
VIGS结合HIGS诱导的烟草对斜纹夜蛾与棉铃虫的抗性比较
2022-02-08 08:20农知旺谢锋华陈宇辰郭珍珍竺锡武
湖南农业科学 2022年12期
农知旺,谢锋华,陈宇辰,吴 娟,2,郭珍珍,2,竺锡武,2
(1. 湖南人文科技学院农业与生物技术学院,湖南 娄底 417000;2. 娄底市农业科学研究所,湖南 娄底 417000)
斜纹夜蛾与棉铃虫都是世界上分布广泛的农业害虫,可为害作物300种以上,包括烟草、棉花和十字花科蔬菜等重要的经济作物,危害甚大,如何有效控制对斜纹夜蛾和棉铃虫的发生为害是重要的科学问题[1-4]。RNA沉默技术快速发展[5-6],为斜纹夜蛾和棉铃虫的防控提供了一条新途径。病毒诱导的基因沉默(virus induced gene silencing,VIGS)是以RNA干扰为基础,主要用于研究动植物基因功能的一种技术,属于转录后基因沉默(post-transcriptional gene silencing,PTGS)[7-9]。寄主诱导的基因沉默 (Host induced gene silencing,HIGS) 是以RNA干扰为基础的进一步发展,近十年来研究应用于提高植物抗病虫害的性能[10-14],展现了很好的前景。有文献报道沉默棉铃虫一个定位于线粒体的 NDPH 脱氢酶黄素蛋白2(NDUFV2,NV2)基因能够抑制棉铃虫的生长发育[15]。笔者利用昆虫NV2基因片段构建的重组病毒质粒,通过VIGS和HIGS诱导的烟草植株对斜纹夜蛾与棉铃虫的抗性进行比较研究,分析对害虫的抗性是否有差异,以期为应用植物病毒载体防治斜纹夜蛾及棉铃虫,降低斜纹夜蛾及棉铃虫的发生程度提供理论依据。
1 材料与方法
1.1 试验材料
pCB-CMVF209-2b61aa、pCB301-CMVF209-no2b、pCB-CMVF109、pCB-CMVF209、pCB-CMVF309、pTRV1、pTRV2、pCB301-CMVF209-2b61aa-SliNV2370、pCB301-CMVF209-no2b-GFP350、pCB301-CMVF209-no2b-SliNV2370、pCB301-CMVF209-no2b-HaNV2370、pTRV2-GFP350、pTRV2-SliNV2370、pTRV2- HaNV2370,由实验室保存或构建。本氏烟种子由实验室保存。
1.2 主要试剂
rTaq DNA聚合酶、 qPCR Mix (SYBR Green I)由Takara公司购得;TRIzol购自Thermo公司;2-(N-吗啡啉)乙磺酸(MES)、乙酰丁香酮(AS)、卡那霉素(Kana)、利福平(Rif)等试剂购自杭州兰堡海博生物技术有限公司。
1.3 主要仪器与引物序列
主要仪器为PCR仪(美国);荧光定量PCR仪(美国);凝胶成像系统(美国)。引物序列以斜纹夜蛾和棉铃虫的NV2及actin基因序列设计RT-qPCR引物(表1),由北京擎科生物公司合成。

表1 PCR引物序列
1.4 方 法
1.4.1 重组病毒在本氏烟植株表达SliNV2370或HaNV2370片段的试验设计 采用重组病毒质粒-农杆菌浸润法在本氏烟烟草植株4~6叶期接种,使病毒载体及其外源基因片段在烟草植株中表达,其具体步骤参照吴娟[16]的研究报道。
试验设计一:利用pCB-CMVF209-2b61aa-SliNV2370重组病毒质粒在本氏烟植株中表达SliNV2-370基因片段。用pCB-CMVF209-2b61aa空载体和不接种病毒的本氏烟作对照。每个处理12钵,每钵3株苗。具体设计如下:(1)pCB-CMVF109+ pCB-CMVF309+ pCBCMVF209-2b61aa-SliNV2370,(2)pCB-CMVF109+pCB-CMVF309+ pCB-CMVF209-2b61aa,(3)Mock。
试验设计二: 利用pCB-CMVF209-no2b-SliNV2370重组病毒质粒在本氏烟植株中表达SliNV2-370基因片段。用pCB-CMVF209-no2b-GFP350和不接种病毒的本氏烟作对照。每个处理12钵,每钵3株苗。具体设计 如 下:(1)pCB-CMVF109+ pCB-CMVF309+ pCBCMVF209-no2b-SliNV2370,(2)pCB-CMVF109+ pCBCMVF309+ pCB-CMVF209-no2b-GFP350,(3)Mock。
试验设计三:利用pTRV2-SliNV2370重组病毒质粒在本氏烟植株中表达SliNV2-370基因片段。用pTRV2-GFP350和不接种病毒的本氏烟作对照。每个处理12钵,每钵3株苗。具体设计如下。(1)pTRV1+pTRV2-SliNV2370,(2)pTRV1+ pTRV2-GFP350,(3)Mock。
试验设计四:利用pCB-CMVF209-no2b-HaNV2370重组病毒质粒在本氏烟植株中表达HaNV2-370基因片段。用pCB-CMVF209-no2b-GFP350和不接种病毒的本氏烟作对照。每个处理12钵,每钵3株苗。具体设计 如 下:(1)pCB-CMVF109+ pCB-CMVF309+ pCBCMVF209-no2b-HaNV2370,(2)pCB-CMVF109+ pCBCMVF309+ pCB-CMVF209-no2b-GFP350,(3)Mock。
试验设计五:利用pTRV2-HaNV2370重组病毒质粒在本氏烟植株中表达HaNV2-370基因片段。用pTRV2-GFP350和不接种病毒的本氏烟作对照。每个处理12钵,每钵3株苗。具体设计如下:(1)pTRV1+pTRV2-HaNV2370,(2)pTRV1+ pTRV2-GFP350,(3)Mock。
1.4.2 本氏烟烟草植物总RNA 的提取 取本氏烟接种12 d后的完全展开的非接种叶片提取总RNA,具体方法参照Trizol 试剂说明书,具体操作步骤参考吴娟[16]的研究报道。
1.4.3 目的基因在本氏烟植株中的表达情况 提取RNA 后,RT-PCR检测目的基因是否在本氏烟中表达,检测重组病毒质粒是否在本氏烟上接种成功,用重组病毒质粒构建时相对应的引物进行检测,具体步骤参考吴娟的研究报道[16]。
1.4.4 接种重组病毒的本氏烟植株对斜纹夜蛾和棉铃虫的抗性试验 RT-PCR 检测接种重组病毒成功的本氏烟植株用于饲养斜纹夜蛾和棉铃虫试验,试验处理与浸润接种病毒的本氏烟处理相对应。以一龄的斜纹夜蛾或棉铃虫作为试验对象,每个处理分别设置4个重复,每个培养皿为一次重复。将一龄的斜纹夜蛾或棉铃虫饥饿2 h后,选取重量、大小、生命活力基本一致的一龄幼虫分别放入每个试验皿中,每个试验皿各4头幼虫(由于棉铃虫存在自相残杀习性,故将试验皿分成四小格,每个小格里面放一头棉铃虫)。分别选取相对应处理的本氏烟植株相同叶位的叶子喂食。试验在温室中进行,温度设置为25℃,光照时间设置为16 h;黑暗时间为8 h,湿度60%,每隔3 d称重一次斜纹夜蛾或棉铃虫的重量。试验结束前,收集各处理的幼虫,称重后用液氮处理保存于-80℃。
1.4.5 供试幼虫NV2基因的表达情况 取-80℃保存的幼虫样品提取总RNA,并以提取的RNA 为模板去除基因组DNA。以设计的RT-qPCR引物,按照RTqPCR仪器说明书步骤开展RT-qPCR,并参考吴娟[16]的具体步骤。
1.4.6 数据处理 所有试验数据用SPSS软件进行方差分析(Duncan法多重比较)。
2 结果与分析
2.1 重组病毒质粒表达SliNV2370基因片段的本氏烟植株对斜纹夜蛾的抗性
2.1.1 各处理重组病毒及其插入的基因片段在本氏烟中的表达 以提取的RNA 为模板,以CMVF209序列设计的通用引物2bNcoIF2和2bAvrⅡR对各CMV重组质粒的试验处理样品作RT-PCR 检测,结果如图1A、B所示,PCR 产物大小与预期大小相符。表明各重组病毒CMV2b61aa、CMV2b61aa-SliNV2370、CMVno2b-GFP350、CMVno2b-SliNV2370已 经 分 别 在相应试验处理的本氏烟植株中成功表达。以TRV RNA2序列设计的通用引物TRV2F1和TRV2R1对各TRV重组质粒的试验处理样品做RT-PCR 检测,结果如图1C所示,表明重组病毒TRV2-GFP350、TRV2-SliNV2370在相应试验处理的本氏烟植株中成功表达。
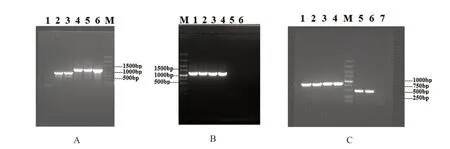
图1 RT-PCR检测目的病毒及其外源基因片段的电泳结果
2.1.2 重组病毒质粒表达SliNV2370基因片段的本氏烟植株对斜纹夜蛾的抗性 试验结果如图2所示。图2A 所示CMV2b61aa-SliNV2370表达SliNV2370的试验结果。分析表明,pCB-CMVF209-2b61aa-SliNV2370处理与mock处理、pCB-CMVF209-2b61aa处理的斜纹夜蛾的平均每头幼虫重量分别相差3%、13%,但方差分析差异都不显著。图2B 所示CMVF209-no2b-SliNV2370表达SliNV2370的试验结果。分析表明,pCBCMVF209-no2b-SliNV2370处理与mock处理之间的斜纹夜蛾的平均每头幼虫重量相差30%,差异显著;与pCB-CMVF209-no2b-GFP350处理之间的斜纹夜蛾的平均每头幼虫重量分别相差23%,但方差分析差异不显著。图2C所示TRV-SliNV2370表达SliNV2370的试验结果。分析表明,pTRV2-SliNV2370处理与mock处理、pTRV2-GFP350处理的斜纹夜蛾的平均每头幼虫重量分别相差16%、15%,方差分析差异都不显著。
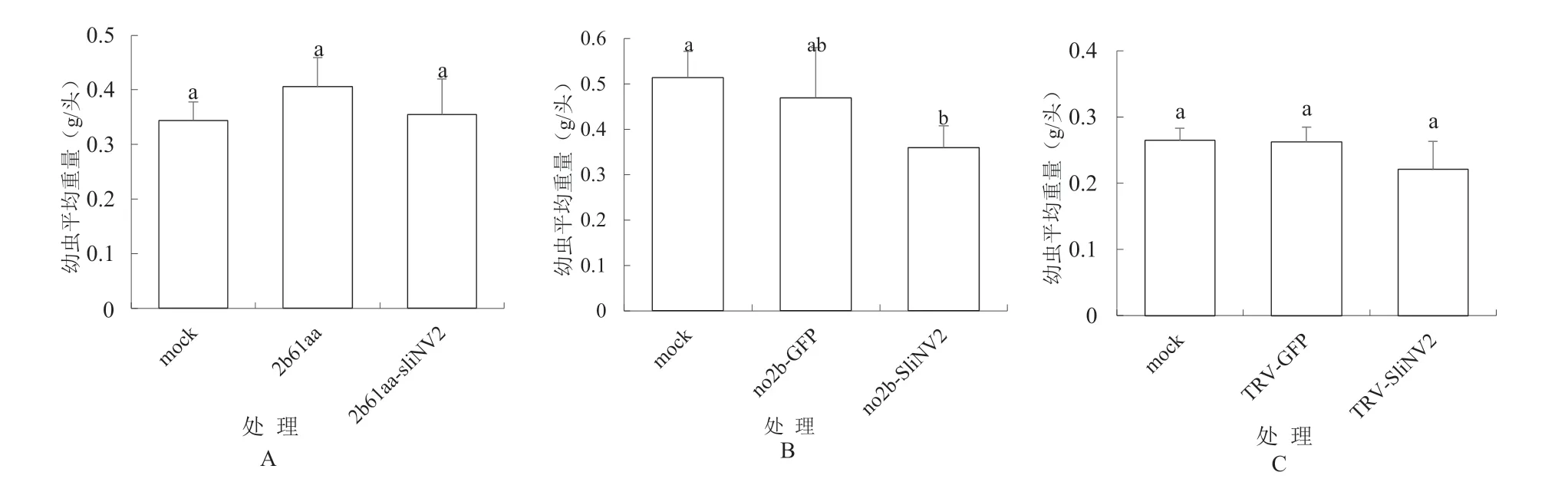
图2 试验各处理的本氏烟植株对斜纹夜蛾的抗性结果
2.1.3 RT-qPCR检测斜纹夜蛾NV2基因表达结果RT-qPCR分析结果如图3所示。图3A所示,pCBCMVF209-no2b-SliNV2370处理中的斜纹夜蛾的NV2基因的表达水平分别是mock处理和pCB-CMVF209-no2b-GFP350处理的0.746和0.801倍,方差分析差异显著,表明pCB-CMVF209-no2b-SliNV2370处理的斜纹夜蛾的NV2基因表达量相对于pCB-CMVF209-no2b-GFP350空载体处理显著性地下调了19.9%。图3B所示,pTRV2-SliNV2370处理中的斜纹夜蛾的NV2基因的表达水平分别是mock处理和pTRV2-GFP350处理的0.763和0.807倍,方差分析差异显著。结果表明pTRV2-SliNV2370处理的斜纹夜蛾的NV2基因表达量相对于pTRV2-GFP350空载体处理显著性地下调了19.3%。但按转录组测序筛选差异表达基因的标准[17][差异倍数大于2,即log2(FoldChange) > 1]判定,两类病毒载体表达SliNV2基因片段试验处理与空载体处理的斜纹夜蛾SliNV2基因表达差异都没有达到差异表达基因的水平。
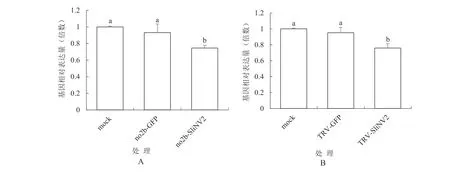
图3 斜纹夜蛾NV2基因表达量的RT-qPCR分析
2.2 重组病毒表达HaNV2370基因片段的本氏烟植株对棉铃虫的抗性
2.2.1HaNV2370基因片段在本氏烟植株中的表达结果 以CMV F 209序列设计的通用引物2bNcoIF2和2bAvrⅡR对各CMV重组质粒的试验处理样品作RT-PCR检测,结果如图4A所示,PCR产物大小与预期大小相符,表明病毒载体pCB-CMVF209-no2b-GFP350、pCB-CMVF209-no2b-HaNV2370已经在本氏烟中表达。用TRV RNA2序列设计的引物TRV2F1 和TRV2R1 对TRV重组质粒的试验处理样品作RT-PCR检测,结果如图4B所示,PCR 产物大小与预期大小相符,表明重组病毒TRV-GFP350、TRV-HaNV2370已经在相应试验处理的本氏烟植株中成功表达。
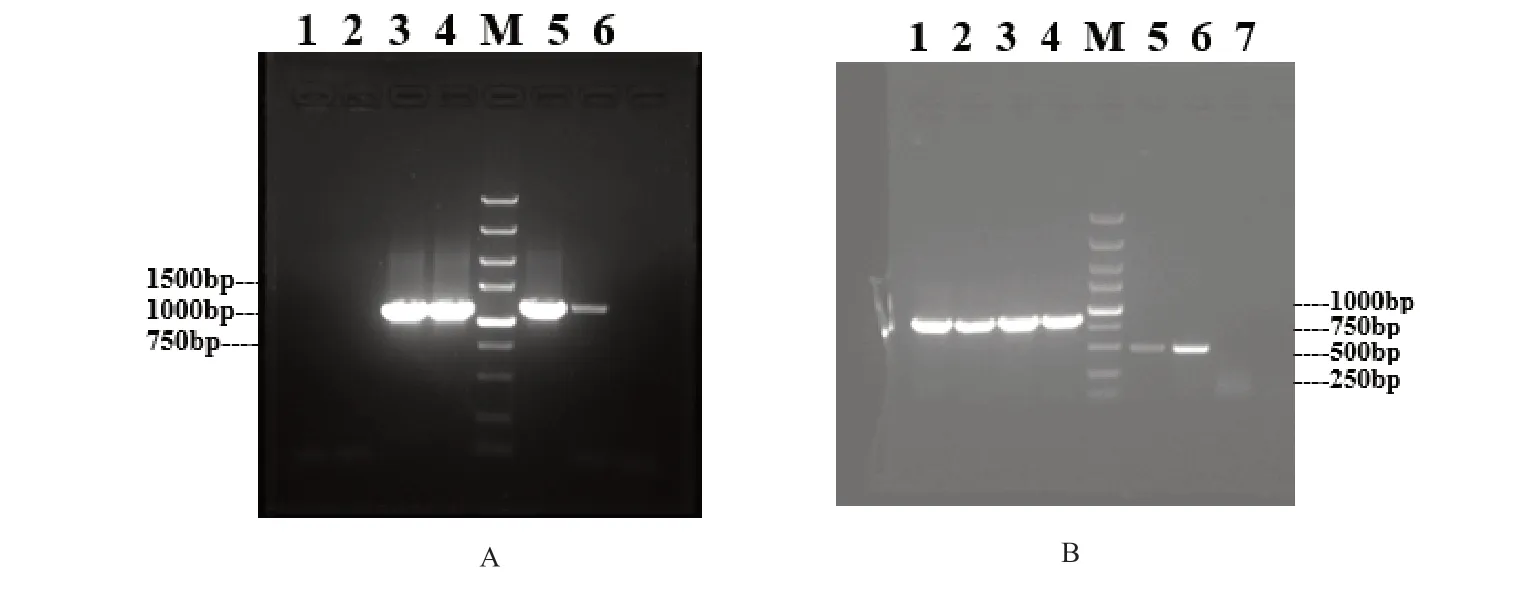
图4 RT-PCR检测试验中目的病毒及其外源基因片段的电泳结果
2.2.2 重组病毒质粒表达HaNV2370基因片段的本氏烟植株对棉铃虫的抗性 试验结果如图5所示。图5A所示CMVno2b-HaNV2370表达HaNV2370的试验结果。分析表明,pCB-CMVF209-no2b-HaNV2370处理与mock处理、pCB-CMVF209-no2b-GFP350处理之间的斜纹夜蛾的平均每头幼虫重量分别相差35%和38%,方差分析差异显著。图5B所示TRV2-HaNV2370表达HaNV2370的试验结果,分析表明,TRV2-HaNV2370处理与mock、TRV2-GFP350处理之间的棉铃虫的平均每头幼虫重量分别相差33%和30%,方差分析差异显著。
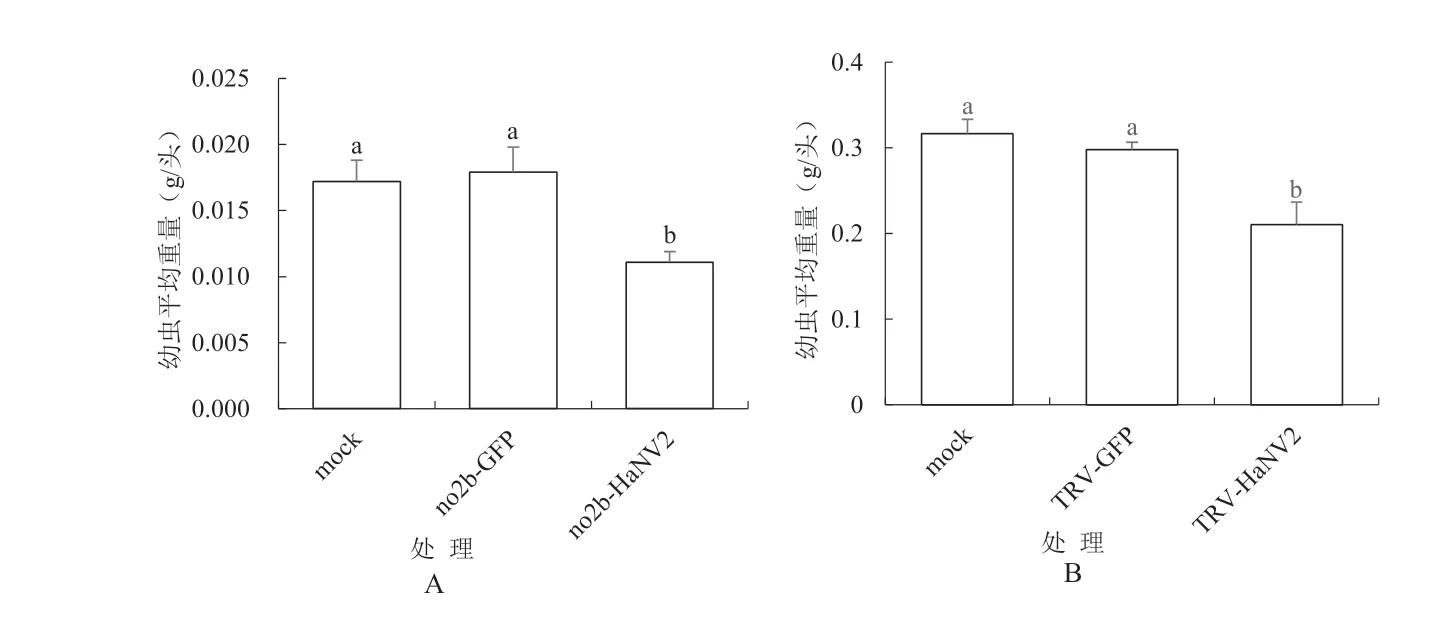
图5 试验中各处理的本氏烟植株对棉铃虫的抗性结果
2.2.3 RT-qPCR检测棉铃虫NV2基因的表达结果RT-qPCR分析结果如图6所示。图6A所示,pCBCMVF209-no2b-HaNV2370处理中的棉铃虫的NV2基因的表达水平分别是mock处理和pCB-CMVF209-no2b-GFP350处理的0.248和0.275倍。结果表明pCBCMVF209-no2b-HaNV2370处理的棉铃虫的NV2基因表达量相对于pCB-CMVF209-no2b-GFP350处理下调了72.5%。图6B所示,pTRV2-HaNV2370处理中的棉铃虫的NV2基因的表达水平分别是mock处理和pTRV2-GFP350处理的0.203和0.213倍。表明TRV-HaNV2370处理的棉铃虫的NV2基因表达量相对于pTRV2-GFP350下调了78.7%。按转录组测序筛选差异表达基因的标准[17][差异倍数大于2,即log2(FoldChange) > 1]判定,两类病毒载体表达HaNV2基因片段试验处理与空载体处理比较,试验中的棉铃虫HaNV2基因表达差异都达到了差异表达基因的水平。
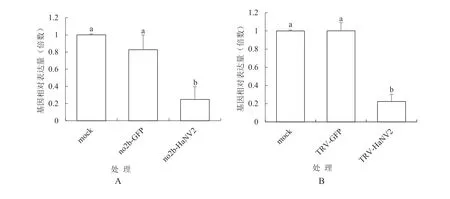
图6 棉铃虫NV2基因表达量的RTq-PCR分析
3 小结与讨论
通过重组病毒质粒-农杆菌浸润接种本氏烟植株,表达SliNV2370或HaNV2370基因片段的本氏烟植株分别用于饲养斜纹夜蛾、棉铃虫试验,结果表明,通过CMV载体或TRV载体表达SliNV2370基因片段的本氏烟植株都不能显著增强对斜纹夜蛾的抗性;CMV载体与TRV载体分别表达HaNV2370基因片段的本氏烟植株都能显著增强对棉铃虫的抗性。RTq-PCR分析,各处理斜纹夜蛾的NV2基因表达差异都没有达到差异表达基因的水平;棉铃虫相关的试验中,CMV载体或TRV载体表达HaNV2370基因片段的处理与空载体处理棉铃虫NV2基因表达差异达到了差异表达基因的水平,与抗性试验结论具有一致性,验证了抗性试验结果的正确性。
在发现3种重组病毒质粒表达SliNV2370基因片段的本氏烟植株都没有显著性地增强对斜纹夜蛾的抗性后,通过CMVno2b载体、TRV载体开展了抗棉铃虫试验,与对斜纹夜蛾的抗性试验结果比较,以确认对斜纹夜蛾的抗性试验结果的真实性、正确性,确认表达SliNV2370基因片段不能显著增强本氏烟对斜纹夜蛾的抗性是否与载体不同有关?是否与试验操作有关?是否与不同昆虫种类有关?不同载体在本氏烟植株中表达NV2基因片段都不能增强本氏烟植株对斜纹夜蛾的抗性,但都能增强对棉铃虫的抗性,表明抗性差异与病毒载体无关,也与试验操作无关,可能与昆虫种类不同有关。
此研究病毒载体结合HIGS诱导的植株对棉铃虫的抗性效果没有毛颖波等[18]报道的效果好,但其抗棉铃虫趋势是相同的。毛颖波等[18]的研究采用了两对引物分别表达HaNV2基因不同片段的寄主植物对棉铃虫的抗性效果,此研究只采用了其中的一对引物可能是其原因之一,原因仍有待进一步分析和试验证实。表达HaNV2基因的本氏烟植株饲养棉铃虫能降低78.7%的棉铃虫NV2基因表达量,这与毛颖波[18]等报道的基本一致。用成功表达SliNV2基因的本氏烟饲养斜纹夜蛾只能相对降低斜纹夜蛾NV2基因表达量19.3%,虽然方差分析结果显示差异显著,但按转录组测序筛选差异表达基因的标准判定,SliNV2基因表达量没有达到基因差异表达水平,因而表观上表现为不能显著性地增强本氏烟植株对斜纹夜蛾的抗性。这可能是HIGS方法不适合应用于沉默斜纹夜蛾基因。已有报道的沉默斜纹夜蛾基因较好的方法都是微注射法[19-20]。并且另有报道,转Bt基因植物对棉铃虫抗性好而对斜纹夜蛾抗性差,是由于斜纹夜蛾的中肠酶液将Cry1Ac蛋白降解为60 kDa的片段大小和水解为38 kDa的片段大小,Cry1Ac蛋白的过度降解导致杀虫活性降低[21],这一研究结果表明斜纹夜蛾具有不同于棉铃虫等鳞翅目害虫的特异的生理生化特性。VIGS结合HIGS方法对斜纹夜蛾幼虫SliNV2基因的沉默效果不佳可能是因为斜纹夜蛾物种由其生理生化特异性导致。总之,病毒载体表达NV2-370基因片段的本氏烟植株对斜纹夜蛾、棉铃虫的抗性有明显差异的原因,可能是因为斜纹夜蛾与棉铃虫两个不同的物种的幼虫有生理生化差异,具体还需后续进一步研究。
猜你喜欢
少儿科学周刊·少年版(2022年10期)2022-06-27
云南农业(2021年11期)2021-11-12
今日农业(2020年13期)2020-12-15
农药科学与管理(2019年6期)2019-11-23
今日农业(2019年13期)2019-08-12
今日农业(2019年10期)2019-06-26
农民致富之友(2019年2期)2019-03-16
女友·家园(2016年4期)2016-04-27
安徽农业科学(2015年10期)2015-12-17
小学科学(2015年9期)2015-09-28
