
松脂醇二葡萄糖苷在人和大鼠肝微粒体中的代谢比较
2022-01-27 10:43凌海燕李春沁杨安东
中成药 2022年1期
凌海燕, 李春沁, 杨安东, 朱 宁,2*
(1.四川省中医药科学院,四川 成都 610041;2.成都中医药大学药学院,四川 成都 611137)
杜仲为杜仲科植物杜仲EucommiaulmoidesOliv.的干燥树皮,功效补肝肾、强筋骨、安胎[1],含有木脂素、环烯醚萜、苯丙素、多糖等多种活性成分,具有降血压、降血脂、降血糖、抗肿瘤、抗菌、抗病毒、保肝、护肾、抗骨质疏松等药理作用[2-3]。其中,木脂素有着降血压、抗骨质疏松、保护内皮细胞修复受损皮肤作用[4-8],并且松脂醇二葡萄糖苷为杜仲主要特征性、功能性成分[9],具有对血压的双向调节活性。Xie等[10]将杜仲皮松脂醇二葡萄糖苷与人粪便悬浮液厌氧培养,得到11种代谢产物,推测其复杂的肠道代谢或肝脏首过代谢产物可能是其药理作用来源。
微粒体体外代谢试验快速简便,可直接观察酶与底物之间的相互选择性作用,减少了诸多体内因素的干扰,能作为体内外整体研究可靠的理论来源[11],其中肝微粒体是最有效的手段。目前,国内外尚未见松脂醇二葡萄糖苷在肝微粒体中代谢的报道,故本实验采用高效液相色谱串联四极杆质谱(HPLC/QQQ-MS)法比较该成分在人和大鼠肝微粒体中的代谢,以期为该成分药理活性及作用机制的进一步研究奠定基础。
1 材料
1.1 仪器 Agilent 1200高效液相色谱仪、Agilent 6410B三重四级杆质谱联用仪(美国Agilent公司);XS205电子天平(瑞士Mettler-Toledo公司);MIX-25涡旋振荡器(杭州米欧仪器有限公司);Heraeus Multifuge X1R冷冻离心机(美国Thermo Fisher Scientific公司);CASCADA Ⅱ超纯水机(美国Pall life Science公司);DZKW-4电子恒温水浴锅(北京中兴伟业仪器有限公司);KH7200E超声波清洗仪(昆山禾创超声仪器有限公司)。
1.2 试剂与药物 松脂醇二葡萄糖苷(成都普思生物科技股份有限公司,批号PS000870);氯霉素(四川省维克奇生物科技有限公司,批号wkq20051911)。PBS缓冲液(北京汇智泰康生物科技有限公司,批号20k001);NADPH再生系统A液、B液[烟酰胺腺嘌呤二核苷酸磷酸四钠盐还原型,北京汇智泰康生物科技有限公司,批号分别为NRS(A)-20L001、NRS(B)-20L001]。人肝微粒体(美国BioIVT Elevating Science公司,Lot#ZZQ);雄性大鼠肝微粒体(瑞德肝脏疾病研究有限公司,批号JYYJ)。甲醇、乙腈、甲酸、甲酸铵均为色谱纯;水为超纯水。
2 方法与结果
2.1 LC-MS/MS分析
2.1.1 色谱条件 Agilent Extend-C18色谱柱(3.5 μm,2.1 mm×100 mm);流动相水(含0.5%甲酸、5 mmol/L甲酸铵)(A)-乙腈(B),梯度洗脱(0~4 min,10%B;4~6 min,60%~90%B;6~12 min,90%B;12~15 min,90%~10%B);体积流量0.3 mL/min;柱温25 ℃;进样量10 μL;内标氯霉素(16.24 μg/mL)。
2.1.2 质谱条件 电喷雾离子源;负离子模式;一级扫描、多反应监测方式;毛细管温度400 ℃;干燥气体积流量11 L/min;毛细管电压4 000 V;雾化器压力30 psi(1 psi=0.133 kPa);松脂醇二葡萄糖苷、内标检测离子对分别为m/z681.3~357.1、321.0~152.0,碰撞能量分别为25、20 V。
2.2 肝微粒体体外代谢体系建立和分组 孵育体系总体积为500 μL,包括PBS缓冲液(0.1 mol/L,pH 7.4)、各种属肝微粒体(含20 mg/mL蛋白)、0.452 0 mg/mL(0.67 μmol/L)松脂醇二葡萄糖苷、辅酶NAPDH再生系统A液和B液(按5∶1比例临用混合)。在试管(12 mm×75 mm)中将50 μL PBS缓冲液与360 μL超纯水混合,加入50 μL冰浴上预先融化的混合肝微粒体(20 mg/L)、10 μL松脂醇二葡萄糖苷底物溶液,涡旋混匀,在37 ℃下预孵育5 min,加入30 μL NADPH再生系统启动反应,继续在37 ℃下孵育,于0、5、15、30、45、60、90、120 min立即加入1 mL预冷、含内标的甲醇终止反应,涡旋2 min,4 ℃、12 000 r/min离心10 min,取上清液,0.22 μm微孔滤膜过滤,进样检测。
分别将人和大鼠肝微粒分为3组,依次为实验组、阴性组、空白组。其中,实验组为含松脂醇二葡萄糖苷的标准孵育体系,阴性组孵育体系中不含起始因子NAPDH,而空白组孵育体系中不含松脂醇二葡萄糖,每组3份,平行3次。
2.3 对照品溶液制备 精密称取松脂醇二葡萄糖苷对照品适量,置于量瓶中, 流动相溶解并定容至刻度,精密量取1 mL至25 mL量瓶中,流动相定容至刻度,即得。
本文中,提出了一种能够有效降低细胞光毒性、动态追踪并且精确量化三维细胞牵引力的3D-TFM方法。该方法将生物相容性的聚丙烯酰胺水凝胶作为细胞黏附和迁移的基底,然后在凝胶基底的表面修饰单层荧光颗粒作为人造散斑追踪细胞的运动,采用激光扫描共聚焦显微镜(Laser Scanning Confocal Microscope, LSCM)延时采集荧光颗粒的三维位置信息,并采用基于二阶形函数的数字体积相关技术计算位移场,根据基底材料的弹性模量和泊松比以及弹性力学基本理论[33],由位移场得到应力场。
2.4 线性关系考察 取不同质量浓度松脂醇二葡萄糖苷溶液,加到热失活的大鼠肝微粒悬浮液中,使反应液中底物最终质量浓度为0.37、0.75、1.51、3.01、6.02、15.05、30.1 μg/mL,按“2.2”项下方法操作,在“2.1”项条件下进样测定。以松脂醇二葡萄糖苷质量浓度为横坐标(X),松脂醇二葡萄糖苷、内标峰面积比值为纵坐标(Y)进行回归,得方程为Y=0.032 3X-0.017 2(R2=0.999 8),检测下限为6.8 ng/mL。
2.5 回收率、精密度试验 按“2.4”项下方法制备1.35、2.70、5.40 μg/mL底物溶液,在“2.1”项条件下各进样测定6次,测得相对回收率分别为90.5%、92.2%、95.6%,日内精密度RSD分别为2.6%、3.5%、3.2%,日间(3 d)精密度RSD分别为5.6%、4.5%、4.2%。
2.6 肝微粒体质量浓度对代谢速率的影响 设置肝微粒体质量浓度为0.2、0.5、1、2、5、10、20 mg/mL,松脂醇二葡萄糖苷质量浓度为3.01 μg/mL,温孵时间为120 min,结果见图1。由此可知,随着肝微粒体质量浓度增加,底物代谢速率升高,最终确定为20 mg/mL。
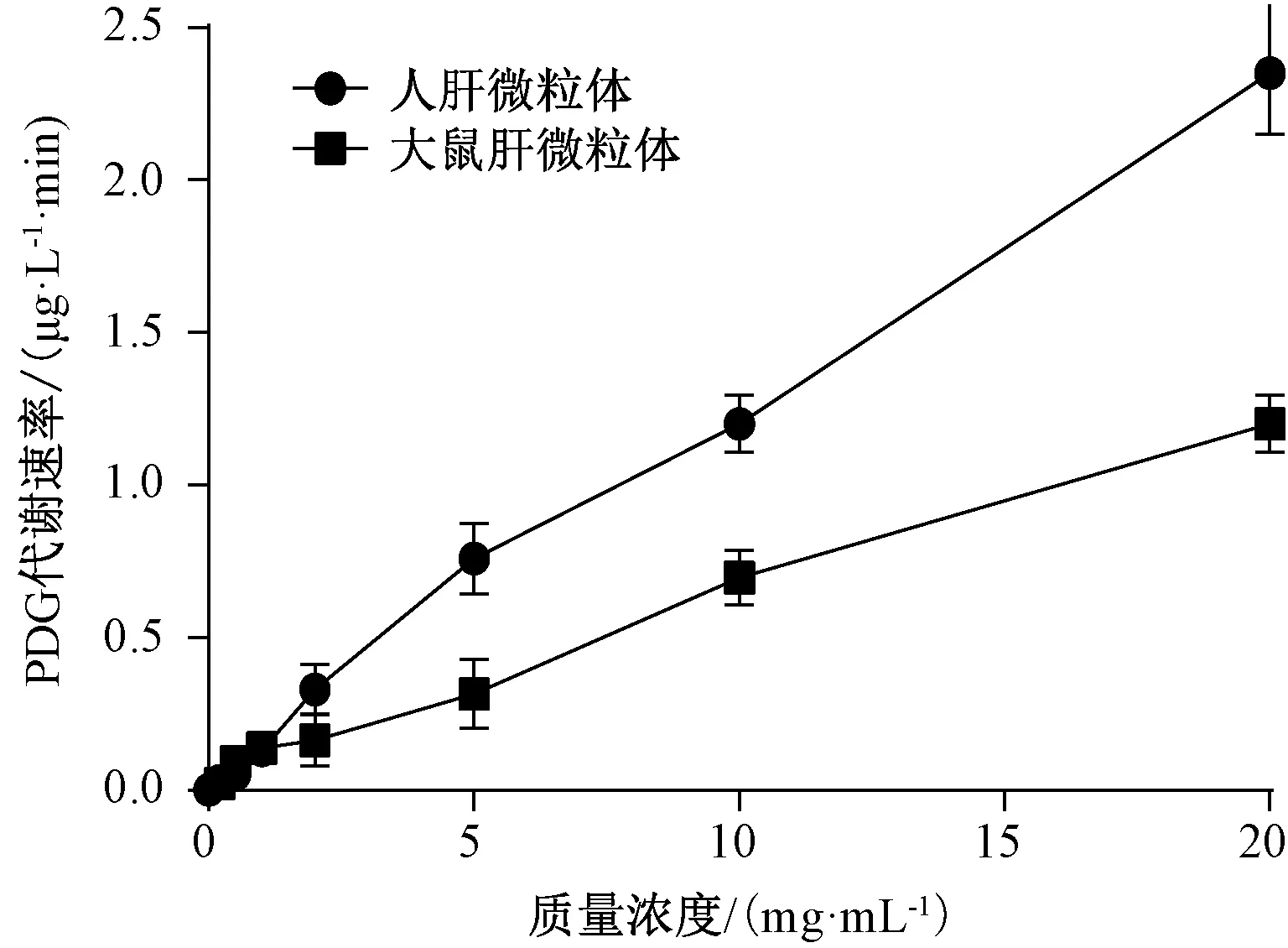
图1 肝微粒体质量浓度对代谢速率的影响Fig.1 Effect of liver microsome concentration on metabolic rate
2.7 温孵时间对松脂醇二葡萄糖苷代谢率的影响 在体外温孵体系中加入人肝微粒体和SD大鼠肝微粒体,质量浓度分别为21.9、20 mg/mL,松松脂醇二葡萄糖在37 ℃下分别孵育0、5、15、30、45、60、90、120 min后检测其糖苷质量浓度,发现在90~120 min时趋于稳定,最终确定孵育时间为120 min。再以被代谢量和加入量的比值计算松脂醇二葡萄糖苷代谢率,测得在人和SD大鼠肝微粒体中其代谢率分别为9.4%和4.8%,见图2。
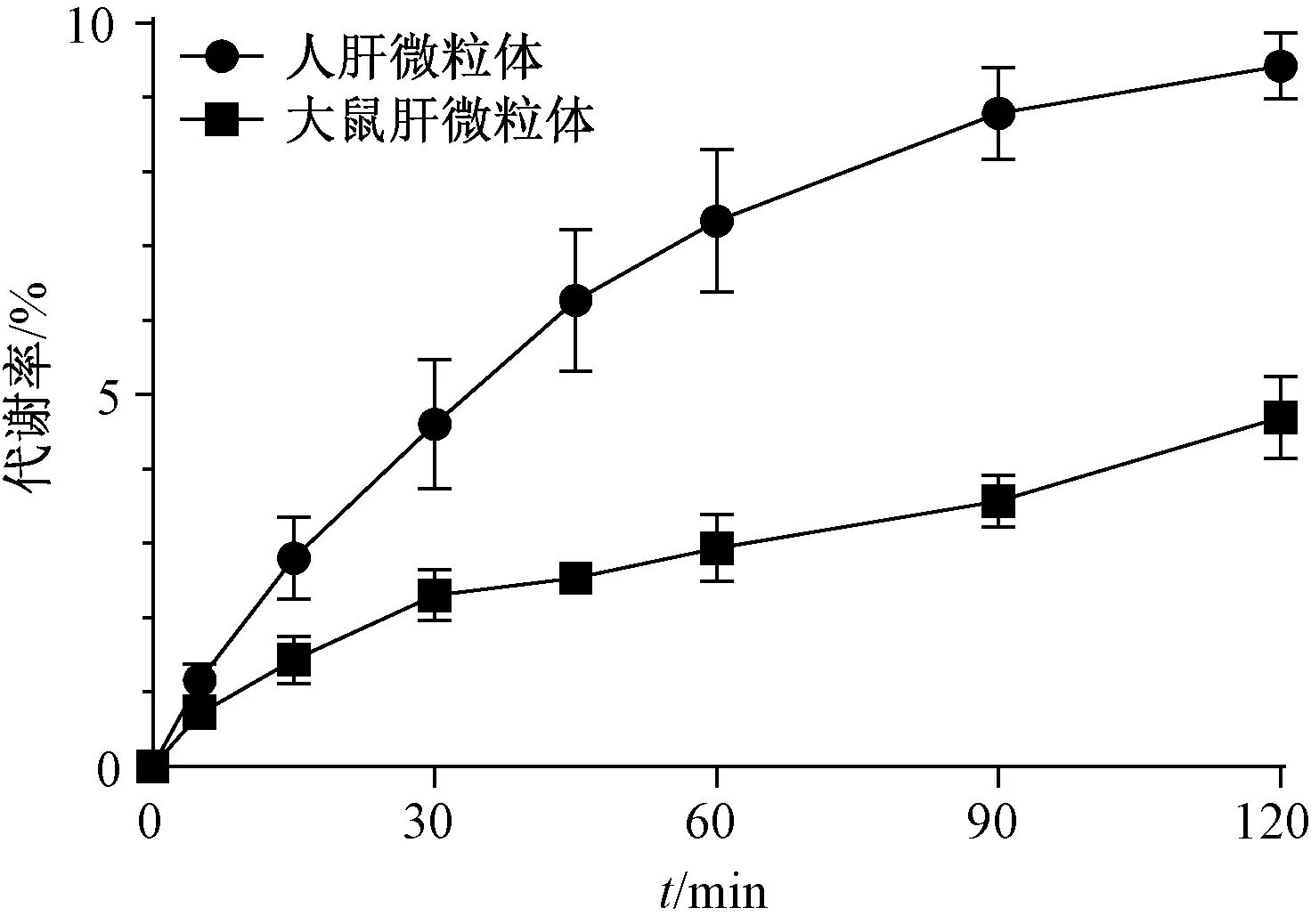
图2 温孵时间对松脂醇二葡萄糖苷代谢率的影响Fig.2 Effect of incubation time on the metabolic rate of pinoresinol diglucoside
2.8 质谱分析 松脂醇二葡萄糖苷保留时间为4.8 min,在m/z681.3处形成准分子离子[M-H]-,其二级质谱图见图3。由此可知,m/z519.3为m/z681.3断裂1个糖苷键形成,而m/z357.3为后者断裂2个糖苷键形成,特征峰强度最高。

图3 松脂醇二葡萄糖苷二级质谱图Fig.3 Secondary mass spectrum of pinoresinol diglucoside
2.9 人肝微粒体中代谢产物分析 根据保留时间、分子离子峰、二级质谱信息对特征峰进行确认,初步确认松脂醇二葡萄糖在人肝微粒体中生成6个主要代谢产物,分别命名为M1(m/z357.3)、M2(m/z345.4)、M3(m/z331.4)、M4(m/z329.2)、M5(m/z301.4)、M6(m/z297.3),总离子流图见图4。
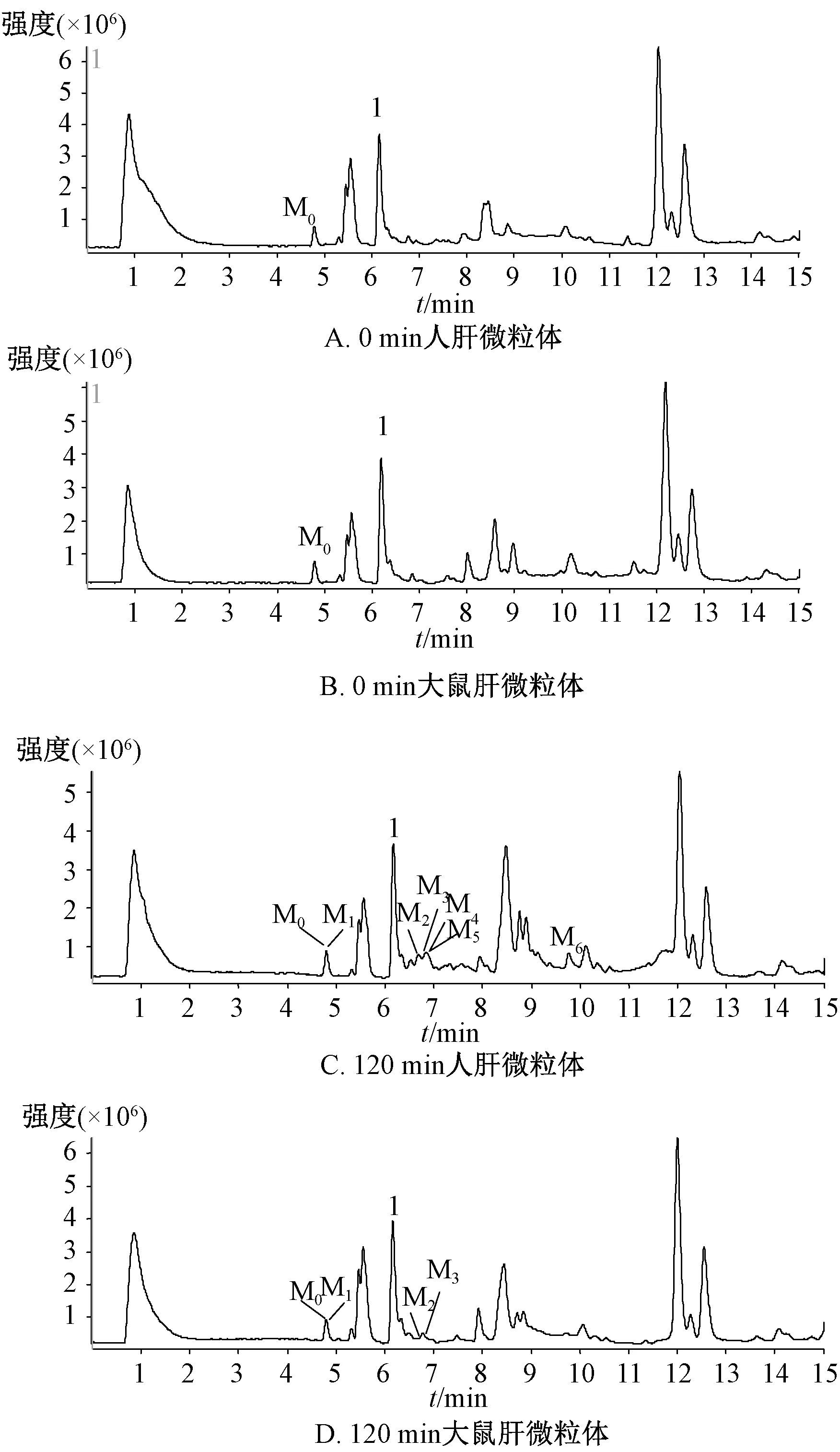
图4 松脂醇二葡萄糖苷总离子流图 Fig.4 Total ion current chromatograms ofpinoresinol diglucoside
M1在保留时间4.8 min处有1个m/z357.3的准分子离子峰[M-H]-,峰质量数比底物松脂醇二葡萄糖[M-H]-少324 Da,由于该底物包含2个糖苷键,故推测m/z357.3可能为底物脱去2个糖基形成;底物离子681.3与M1的主要碎片离子峰相同,均为m/z151.1[M-H-C12H14O3]-,二级质谱图见图5A。
M2在保留时间6.67 min处有1个m/z345.4的准分子离子峰[M-H]-,主要碎片离子[M2-H-H2O]-m/z327.3,推测其可能为m/z359.3发生氧脱甲基反应得到;在m/z357.3[M-H]-峰的基础上加2个氢还原,即1个呋喃环开环生成m/z359.3[M-H]-峰,主要碎片离子为m/z257.2、152.3,但它在反应液里仅检测到少量,可能很容易发生脱甲基反应,进而生成m/z345.4,二级质谱图见图5B。
M3在保留时间6.74 min处有1个m/z331.4的准分子离子峰[M-H]-,它是m/z357.3[M-H]-发生2个呋喃环开环、1个氧脱甲基、1个脱羟基反应得到,主要碎片离子m/z313.3[M3-H-H2O]-,二级质谱图见图5C。
M4在保留时间6.76 min处有1个m/z329.2的准分子离子峰[M-H]-,是m/z357.3[M-H]-峰发生2个呋喃环开环、脱氢形成内酯环、2次氧脱甲基反应而得,主要碎片离子为m/z201.1[M4-110-18-H]-,即m/z201.1[M4-H-C6H6O2-H2O]-,二级质谱图见图5D。
M5在保留时间6.93 min处有1个m/z301.1的准分子离子峰[M-H]-,在M1结构上发生环氧开环、2次氧脱甲基、2次脱羟基反应,得到m/z301.4[M-H]-峰,碎片离子m/z265.1[M5-H-36]-,即[M5-H-2H2O]-;碎片离子m/z221.0[M5-36-44-H]-,即[M5-2H2O-COOH]-,二级质谱图见图5E。
M6在保留时间9.79 min处有1个m/z297.3的准分子离子峰[M-H]-,在1个m/z301.4[M-H]-的基础上脱去4个氢形成内酯环,得到m/z297.3[M-H]-峰,主要碎片离子m/z250.9[M6-46-H]-,即[M6-HCOOH-H]-,二级质谱图见图5F。
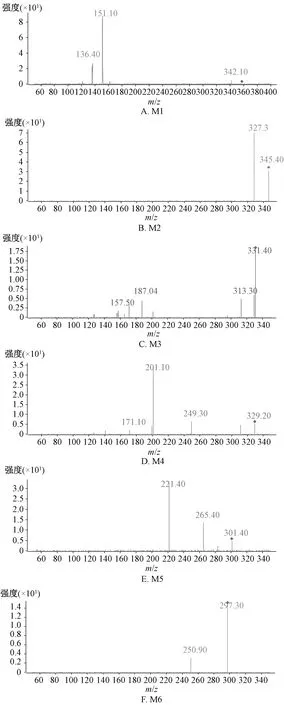
图5 松脂醇二葡萄糖苷代谢产物二级质谱图Fig.5 Secondary mass spectra for metabolites of pinoresinol diglucoside
2.10 松脂醇二葡萄糖苷代谢途径分析
2.10.1 人肝微粒体 见图6。
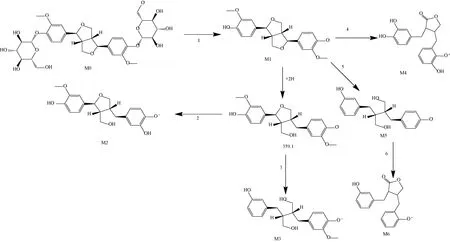
注:1为脱糖,2为加氢、脱甲基,3为加氢、脱甲基、脱羟基,4为开环、脱氢、脱甲基,5为开环、脱甲基、脱羟基,6为脱氢成环。图6 松脂醇二葡萄糖苷在人肝微粒体中的代谢途径Fig.6 Metabolic pathways of pinoresinol diglucoside in human liver microsomes
2.10.2 大鼠肝微粒体 松脂醇二葡萄糖苷代谢产物有3种,分别为M1、M2、M3,与在人肝微粒中的代谢存在差异。
3 讨论与结论
本实验建立了灵敏、快速的HPLC-QQQ/MS法来测定人和大鼠肝微粒体中松脂醇二葡萄糖苷及其代谢产物,分别初步确定了6个和3个,均以脱糖基反应为主,并且生成的苷元松脂素进一步发生还原、氧脱甲基、开环等一系列反应。通常认为,药物代谢过程可使化合物极性增大,从而促进药物排泄,但也有很多例外[12],例如在肠道菌群的作用下皂苷会发生结构变化,主要是逐步脱糖过程,生成的转化产物比原皂苷具有更好的生物利用度或更强的生物活性[13]。Kim等[14]发现,人参皂苷酶促转化只发生去糖基化;Zhai等[15]报道,黄芪甲苷生物利用度很低,需要在肠道菌群的代谢作用下产生次生皂苷才能大大提高。
本实验所得结果与Xie等[10]报道比较,松脂醇二葡萄糖苷在肝微粒体中的代谢产物数量较少,推测其在肝脏中的专属代谢酶种类可能少于在肠道菌群中的生物酶,但主要代谢部位尚无法确定。松脂醇二葡萄糖苷属于木脂素类化合物,在体内主要发生Ⅰ相代谢转化, 即水解、氧化、去甲基化或羟基化等[16],而且本实验也只发现了该类代谢产物,尚未找到II相反应相关产物。
综上所述,本实验探讨了松脂醇二葡萄糖苷在人和大鼠肝微粒体中的转化规律,推测了主要转化途径及转化产物,可为进一步研究该成分体内代谢过程提供实验依据。
猜你喜欢
中国药业(2022年18期)2022-10-12
中国民族民间医药(2022年10期)2022-07-27
中国现代中药(2022年6期)2022-07-06
阅读(科学探秘)(2021年8期)2021-09-01
发明与创新·中学生(2021年8期)2021-08-24
发明与创新(2021年30期)2021-07-22
科教新报(2021年14期)2021-05-11
教育周报·教育论坛(2020年3期)2020-10-21
科技资讯(2018年16期)2018-10-26
科技信息·下旬刊(2018年8期)2018-10-21
