看出,与对照相比,PVP处理从第8天开始PAL活性明显受到抑制,观察发现PVP处理表现出明显抑制黑皮油松外植体褐化的时间点也是从第8天开始,褐变严重的对照PAL活性显著高于外植体褐变程度轻的PVP处理,因此推测PVP处理可能通过抑制PAL活性来阻止黑皮油松外植体发生褐变。
3) POD活性的动态变化 随着培养时间的延长,对照外植体第4天时POD活性显著增加,4~16天POD活性下降,16~20天急剧增加, 20天出现峰值,活性为1 419.12 U·g-1min-1,显著高于其他处理天数(P< 0.05),随后POD活性再次降低直至培养结束。与对照相比,PVP处理的外植体POD活性呈现不同的变化趋势,8天时出现第1个小高峰(488.33 U·g-1min-1),随后降低,16~28天POD活性再次提高(717.08~818.75 U·g-1min-1),显著高于其他处理天数(P< 0.05),28~32天时活性再次降低(图3C)。整体来看POD活性与褐变数据变化的规律不明显,可能由于取样持续时间历时较长,培养基成分和萌发生长阶段等多因素影响导致POD活性测定结果呈现波动式变化。
2.3 PVP处理过程中酚酸类物质含量的动态变化1) 绿原酸含量动态变化 与对照相比,PVP处理的外植体中绿原酸含量在整个培养周期始终处于较高的水平,从第4天开始直至培养结束均显著高于对照,且整体呈现“降—升—降—升”的交替式变化,PVP处理0~12天绿原酸含量逐渐降低,随后再次提高,16天出现小高峰(含量为1.875 μg·mL-1),但显著低于0 天的含量,16~20天再次降低后有小幅度的增加,随后保持稳定(图4A)。上述研究结果表明PVP处理从第8天开始保护外植体内绿原酸不被氧化,进而抑制黑皮油松外植体发生褐变。
2) 香豆酸含量动态变化 对照与PVP处理过程中香豆酸含量变化出现多次波动,但整体来看对照与PVP处理香豆酸含量较为接近,始终维持在较低的水平(0.312~0.825 μg·mL-1),据此推测PVP处理抑制黑皮油松外植体褐化的作用可能与香豆酸关系较小。具体来看,对照处理0天香豆酸含量最高(0.825 μg·mL-1),除与20天差异不显著外,均显著高于其他处理天数(P< 0.05),0~12 天香豆酸含量逐渐降低,12~20 天逐渐增加,随后再次降低。PVP处理香豆酸含量呈现“降(0~8天)—升(8~16天)—降(16~20天)—升(20~24天)—降(24~32天)”交替式变化(图4B)。
3) 阿魏酸含量动态变化 对照外植体中阿魏酸含量在0~16 天递减,16~32天小幅波动; 而PVP处理则出现多次波动,呈现“降(0~12 天)—升(12~16 天)—降(16~20 天)—升(20~24 天)—降(24~32 天)”交替式变化。0~12 天PVP处理与对照阿魏酸含量较为接近,变化趋势也类似; 但16~32 天PVP处理阿魏酸含量显著高于对照,24 天时PVP处理外植体阿魏酸含量最高(5.433 μg·mL-1),显著高于其他处理天数(P< 0.05)(图4C)。据此推测PVP处理抑制外植体褐变可能主要是通过保护绿原酸和阿魏酸不被氧化而起作用,其中对绿原酸在全程起到保护作用,对阿魏酸的保护作用仅在中后期(16~32 天)。
3 讨论
3.1 PVP处理黑皮油松休眠芽外植体的抗褐化效果 PVP作为一种无机吸附剂,通过吸附外植体释放出来的酚类物质,阻止酚类物质不被氧化进而达到抑制外植体褐变的目的,因此在植物组织培养中广泛用于抑制褐化,而且效果显著(Babbaretal., 1986; 饶慧云等, 2015; 高红兵等, 2017; 彭春雪等, 2019)。本研究结果表明,300~1 500 mg·L-1PVP处理可以有效抑制黑皮油松休眠芽外植体的褐化,并促进休眠芽启动萌发。叶睿华等(2018)的研究也发现PVP对杜鹃兰(Cremastraappendiculata)原球茎褐化具有抑制作用,并能提高原球茎增殖率。本研究筛选出PVP最佳处理浓度为1 500 mg·L-1,说明培养基中添加适宜浓度的PVP能有效吸附外植体释放的酚类物质,阻止酚类物质被氧化成醌类物质,从而达到抑制外植体褐化的效果,但当PVP处理浓度增至2 000 mg·L-1时,防褐化效果显著降低,说明高浓度的PVP会影响其抑制褐化作用的发挥。葡萄愈伤组织的继代研究中也发现2 000 mg·L-1PVP有助于抑制褐化,但增至3 000 mg·L-1时褐化率再次增加(饶慧云等, 2015),分析可能是由于PVP浓度过高导致其吸附酚类物质的同时也吸附培养基中的营养物质,进而影响了植物对有益物质的吸收和利用,最终影响抗褐化效果(Teixeriaetal., 1994)。
3.2 主要酚酸、酶活性与外植体褐化之间的关系 褐化是复杂的生化过程,由酶引起细胞内的氧化褐变是组培中外植体褐化的主要方式。PPO、POD和PAL被认为是在组培褐变过程中发挥作用较大的酶,其中PPO是引起酚类物质氧化的主要催化酶; PAL是酚类物质前体合成中的关键酶,可通过抑制PAL活性,抑制酚类的积累和合成,从而抑制褐变的发生; POD可在被切割外植体释放的H2O2作用下促进酚类物质氧化导致组织褐变(Pérezetal., 2012)。
本研究中综合PVP抑制黑皮油松外植体褐变时间效应与酶活作用时间点数据发现,CK处理PPO、PAL活性在8~12天或20~24 天急剧增加,这2个时间点也恰好是外植体褐变愈发严重的时期,而PVP处理在第8 天开始褐变程度明显轻于对照,与此同时PPO、PAL活性也显著低于CK处理,说明PVP抑制黑皮油松外植体褐化过程伴随着PPO、PAL活性降低,PVP可通过抑制PPO活性的增加阻止酚类物质的氧化而导致形成醌类物质的减少,或者通过影响PAL的活性进而阻止酚类物质的合成,最终有效抑制黑皮油松外植体褐变。阿月浑子(Pistaciavera)组培苗(冷平生等, 2012)和葡萄愈伤组织(饶慧云等, 2015)研究中也发现,PVP处理下PPO活性明显低于对照,PVP可通过抑制PAL活性增加有效抑制桃(Prunuspersica)果肉褐变(杨青珍等, 2016)。最近芍药(Paeonialactiflora)研究中也发现,抑制褐变过程伴随着PPO、PAL活性的降低(Caietal., 2020)。对于POD与褐化的关系说法不一,不同植物材料差异较大。马铃薯(Solanumtuberosum)块茎酶促褐变程度与PPO和POD活性显著正相关(王丽等, 2019); 而凤丹牡丹(Paeoniaostii)愈伤组织褐变与POD活性呈负相关(毛沛琪等, 2018)。据此推测在黑皮油松组培中PVP主要通过抑制PPO和PAL的活性来阻止酚酸被氧化或者抑制酚类物质的合成,进而阻止黑皮油松休眠芽外植体发生褐变。因此在今后的黑皮油松组培中可通过抑制PPO、PAL活性以达到抑制褐变的目的。
酚类物质作为植物组培中褐变的物质基础,其种类和含量对组织褐变起到至关重要的作用,酚类物质含量越高,褐变越严重。本研究PVP处理过程中外植体绿原酸含量显著高于对照,说明PVP处理对外植体内的绿原酸具有保护作用,马铃薯(Fengetal., 2020)和天女木兰(高红兵等, 2017)抗褐化研究中也得出类似的结论。此外,研究发现添加48.5 mg·L-1阿魏酸能显著抑制地涌金莲(Musellalasiocarpa)愈伤组织的褐化,说明阿魏酸在组织培养中具有抗褐化的作用(侯健华等, 2015)。与上述研究结果类似,在本研究中也证实了阿魏酸具有抑制黑皮油松外植体发生褐变的作用,本研究中PVP处理与对照0~12 天阿魏酸含量接近,且变化趋势类似,16~32 天阿魏酸含量显著高于对照,说明除绿原酸外,PVP处理也可以有效阻止阿魏酸被氧化,进而起到抑制外植体发生褐变的作用。本研究中香豆酸含量在PVP与对照处理全程均处于较低水平,说明香豆酸可能化学性质相对稳定不易被氧化,天女木兰的研究中也得到相似的结论(高红兵等, 2017)。综上得出PVP处理抑制黑皮油松外植体褐变可能主要是通过保护绿原酸和阿魏酸起作用,其中全程对绿原酸起保护作用,而对阿魏酸仅在中后期发挥作用。
除上述3种酚酸外,本研究同时对没食子酸、儿茶酚、对羟基苯甲酸、咖啡酸进行了分析,但试验结果中仅检测到了绿原酸、香豆酸和阿魏酸3种酚酸,其余4种酚酸含量过低未检测出,说明这4种酚酸对黑皮油松外植体的褐化现象影响不大。
4 结论
300~1 500 mg·L-1PVP处理可以有效抑制黑皮油松休眠芽外植体的褐化,并促进休眠芽萌发,1 500 mg·L-1PVP为最佳处理浓度。抗褐化剂PVP通过抑制PPO、PAL活性来阻止黑皮油松外植体发生褐变。PVP处理抑制外植体褐变与绿原酸和阿魏酸不被氧化有关,PVP处理可以对绿原酸全程起保护作用,而对阿魏酸仅在中后期起作用。
目前对于抗褐化剂处理抑制外植体褐化的时间效应的研究相对较少,本研究探讨了PVP抑制黑皮油松外植体褐化的时间效应及相关酶活性、酚酸类物质与外植体褐变的关系,结果可为黑皮油松组织培养过程中褐变这一瓶颈问题的解决提供理论依据和技术支撑。
猜你喜欢
安徽农业大学学报(2022年2期)2022-11-09
现代食品科技(2022年7期)2022-07-29
农业工程学报(2022年6期)2022-06-27
中草药(2022年1期)2022-01-13
烟台大学学报(自然科学与工程版)(2021年1期)2021-03-19
中华养生保健(2020年4期)2020-11-16
山东农业科学(2016年11期)2016-12-17
江苏农业科学(2016年9期)2016-11-28
医学研究杂志(2015年12期)2015-06-10
分析化学(2014年7期)2014-12-13
