
种内捕食对杂食食物链中群落动态的影响
2022-01-09 10:20张荣周帅朱文君周晓梅李小珍
六盘水师范学院学报 2021年6期
张荣周帅朱文君周晓梅李小珍
(1皖西学院金融与数学学院,安徽 六安237012;2安徽国防科技职业学院基础教学部,安徽 六安237011;3合肥经济学院基础教学部,安徽 合肥230016)
尽管种内捕食现象在食物网中普遍存在,且广泛存在于集团内捕食系统中(IGP),但它对物种间相互作用、群落动态、群落结构的影响仍不清楚。除此之外,现有的关于种内捕食的研究多数集中在顶级捕食者的种内捕食上,且忽略了生物体型、年龄等因素的影响。然而,一组结构模型表明:种内相食可以完全改变三种群IGP系统的动力学和结构,这取决于种内相食发生的营养位置。而且,从出生到成熟阶段,生物的体型一般会随着年龄的增长不断变大,捕食幼体和捕食成体的难度和能量回馈都是不一样的。最简单的杂食形式之一是集团内捕食(IGP),即杂食性捕食者与其食饵竞争同一种资源。基于Holt和Polis[1]提出的开创性理论以及该模型的后续扩展[2-5],人们普遍认为IG捕食者和IG食饵的共存参数空间非常狭窄,且要求食饵在资源开发上更有效率。这些预测证实了先前的观点,即杂食性食物链与简单线性食物链相比具有不稳定性[6]。尽管在微生物系统中有一些实验支持这些预测[2,7-8],但这通常与经验数据和最近的分析相冲突,这些数据和分析表明,集团内捕食是一种常见且稳定的物种相互作用形式[9-12]。此外,实地试验并没有一致地表明,IG捕食者在资源开发方面效率较低[13]。这就提出了一个问题:在当前的模型中,是否遗漏了现实的某些关键部分。
种内捕食指的是吃掉和杀死同类个体的过程。从社会人类学和生态学的角度来看,种内捕食是一种在世界范围内发生的令人憎恶的现象。在由甲虫、蝗虫、昆虫、蜘蛛和昆虫组成的一系列动物中均会发生种内捕食现象[14-18]。通常情况下,捕食和被捕食对象属于不同的成熟阶段,如成体期和幼体期。然而在有些情况下,成体捕食者会捕食成体同类。此类研究比较少,需要补充。在捕食者—食饵模型的动力学中,种内捕食通常是一个生存过程,事实上也比较常见[19],相关研究可以参考文献[20-21]。在生态学和数学生物学领域中,种内捕食现象是研究的热点,由于一些反直觉的影响,种内捕食现象能够影响种群动态。与资源相比,种内捕食现象出现在许多种群规模非常大的物种中[22]。最初的种内捕食仅考虑了捕食者[23-24],然而在自然界中,在食饵中也经常发生种内捕食[25-27]。因此,有必要研究不同营养级内部发生的种内捕食的影响。Rudolf研究了在食饵中发生种内捕食的影响[27],然而没有考虑生物体型在种内捕食中差异,因此需要进一步补充。
本文基于Rudolf[26]的集团内捕食系统模型,建立两个新的模型,分别研究在捕食者和食饵两个营养级内发生的成体对成体以及成体对幼体种内捕食的影响。
1 模型
模型1:在捕食者中发生种内捕食。模型具体假设为:
(1)资源的生长遵循Logistic生长规律;资源的消耗主要体现食饵、成体捕食者、幼体捕食者对资源的开发利用;
(2)成体捕食者、幼体捕食者、食饵均会开发利用资源,其中食饵利用资源增长自身密度,成体捕食者开发的资源用来维持生存和繁殖后代,而幼体捕食者由于年龄和体型尚小,无法生殖,因此开发的资源主要用来自身的生存和生长,体现在转化为成体捕食者方面;
(3)成体捕食者可以捕食食饵,幼体捕食者不行;
(4)捕食者内会发生成体对成体的种内捕食以及成体对幼体的种内捕食。
基于以上假设,建立如下模型(模型1、2中参数定义见表1):

表1 模型1、2中参数定义
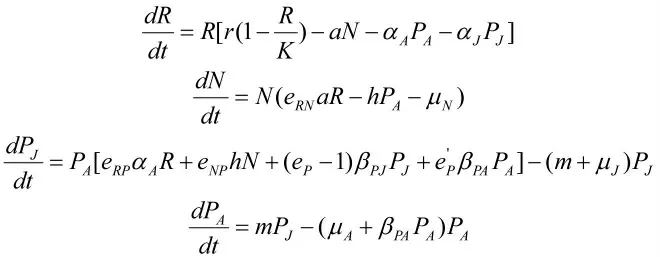
模型2:类似的方法可以建立在食饵中发生种内捕食的模型,其微分方程组表示如下:
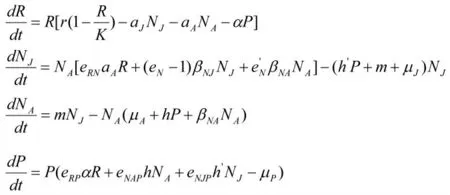
2 模拟结果
通过对模型1和模型2利用MATLAB进行数值模拟,分析捕食者和食饵种群中成体对成体、成体对幼体种内捕食作用对群落动态的影响。
2.1 捕食者种内捕食对群落动态的影响
图1A中没有考虑种内捕食的影响(βPJ=βPA=0),在环境容纳量较低时,捕食者由于ALLEE效应、资源利用率较低等因素导致在和食饵的竞争中处在不利地位,一直处于灭绝状态;而当环境容纳量超过10时,捕食者的密度迅速上升,食饵的密度则迅速下降直至灭绝,最终捕食者单独存在,这主要是由于环境容纳量增加使得捕食者的密度超过了建群阈值,捕食者能够建立种群,此外,捕食者可以通过捕食食饵、利用资源实现自身密度的增加。图1B中则考虑了捕食者中成体对幼体种内捕食作用的影响(βPJ=0.02,βPA=0),模拟显示:资源、食饵、幼体捕食者和成体捕食者的总体变化趋势和图1A一致,然而食饵灭绝的容纳量要求更大了,这表明捕食者中成体对幼体种内捕食有利于食饵的生存,主要是由于成体捕食者能够通过捕食幼体捕食者获得能量,食饵灭绝时资源、幼体捕食者和成体捕食者密度和图1A几乎一致。图1C中考虑了捕食者中成体对成体的种内捕食作用的影响(βPJ=0,βPA=0.01),模拟显示:随着环境容纳量的增大,捕食者和食饵实现了共存,且食饵的密度远高于捕食者的密度,对比图1B、C中食饵的变化动态可以得出:捕食者中成体对成体的种内捕食作用的影响强于捕食者中成体对幼体的种内捕食作用。而对比图1A、B、C中资源的变化动态可知:食饵对资源的利用率要高于捕食者。图中模拟的时间T=1 000,大约经历了1 000多代,模拟时间足以保证物种达到稳定状态。
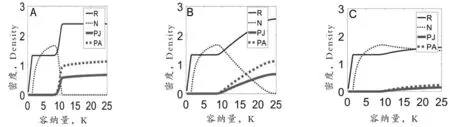
图1 环境容纳量和捕食者种内捕食对资源、食饵、幼体捕食者和成体捕食者密度的影响

图2分别模拟了捕食者成体幼体以及成体对成体种内捕食率对资源、食饵、幼体捕食者和成体捕食者密度的影响,随着捕食强度的增加,资源、食饵、幼体捕食者和成体捕食者的变化趋势总体一致,但是当βPA=0.03时,捕食者就灭绝了,而βPJ=0.06时,捕食者仍然能够续存。这表明:βPA对群落的影响力度明显强于βPJ,这与图1得出的结论一致。

图2 捕食者种内捕食率对资源、食饵、幼体捕食者和成体捕食者密度的影响
2.2 食饵种内捕食对群落动态的影响
图3A、B、C分别模拟了无种内捕食作用,食饵中成体对幼体种内捕食作用以及食饵中成体对成体种内捕食作用对群落动态的影响。图3A中可以得出:随着环境容纳量的增加,捕食者的密度逐渐由0慢慢开始线性增加,开始增加的容纳量大约为1.75;图3B、C中资源、食饵、幼体捕食者和成体捕食者密度的整体变化趋势和图3A一致,但是捕食者密度开始增加要求的容纳量分别增加为2.25(图3B)、2.35(图3C)。这表明食饵中成体对成体种内捕食作用略强于食饵中成体对幼体种内捕食作用。而在调节作用最强的图3C中,食饵的密度最低,主要是由于食饵通过种内捕食可以获取一定的能量,减弱了被捕食的风险,弱化了种间接触作用。
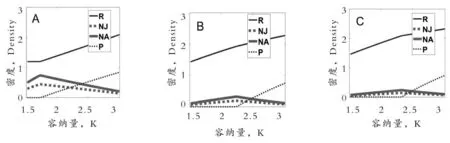
图3 环境容纳量和食饵种内捕食率对资源、食饵、幼体捕食者和成体捕食者密度的影响
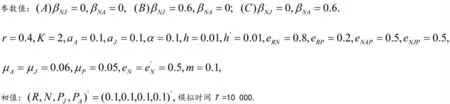
对于模型1和模型2,可以令模型的右边等于零,得出平衡点表达式,其共存稳定性可以通过局部稳定分析法进行数值模拟得出。
3 讨论
本文通过建立两个常微分方程组模型,分别研究了捕食者和食饵种内捕食对杂食食物链群落结构的影响,结果显示:成体对成体的种内捕食和成体对幼体的种内捕食产生的影响是不同的,成体对成体的影响力大于成体对幼体的影响力,这主要是由于成体的体型明显高于幼体,蕴含的能量物质亦较高,捕食一个成体产生的益处明显高于幼体,尤其在捕食者种内捕食模型中。而在食饵种内捕食模型中,也有同样的结论,但是影响力差别没有那么明显,主要是由于食饵所处营养级较低,对变化不敏感。此外,两个模型中的种内捕食均有利于食饵的生存,捕食者种内捕食相当于给捕食者提供了另外一种生存的选择,进而减弱了对食饵的捕食强度;而食饵种内捕食使得自身密度降低,减少了对资源的开发,多出来的资源能够养活捕食者,减弱了捕食者对食饵的种间捕食作用。本文的研究进一步补充了Rudolf[26]的研究结论,为构建更加合理群落,丰富物种多样性提供了方法和指导。
尽管如此,文章考虑的是比较简单的食物链模型,而在自然界中,多物种主要是以复杂食物网联系在一起的,因此,可以进一步研究食物网中种内捕食的影响。此外,食饵利用避难所可以帮助自身躲避捕食者[28],可以进一步将食饵策略融入模型中,而且可以延伸到植物学[29]和遗传学[30]中。
猜你喜欢
当代水产(2022年8期)2022-09-20
中国海洋大学学报(自然科学版)(2022年9期)2022-09-05
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
百科探秘·海底世界(2020年11期)2020-12-31
水产养殖(2020年9期)2020-09-24
动漫星空(兴趣英语)(2018年4期)2018-10-30
河北渔业(2018年4期)2018-05-08
中外文摘(2016年13期)2016-08-29
河北渔业(2015年1期)2015-01-19
