
高脂饮食小鼠脂肪组织胞外囊泡损伤海马神经元的研究
2022-01-14 09:38吕子阳聂盛丹周高雅
中国药理学通报 2022年1期
周 迪, 吕子阳, 聂盛丹, 周高雅, 王 珊
(1. 中南大学化学化工学院制药工程系,湖南 长沙 410083, 2. 湖南省人民医院药物临床试验机构办, 湖南 长沙 410081, 3.湖南省脑科医院神经内科,湖南 长沙 410007)
随着世界经济的快速发展,以高脂饮食为主的饮食模式越来越普遍。长期高脂饮食使机体脂肪异常堆积,引发代谢紊乱,损伤中枢神经系统的功能[1]。有研究显示高脂饮食可以引发认知障碍[2],但其具体机制尚未明确。长期高脂饮食可通过促进脂肪组织释放各种脂肪因子,从而影响其他组织器官的功能,进而介导疾病的发生发展[3]。胞外囊泡(extracellular vesicles,EVs)是由细胞分泌的一类纳米级膜囊泡,可在细胞间传递蛋白质、脂质、核酸等遗传信息,帮助维持生理稳态或介导疾病发展[4-5]。研究发现,高脂饮食小鼠脂肪组织分泌的胞外囊泡可以增加肝细胞、巨噬细胞中炎性因子的表达,加快相关并发症的发展[6-7]。同时,越来越多的研究表明,许多外周细胞,如红细胞、间充质干细胞等,分泌的胞外囊泡可以透过血脑屏障调节中枢神经系统疾病[8-9]。然而,高脂饮食小鼠脂肪组织胞外囊泡能否透过血脑屏障直接损伤海马神经元,尚未见相关报道。因此,本研究通过建立高脂动物模型,提取脂肪组织胞外囊泡,分析海马神经元对其摄取情况,并检测其对海马神经元树突发育及细胞活性的影响,旨在探讨高脂饮食引发认知障碍的机制。
1 材料与方法
1.1 材料
1.1.1实验动物 C57BL/6J雄性小鼠(22只),体质量为(21±2)g,SPF级(湖南省斯莱克景达动物实验公司提供),许可证号:SCXK(湘)2019-0008,饲养于中南大学湘雅医学院实验动物中心。饲养于21 ℃~25 ℃,湿度46%~54%,光照12 h的环境中。其中20只用于高脂饮食分组和造模,并提取脂肪组织胞外囊泡,余下2只用于尾静脉注射观察海马组织对脂肪组织胞外囊泡的摄取。孕18 d C57BL/6J雌鼠(1只),SPF级(湖南省斯莱克景达动物实验公司提供),许可证号:SCXK(湘)2019-0009,用于提取、培养原代海马神经元。实验过程中无动物意外死亡。
1.1.2主要试剂 小鼠标准颗粒状饲料脂肪占比0.04,为SJA常规啮齿类动物饮食,由湖南斯莱克实验动物有限责任公司提供;高脂饲料(货号:D12492I)脂肪占比0.60,由美国Research Diets公司提供。
Neurobasal培基(货号:21103049)、DMEM/F12培基(货号:11320033)、B27营养因子(货号:A3582801)、L-谷氨酰胺(货号:25030081)、胎牛血清(货号:A31608-02)、双抗(货号:15240062)购自美国Gibco公司,左旋多聚赖氨酸(货号:P1399)、PKH 67染料(货号:MKCK5701)购自美国Sigma公司,胶原酶I(货号:1904GR001)购自德国BioFroxx公司;MAP-2抗体(货号:ab5392)、β-tubulin III抗体(货号:ab18207)购自英国Abcam公司,CCK-8试剂盒(货号:KGA317)、BCA蛋白定量试剂盒(货号:KGPBCA)购自江苏凯基生物公司。
1.1.3仪器 Morris水迷宫装置(北京吉安得尔科技有限公司);电子血糖仪(三诺生物传感股份有限公司);酶标仪(赛默飞世尔科技公司);超高速离心机(美国贝克曼库尔特有限公司)。
1.2 实验方法
1.2.1分组及造模 将健康8周龄小鼠20只随机分为两组,即正常饮食组(normal diet,ND),高脂组(high-fat diet,HFD),每组10只。ND组给予足够的标准动物饲料;HFD组给予足够的高脂饲料,两组均自由饮水,共喂养28周,每周检测体重1次[10]。为避免采集血糖的刺激导致动物的应激反应影响水迷宫的检测结果,我们于进入第27周的d 1分别测量血糖值(fasting blood glucose, FBG)、胰岛素水平(insulin, INS),计算胰岛素抵抗指数(HOMA-IR),于进入第28周的d 1进行行为学检测。
1.2.2空腹血糖及胰岛素检测 空腹血糖及胰岛素水平检测于第27周进行,检测前将小鼠禁食6 h,对小鼠剪尾用毛细管采血约100 μL,分别用电子血糖仪检测血糖和胰岛素检测试剂盒检测胰岛素水平。
1.2.3糖耐量试验及胰岛素耐量试验 糖耐量试验于空腹血糖及胰岛素检测后隔天进行,试验前,禁食16 h,于d 2早8点给小鼠称重,以2 g·kg-1的剂量给小鼠腹腔注射葡萄糖溶液,分别于注射0 min、15 min、30 min、60 min、90 min、120 min后,用采血针从尾静脉取血,擦去第一滴血,并用电子血糖仪检测第二滴血的血糖值。
胰岛素耐量试验在糖耐量试验后隔天进行。试验前,将小鼠禁食6 h,以1 U·kg-1的剂量给小鼠腹腔注射胰岛素溶液,分别于注射0 min、15 min、30 min、60 min、90 min、120 min后,通过采血针尾静脉采血测量小鼠血糖值。
1.2.4Morris水迷宫实验 Morris水迷宫实验于第28周d 1开始进行,用于研究空间识别记忆能力。水迷宫直径100 cm,平台直径10 cm。水迷宫分为四个象限,平台置于一个象限中间。d 1平台露出水面1 cm,d 2~5将平台没入水下1 cm,d 6撤掉平台。前5 d进行获得性训练,将小鼠放入迷宫,记录小鼠从入水到站上平台所用时间,作为逃避潜伏期。如果小鼠在60 s内站上平台,则允许它们在平台上停留5 s,如果未找到平台,则用木棍引导小鼠站上平台并停留15 s。所有小鼠每日进行4次训练。最后1 d进行探查训练,拆除平台,记录小鼠入水中后在目标象限(原先平台放置的象限)的停留时间。
1.2.5脂肪组织取材、称重及HE染色 造模完成后,两组小鼠用异氟烷麻醉,剪开腹部皮肤,取出附睾脂肪(epididymal white adipose tissue, eWAT),称量脂肪质量后,一半固定做HE染色,一半用于脂肪组织胞外囊泡提取。
脂肪组织取材后,用多聚甲醛固定2 d,制成石蜡切片后进行苏木精-伊红染色,于显微镜下观察。
1.2.6脂肪组织胞外囊泡提取及鉴定 脂肪组织取材后立刻用预冷PBS漂洗,加入3 mL I型Collagenase消化液,于恒温震荡仪上37 ℃消化30 min,用完全培养基终止消化,400×g离心1 min,留上清液重悬于缓冲液中,重复3次,用含去胞外囊泡血清培养基重悬,转移至90 mm培养皿培养24 h,收集培基。收集的培基2 500×g离心20 min,10 000×g离心30 min,并用0.22 μm滤器对其过滤,100 000×g离心2 h得到胞外囊泡及蛋白沉淀,再PBS缓冲液清洗(100 000×g, 2 h)得到脂肪组织胞外囊泡。
对提取的胞外囊泡进行鉴定,取部分样品滴加在铜网上,用乙酸双氧铀进行负染, 使用透射电子显微镜(TEM)对其进行结构分析;另取部分样品用纳米颗粒跟踪分析仪(NTA)对其进行粒径分析。
1.2.7原代海马神经元细胞培养 在24孔板中预先放置细胞爬片,用0.1 mg·L-1的Poly-lysine溶液进行包被,使用前用PBS清洗3遍。取孕18 d左右的C57BL/6J鼠1只,异氟烷麻醉,在医用酒精中浸泡30 s,取出胎鼠,断头后,用显微镊划开头骨,取出整个大脑于预冷的HBSS中,分离海马组织。将取出的海马组织剪碎,加入胰蛋白酶,于37 ℃消化15 min,加入FBS终止消化,轻轻吹打,放入离心机中1 000 r·min-1离心5 min,弃去上清,制成单细胞悬液,以2×105个·L-1的浓度种板,4 h后换液,用PBS洗3遍,换成neurobasal培基,每2 d半量换液。
1.2.8脂肪组织胞外囊泡的染色 向1 mL的Diluent C溶液中加入4 μL PKH 67试剂,振荡混匀后加入胞外囊泡样品,25 ℃ 孵育10 min,加入等量20 g·L-1BSA溶液孵育1 min终止染色后,重复1.2.6中提取胞外囊泡的方法,收集染色后的胞外囊泡,染色全过程避光操作。
1.2.9海马神经元对脂肪组织胞外囊泡的摄取 取8周左右C57BL/6J雄鼠2只,分别尾静脉注射蛋白总量30 μg的PKH 67染色的脂肪组织胞外囊泡及PBS,30 h后将小鼠麻醉处死,迅速取出脑组织保存于-80 ℃,取海马区冠状切片,染核,采用倒置荧光显微镜观察。
向体外培养8 d的原代海马神经元中加入20 mg·L-1PKH 67染色的脂肪组织胞外囊泡,4 h后用多聚甲醛固定,并用anti-MAP-2抗体(1 ∶500)进行免疫荧光染色,Hochest染核。
1.2.10高脂饮食小鼠脂肪组织胞外囊泡对神经元的影响 用20 μg·mL-1高脂饮食小鼠脂肪组织胞外囊泡(HFD-EVs组)、正常饮食小鼠脂肪组织胞外囊泡(ND-EVs组)及PBS(Control组)分别处理神经元,用MAP-2做免疫荧光观察树突数量,分析胞外囊泡对神经元形态的影响[11]。
神经元接种于96孔板,培养贴壁后,进行细胞活性检测。每组设5个复孔,分别加入20 mg·L-1高脂饮食、正常饮食小鼠脂肪组织胞外囊泡及PBS孵育培养1 d后,每孔加CCK-8溶液10 μL,37 ℃孵育2 h,酶标仪450 nm测定吸光度,实验重复3次。

2 结果
2.1 高脂饮食对小鼠体质量及糖代谢指标的影响分组造模后,HFD组小鼠的体质量、附睾脂肪质量、血糖、胰岛素水平及胰岛素抵抗指数明显高于ND组(P<0.05),见Tab 1。
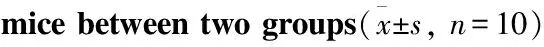
Tab 1 Comparison of obesity indexes of
葡萄糖耐量实验中2组小鼠血糖值均随时间呈先上升后下降的趋势,于15 min时达到最高值。注射葡萄糖溶液后,HFD组血糖值均高于ND组。在30 min和120 min时,2组血糖值差异存在显著性(P<0.05),见Tab 2。胰岛素耐量试验中各组小鼠血糖值均随时间呈先下降后上升的趋势,HFD组血糖值均高于ND组。在15 min、30 min和60 min时,两组血糖值差异存在显著性(P<0.05),见Tab 3。
2.2 高脂饮食对小鼠学习记忆能力的影响获得性训练d 1、2、3,2组小鼠的逃避潜伏期无明显差异(P>0.05),表明小鼠体质量对逃避潜伏期影响较小。获得性训练d 4、5,与ND组相比,HFD组逃避潜伏期明显延长(P<0.05),且游泳轨迹杂乱无章,见Fig 1A,D。探查训练结果表明,HFD组小鼠进入目标象限次数及在目标象限中游泳时间明显低于ND组(P<0.05),见Fig 1B,C。提示高脂饮食可损伤小鼠的学习记忆能力。

Fig 1 Results of Morris water maze

Tab 2 Changes of blood glucose at each time point in glucose tolerance

Tab 3 Changes of blood glucose at each time point in insulin tolerance
2.3 高脂饮食对小鼠脂肪组织形态学及其分泌的胞外囊泡的影响HE染色结果显示,ND组小鼠脂肪细胞大小均一;HFD组小鼠脂肪细胞大小不一,且平均面积较ND组明显增大,见Fig 2。

Fig 2 Influence of high-fat diet on adipose
透射电镜结果显示脂肪组织提取的胞外囊泡呈杯状囊泡结构,形态完整,NTA结果显示粒径主要分布在50~150 nm之间,见Fig 3A,B。BCA蛋白定量结果显示,HFD组小鼠脂肪组织分泌胞外囊泡的量明显多于ND组(P<0.05),见Fig 3C。
2.4 海马神经元对脂肪组织胞外囊泡的摄取情况对培养的原代海马神经元进行纯度鉴定,免疫荧光结果显示神经元细胞形态正常且纯度达到98%以上,见Fig 4A,可用于后续实验。体外孵育PKH 67标记的脂肪组织胞外囊泡,免疫荧光结果显示,原代海马神经元能够摄取脂肪组织胞外囊泡,见Fig 4B。小鼠尾静脉注射PKH 67标记的脂肪组织胞外囊泡,在海马组织中可检测到明显的荧光信号,见Fig 4C,提示脂肪组织胞外囊泡可以通过血脑屏障被海马神经元摄取。
2.5 高脂饮食小鼠脂肪组织胞外囊泡对海马神经元的影响用相同浓度的高脂饮食、正常饮食小鼠脂肪组织胞外囊泡及PBS分别处理原代海马神经元,免疫荧光结果显示,HFD-EVs组海马神经元较Control组及ND-EVs组,树突长度降低、初级树突数和次级树突数减少,见Fig 5。
CCK-8结果显示,高脂组小鼠脂肪组织胞外囊泡处理神经元使神经元细胞活性下降(P<0.05),见Fig 6。

Fig 3 Influence of high-fat diet on EVs derived from adipose

Fig 4 EVs purified from adipose uptaken by hippocampal neurons

Fig 5 Effects of EVs purified from adipose on morphology of hippocampal neurons

Fig 6 Effect of EVs on cell viability of
3 讨论
长期的高脂饮食可导致肥胖,并诱导心血管疾病及脂肪性肝病、糖尿病等代谢相关性疾病的发生[12-14]。近年来,高脂饮食与神经系统疾病的关系也倍受瞩目[2]。研究发现在动物及临床试验中,高脂饮食会导致海马功能受损,引发认知障碍[2, 15]。但是高脂饮食介导认知障碍的具体机制仍有待深入研究。
脂肪组织不仅可以平衡能量的储存和消耗,还可以作为一种内分泌器官,分泌各种脂肪因子影响机体代谢[16]。有研究表明,脂肪组织的异常堆积是诱发胰岛素抵抗、2型糖尿病及代谢综合征的重要驱动因素[17]。本研究中,HFD组小鼠的体质量、附睾脂肪质量、血糖、胰岛素水平明显高于ND组,出现胰岛素抵抗状态。与ND组相比,HFD组小鼠在Morris水迷宫中逃避潜伏期时间明显延长,目标象限内游泳时间明显减少,表明高脂饮食能够介导认知功能障碍。
近年来,一些研究者开始关注脂肪组织来源的胞外囊泡在多种疾病中的作用,他们证实高脂饮食可通过影响脂肪组织分泌的胞外囊泡引起肝细胞、巨噬细胞等外周细胞的损伤[6-7]。此外,因胞外囊泡的纳米级膜结构,使其具有透过血脑屏障的能力[18];外周细胞产生的胞外囊泡可以携带蛋白或药物顺利透过血脑屏障调节中枢系统疾病[8-9]。那么,脂肪组织分泌的胞外囊泡是否可以透过血脑屏障直接造成中枢神经系统的损伤呢?本研究结果表明,脂肪组织分泌的胞外囊泡可被原代培养的海马神经元摄取,通过尾静脉给小鼠注射标记过的脂肪组织胞外囊泡,可在海马组织中检测到明显的荧光信号。提示,脂肪组织胞外囊泡可被脑组织海马神经元摄取,病理状态下脂肪组织胞外囊泡的改变可能介导海马依赖性认知功能障碍的发生。
长期高脂饮食诱导脂肪组织异常堆积,很可能改变了脂肪组织分泌的胞外囊泡的分泌量及内容物[19]。在本研究中,相比ND组,HFD组小鼠脂肪细胞体积显著增大,其分泌的胞外囊泡明显增多;HFD组小鼠脂肪组织胞外囊泡可明显降低神经元细胞活性,减少海马神经元树突分支总数和总长度。提示,高脂饮食可能通过影响脂肪组织分泌的胞外囊泡损伤海马神经元,介导小鼠认知障碍的发生。但给正常小鼠多次注射高脂动物脂肪组织胞外囊泡是否足以导致小鼠认知功能障碍,有待后续进一步研究。
综上所述,长期高脂饮食会使小鼠脂肪堆积,出现糖耐量异常、胰岛素抵抗,学习记忆能力下降,其机制可能与脂肪组织分泌的胞外囊泡异常有关。本研究提示了高脂饮食介导认知障碍的可能机制,为高脂饮食介导认知障碍的防治提供新思路。
猜你喜欢
昆明医科大学学报(2022年1期)2022-02-28
疯狂英语·新策略(2021年7期)2021-08-26
中老年保健(2021年9期)2021-08-24
昆明医科大学学报(2021年8期)2021-08-13
服饰导报·鞋世界(2021年4期)2021-05-17
家庭科学·新健康(2019年7期)2019-08-21
中国新闻周刊(2016年33期)2016-10-27
医学研究杂志(2015年12期)2015-06-10
食品工业科技(2014年5期)2014-03-11
网球俱乐部(2009年5期)2009-06-15
