
不同苗龄云南松异速生长及其表型可塑性
2022-02-23 07:29李江飞李亚麒颜廷雨诗a许玉兰蔡年辉
中南林业科技大学学报 2022年1期
王 丹,李江飞,李亚麒,颜廷雨,陈 诗a,许玉兰,蔡年辉
(1.西南林业大学 a.西南山地森林资源保育与利用教育部重点实验室;b.云南省高校林木遗传改良与繁育重点实验室,云南 昆明 650224;2.云南省农业科学院 热带亚热带经济作物研究所,云南 保山 678000)
随着植物的生长发育和资源可获得性的改变,植物会对现有资源的利用策略做出相应的改变[1],并以异速生长的方式来获取所需资源和对环境的适应[2]。异速生长是生物体两个性状间不成比例生长的现象[3],能表达个体形态指标、器官生物量等生物学特征与个体大小问题的关系[4-6],能揭示植物生长的内在规律[7]。由异速生长导致植物性状的可塑性响应,被认为是植物发生适应性变化的表现,且这种响应正是对植物功能群间属性趋同或趋异的有力体现[8]。
表型可塑性的定义为同一个基因型对不同环境响应产生不同表型的特性,是发生在植物发育过程中的现象[9],被认为是植物对环境条件或刺激的重要反应特征,这一特征是植物发生适应的表型基础[10]。植物的异速生长与生物量分配对资源的利用和异质环境的形态适应有密切联系,是目前研究植物表型可塑性较为合适的方法。物种的一些表型差异,可能只是与植株大小不一,或者发育阶段不同有关,异速生长分析能把环境导致的表型差异从植物本身个体生长的影响中区分开来。通过异速生长分析研究表型可塑性,可以更好地了解植物的表型可塑性,从而进一步了解植物的适应机制[11]。
云南松Pinus yunnanensis生长迅速,喜光且耐干旱耐瘠薄,广泛分布于23°00′~29°00′N、98°30′~106°00′E 的西藏东南部、四川西南部、贵州西部、广西西部及云南大部分地区[12],是我国西南地区荒山造林的先锋树种和主要的用材树种,具有较高的生态效益和经济价值。目前,异速生长分析主要被应用于生物量分配、植物各器官的年增长率、植物对繁殖的投入等研究[13]。云南松异速生长分析也仅局限于不同器官[14]和不同地理种源[15]的研究,尚未见对不同苗龄云南松进行异速分析研究表型可塑性的研究报道。为此,本研究以不同苗龄云南松为研究对象,探明云南松随着苗龄的增加各器官生物量及针叶长的异速生长关系,有利于揭示云南松随着苗龄的增加,各器官及针叶长的生长速率变化和苗木的适应机制与生活史策略,同时可丰富云南松在生长过程中异速生长现象和表型可塑性的研究内容,为云南松高效培育提供理论参考。
1 材料与方法
1.1 研究地概况
试验地设置在西南林业大学苗圃,位于云南省昆明市盘龙区。年均温14.7℃,绝对最高温32.5 ℃,绝对最低温-9 ℃。在5—10月集中降雨,月最大降水量208.3 mm,日最大降水量153.3 mm;年平均相对湿度68.2%,年降水量700~1 100 mm。年均日照时数约2 445.6 h,年均蒸发量1 856.4 mm。终年太阳投射角度大,年均总辐射量达5.43×105J/cm2。
1.2 种子采集与处理
云南松种子的采集在云南省昆明市宜良县禄丰村林场进行,采种母树成熟、结实状况良好,采集饱满的当年生成熟球果。将采摘的球果做好标记带回实验室风干,待球果开裂后取出发育良好的种子。于2015年3月把种子点播种于苗床,株行距为5 cm×10 cm,试验采用单因素完全随机区组设计,苗期进行常规养护管理。分别于播种后第2年的12月随机选择苗木615 株、第3年12月随机选择苗木425 株、第4年12月随机选择苗木57 株作为样株,即后续分析中的2年生苗木、3年生苗木、4年生苗木。
1.3 形态指标和生物量的测定
用整株收获法挖取云南松苗木,用游标卡尺测量苗木地径,精确到0.01 mm,用直尺测量苗高、主根长、侧根长和针叶长,精确到0.1 cm。将苗木分根、茎、叶装信封袋,在105℃的烘箱中杀青30 min 后于80℃下烘干,分别记下其干物质质量,精确至0.001 g。
1.4 数据分析
异速生长关系可表示为y=axb的形式[16],线性转化为:logy=loga+blogx。在本试验中较小的苗龄有较大的样本数,因保存和取样困难等原因,较大的苗龄样本数目较小,但经对数化减弱了比例系数所带来的波动性或单纯使用生物量学特征的绝对量。式中的y与x为依赖个体大小变化的器官生物量或针叶长;a为性状关系的截距;b为异速生长指数,即线性关系的斜率,当b=1 时,为等速生长;b≠1 时,为异速生长[17]。异速生长方程的参数估计采用标准主轴法获取,并对异速生长指数和截距进行多重比较,均由统计软件R中的Smatr 包来计算完成[18-19]。若不同苗龄的斜率b差异显著,则表明异速生长关系发生了变化。
2 结果与分析
2.1 不同苗龄云南松苗木生长性状
从表1可以看出,不同苗龄云南松苗木地径、苗高和侧根长均有显著差异(P<0.05),且都是随着苗龄的增长而增长。4年生苗木主根长与2年生苗木、3年生苗木的有显著差异,但2年生苗木、3年生苗木间无显著差异,随着苗龄的增长而增长。2年生苗木的针叶长与3年生苗木、4年生苗木的有显著差异,但3年生苗木、4年生苗木间无显著差异,呈现下降趋势,表明苗木随着苗龄的增长,地径、苗高、主根长、侧根长均呈现增长趋势,而针叶长为下降趋势。

表1 不同苗龄云南松苗木生长性状Table 1 Growth characteristics of Pinus yunnanensis seedlings at different ages
从变异系数来看,地径、苗高、主根长、侧根长和针叶长苗龄均在4 a 时有最小变异系数,地径、主根长苗龄在2 a 时有最大变异系数,苗高、侧根长和针叶长苗龄在3 a 时有最大变异系数,表明苗龄为4 a 时,各指标变异性较低;苗龄为2、3 a 时,变异性较高,分化明显。
2.2 器官与个体大小的异速生长关系
不同苗龄各器官生物量与单株生物量均呈极显著正相关(P<0.01),随着苗龄的增加,根、茎、叶与单株生物量、地上生物量与根的异速生长轨迹均发生了改变(表2,图1)。从根与单株生物量来看,3年生苗木的斜率为显著小于1.0 的异速生长关系;而2年生苗木、4年生苗木的斜率为与1.0 差异不显著的等速生长关系,即3年生苗木根生物量的生长速率显著小于2年生苗木、4年生苗木。从茎与单株生物量来看,2年生苗木的斜率为显著大于1.0的异速生长关系;而3年生苗木、4年生苗木的斜率为与1.0 差异不显著的等速生长关系,即2年生苗木茎生物量的生长速率显著大于3年生苗木、4年生苗木。从叶与单株生物量来看,随着苗龄的增加,斜率为递增趋势;3年生苗木、4年生苗木的斜率为显著大于1.0 的异速生长关系,且显著大于2年生苗木,即3年生苗木、4年生苗木叶生物量生长速率显著大于2年生苗木。从地上与根的生物量来看,2年生苗木、4年生苗木的斜率为与1.0差异不显著的等速生长关系,3年生苗木的斜率为著大于1.0 的异速生长关系,且显著大于2年生苗木,即3年生苗木地上生物量的生长速率显著高于2年生苗木。
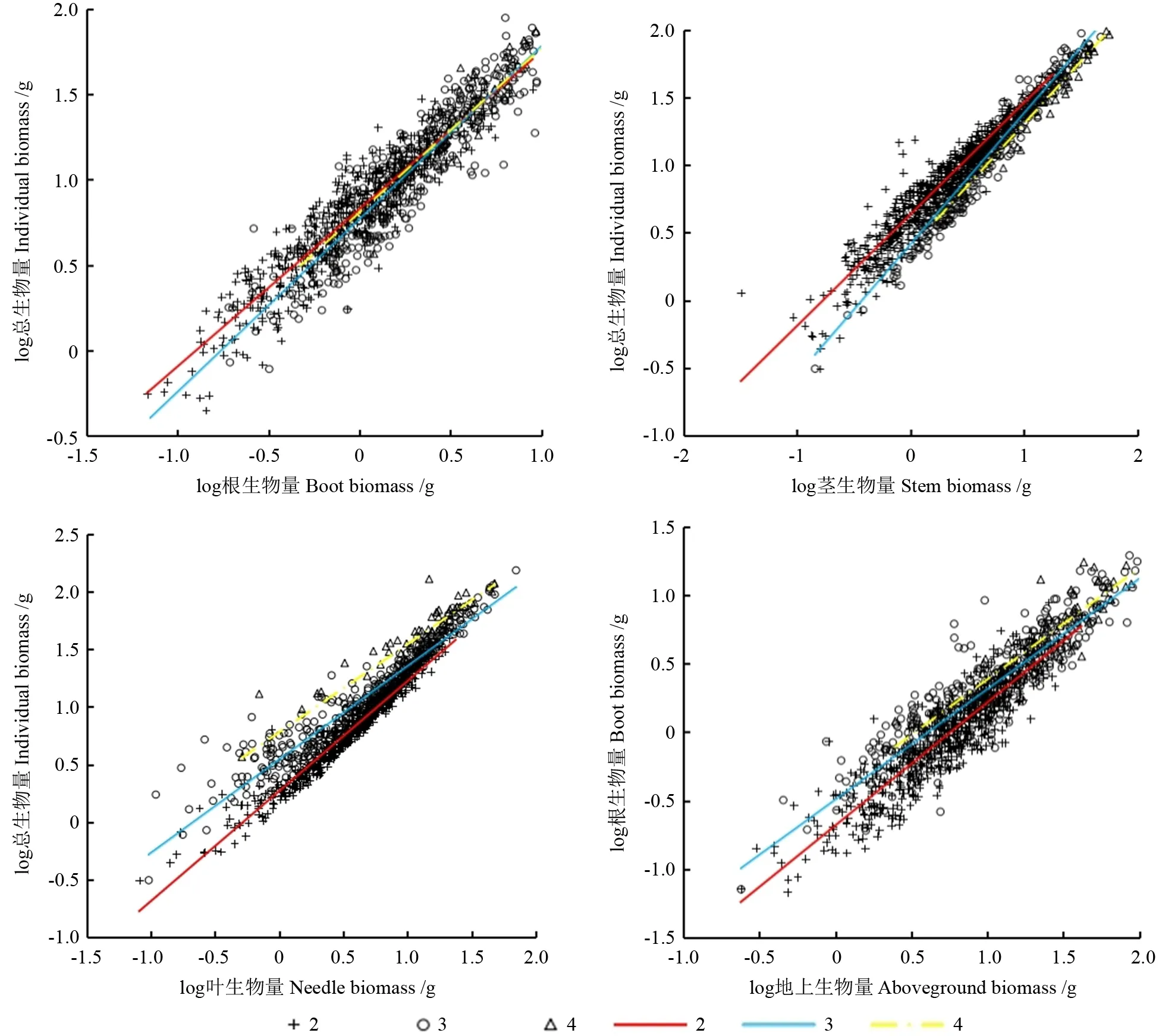
图1 器官与个体大小、地上生物量和根的异速生长关系Fig.1 Allometric relationship between organs and individual size,aboveground biomass and root
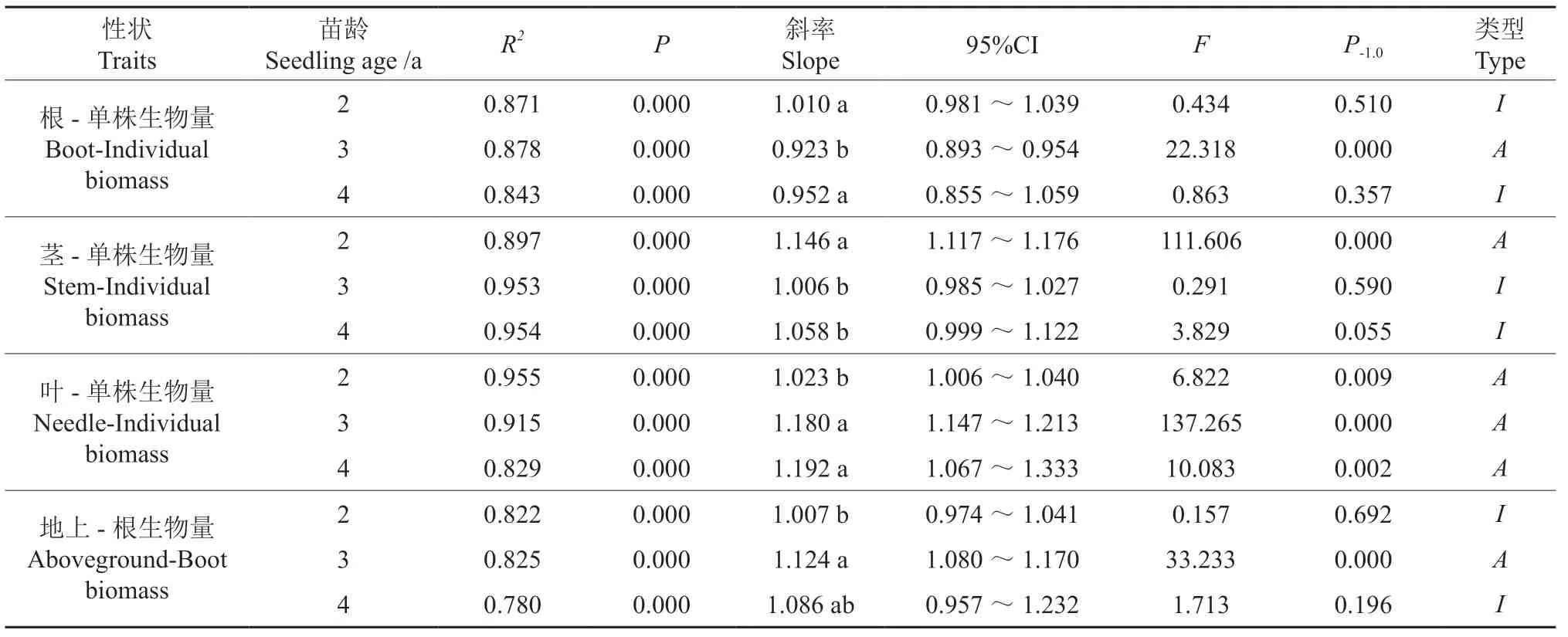
表2 不同苗龄云南松器官与个体大小、地上生物量和根的异速生长关系†Table 2 Allometric growth of organs and individual size,aboveground biomass and root of Pinus yunnanensis at different seedling ages
2.3 器官与器官、针叶长的异速生长关系
不同苗龄器官与器官、针叶长的异速生长关系均呈极显著正相关(P<0.01),各器官的生长速率在3 a 时表现为:叶>茎>根;随着苗龄的增加,除了茎与根生物量外,叶与根、茎生物量和针叶长与叶生物量的异速生长轨迹均发生了改变(表3,图2)。从茎与根生物量看,各苗龄的斜率无显著差异,共同斜率为1.113,且在共同斜率下,截距差异并不显著(P>0.05)。从叶与根生物量看,2 a 的斜率为与1.0 无显著差异的等速生长关系;3 a、4 a 为显著大于1.0 的异速生长关系,即3 a、4 a 时叶显著大于根的生长速率。从叶与茎生物量看,3 a 显著大于2 a 的斜率,且均呈异速生长关系,4 a 的斜率为与1.0 无显著差异的等速生长关系,即3 a 时叶显著大于茎的生长速率。从针叶长与叶生物量看,各苗龄的斜率均为显著小于1.0 的异速生长关系,其中4 a 显著小于2 a、3 a 针叶长生长速率,在3 a 时有较大的针叶长生长速率。
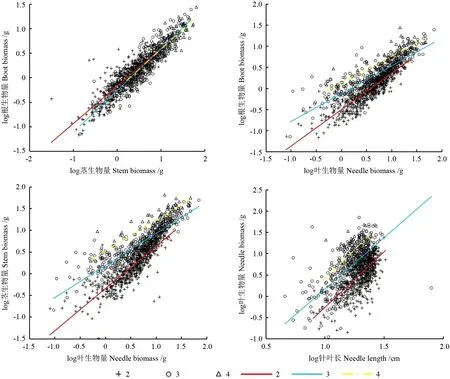
图2 器官与器官、针叶长的异速生长关系Fig.2 Allometric growth relationship among organs and needle length
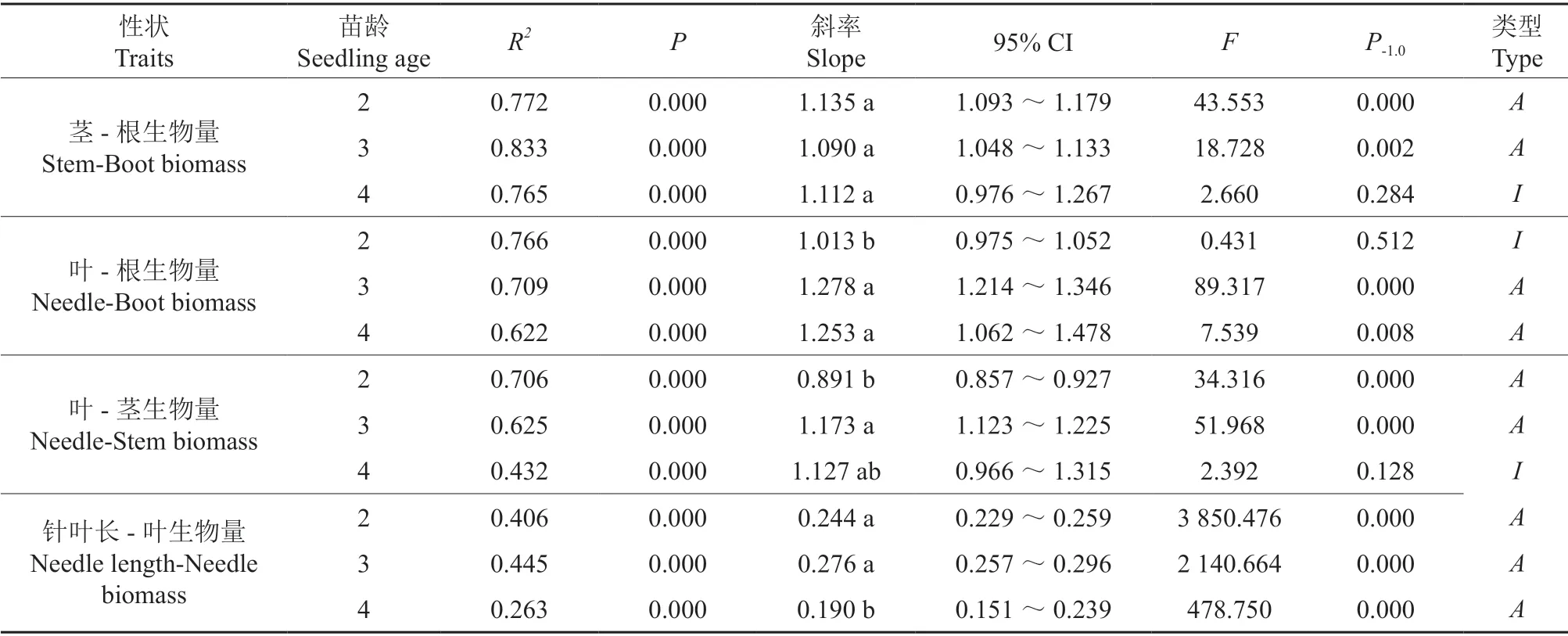
表3 不同苗龄云南松器官与器官、针叶长的异速生长关系†Table 3 Allometric growth of organs and needle length of Pinus yunnanensis at different seedling ages
P-1.0indicates the significant difference between the estimated model slope and theoretical value 1.0,and different lowercase letters show a significant difference between different seedling ages (P<0.05),Ameans the allometric relationship,Iindicates the isometric relationship.
3 结论与讨论
3.1 结 论
各苗龄云南松根、茎、叶与植株大小、地上生物量与根的异速生长轨迹均发生改变,个体发育异速关系有不同的变化趋势,可能与遗传差异或存在发育漂变有关,具有物种特异性。地上生物量与根在苗龄为3 a 时,因光成为限制资源,所以倾向于地上分配。灵活的生长对策反映了其不同苗龄资源利用策略、能量物质投入的差异。在不同生长发育阶段的重点有所不同,云南松2年生苗木倾向于茎生长,3年生苗木以牺牲根生长为代价向叶分配,4年生苗木的生物量继续倾向于叶分配,因此叶的异速生长指数随苗龄的增加呈递增趋势,说明叶受苗龄和个体大小的影响显著,具有较大的表型可塑性。各器官间除根与茎出现趋同进化外,叶与根、叶与茎和针叶长与叶生物量的异速生长轨迹发生改变,均具有表型可塑性。叶生物量和针叶长较为敏感,在苗龄为3 a 时有较大的叶生物量和针叶长,因而3年生云南松具有较高的生产能力。不同苗龄云南松地径、苗高、主根长、侧根长、针叶长在2年生苗木、3年生苗木有较高的变异度,可考虑在第1年后期对云南松进行密度、施肥和土壤微生物等因素调控,进而结合选育措施实现苗木生物量的改良,对更好地指导云南松高效培育技术的实施具有重要意义。
3.2 讨 论
3.2.1 器官与个体大小的异速生长关系
苗木的根、茎、叶都有特定的功能,根有锚地及养分和水分的吸收作用,茎有机械支撑和水力学疏导路径的作用,叶主要是有固定CO2并合成光合产物的作用,其分配受植株种类、大小、水分、年龄、温度及光照等因素的影响,以异速生长的形式表现出来,是维持其生长发育的一个生活史对策[20]。随着苗龄的增加,在云南松器官与个体大小的异速生长关系中,根生物量先下降后上升,呈等速生长关系趋势;茎生物量先下降后上升,呈等速生长关系趋势;叶生物量呈显著大于1.0 的异速生长关系趋势;地上生物量与根生物量先上升后下降,呈等速生长关系趋势。云南松随着苗龄的增加,其个体异速生长关系并不是恒定不变的,植株个体大小和苗龄同时影响着云南松的生长关系,随着个体发育异速生长关系有不同的变化趋势,具有物种特异性。有研究表明,地上与根生物量的大小能体现苗木对土壤养分或光照的需求。地上生物量与根在苗龄3 a 时有最大斜率,呈显著大于1.0 的异速生长关系,且异速生长轨迹发生改变,表明倾向于地上分配,这时地上生物量有较大的生产力。地上生物量与根是一个相互依赖、促进的有机整体。云南松在苗圃培育养分充足的条件下,地下部分能支撑地上部分,这时光成为了限制资源,苗木资源会倾向茎分配,帮助其茎干生长,增大叶面积,提高光捕获能力[21],导致了养分倾向于地上分配。云南松灵活的生长对策反映了其不同苗龄资源利用策略、能量物质投入的差异,是云南松对资源可利用水平与竞争胁迫生境的塑性适应[22]。
根在3 a、茎在2 a、叶在3 和4 a 与个体大小均为异速生长关系,且在这些苗龄中各器官的异速生长指数与其余苗龄异速生长指数差异显著,说明异速生长轨迹发生改变。个体大小显著影响着苗木生长速率,亦或生长速率存在显著的个体发育漂变。苗木生长速率的大小能体现苗木对资源的获取与利用能力,2年生云南松倾向于茎生长,3年生苗木以牺牲根生长为代价向叶分配,4年生苗木生物量继续倾向于叶分配,各器官与植株大小、地上与根异速生长关系随苗龄增加发生改变,使植株表现出一定的性状,满足自身发育的需要。其中叶的异速生长指数随苗龄增加呈递增趋势,说明叶受苗龄和个体大小的影响显著,具有较大的表型可塑性,即在有限的资源下,优先分配于叶生长,增加光合器官。有研究表明,分配于叶的资源物质随植物生活史阶段而变化,在幼苗时期的植物以增加光合器官的资源分配,来实现初始生长阶段的生物量积累;当植物个体逐渐增大时,减少叶生物量分配将有利于其在有限的资源条件下生存[23-24]。本研究中的云南松苗龄均较小,处于小个体时期,与这一理论相符。因此云南松的生长特性具有表型可塑性,而不同苗龄对云南松表型可塑性的影响不同[25]。这些生长特性增加了云南松的耐受性,使其能适应不同的环境和占据广阔的地理范围,成为广布性物种[26]。
3.2.2 器官与器官、形态指标间的异速生长关系
在植物个体生长发育的过程中,不同器官会相互协调发展[27],以最大化地利用资源来达到最适生长状态,致使不同器官相对生长速率出现差异。植物会在资源获取和物质消耗之间做出权衡,体现了生长与物质相协调的原则,是植物在一定环境中适应策略的重要表现[28]。云南松茎与根生物量有共同斜率,且截距、漂移差异不显著,说明茎与根在各苗龄中异速生长轨迹没有发生改变,呈比例生长,出现趋同进化趋势(即有相同的异速生长指数),异速生长关系较为稳定[7]。随着苗木的生长,保持表型发育稳定性在一定水平内,为确保发育进程的精准,因而打破这种平衡对有机体可能是致命的。有机体具备维持发育平衡、抵抗外界环境干扰的能力,也是确保其在不同环境中存活与生长的必要前提[29]。叶与根随苗龄的增加,从等速生长关系向显著大于1.0 的异速生长关系转变,即异速生长轨迹发生改变,表明3年生苗木、4年生苗木叶的生长速率要快于根,资源分配上向叶倾斜。这与陈国鹏等[30]对甘肃南部杜鹃的研究有差异,可能是由于云南松生长环境温暖,资源相对丰富,主要限制因子是光照有关。叶与茎的权衡不仅反映器官间的分配策略,本质上取决于植物生物力学特性和水力输送结构的需求。在2年生苗木、3年生苗木叶与茎生物量的异速生长关系和李亚男等[31]对杜鹃属植物茎与叶生物量关系的研究结果相似,随着苗龄的增加,叶与茎生物量斜率呈递增的趋势;在苗龄为3 a 时,呈显著大于1.0 的异速生长关系,叶显著大于茎的生物量增加速率,即较大的茎会有更高比例的叶生物量分配,增大叶面积,来提高同化速率和增加光合产物[32]。但苗龄在4 a时呈下降趋势,为与1.0差异不显著的等速生长关系。这可能是增大的叶需要更长的茎来减少内部对光能的遮挡,大叶片较小叶片需要更高比例的叶片生物量用于支持功能,茎就需要具有更高的支持能力[33]。
植物在各个生长阶段的发育重点不同,因此植株性状异速生长关系与生物量投资策略常随着发育而变化。叶片是植物进行光合作用及与外界环境进行物质和能量交换的主要场所,是对外界环境变化最敏感的器官,因而在众多生长特征中,叶性状最能够反映植物自身生长状态及适应环境的生存策略[34]。针叶长与叶生物量的异速生长关系和林华等[35]对苦竹叶片的性状研究相似,随着苗龄的增加针叶长呈先升高后降低的趋势,说明苗龄对针叶长性状影响显著。随着苗龄的增加总体会使针叶变长,且在苗龄为3 a 时有最大的生长速率。异速生长的变化可能与针叶生长投资出现“回报递减”现象有关。在发育早期叶细胞层较少,细胞排列相对松散,随着叶的生长,针叶长的增加有助于增加光合面积和光合同化结构,为了接触更多的光源和提高水分运输到叶肉细胞的效率,加大生物量投资用于微管组织和厚壁组织的建设,从而导致分配到叶肉细胞的生物量减少[36-37]。云南松可能凭借针叶长较大的表型可塑性,最大限度地进行光合积累获取资源,并进行资源的再分配,从而实现资源的有效利用。针叶长的表型可塑性提高了云南松在各苗龄中发育的合适度,对了解植物生活史对策和生态适应性具有重要意义。
生态代谢理论预测非木本植物或缺乏次生组织的小个体木本植物,根、茎、叶各器官间,各器官与总生物量间均为等速生长关系,本文中有一部分生长关系也出现了等速生长关系。其原因为对于小个体植物来说,除叶外茎也含有叶绿体,也可以固定CO2,不同组织和器官的代谢速率不同,光合速率可能随着资源分配和苗龄发生变化。而光合组织不只仅限于叶片,因此背离了等速生长关系[38]。亦或与遗传差异有关,云南松具有各自独特的生物学特性,随苗龄变化生长类型不唯一[7]。植物各器官之间因结构、功能的差异,代谢活性组织和异速指数随个体发育存在一个动态变化的过程,同时又受环境条件的制约。比如,当植物受到风的机械扰动,会导致根的生物量增加及茎的生物量减小;当植物的茎自然弯曲以及受到其他机械作用时,会对地上部分的生物量产生一定的影响;当受到干旱胁迫时,植物对根的生物量分配会有较大的增长;当温度较低时,植物会增加根的生物量分配,而减小茎和叶的生物量分配。反过来可以人为控制环境条件,以此来促进植物生长,以达到培育壮苗的目的。如改善热量条件,合理施水施肥,控制种群密度等。张霁等[39]用不同栽培方式,以光竞争的差异,使云南黄连异速生长的轨迹发生改变,相同大小的个体,栽培种比野生种具有更高的根茎生物量(植株药用部位)。本研究中云南松苗木生长性状在2、3 a 有较高的变异度,可考虑在第1年后期对云南松进行密度、施肥和土壤微生物等因素调控并研究适宜方案,并了解不同条件对云南松各个性状和根、茎、叶生物量分配及异速生长关系的影响;进而结合选育措施实现苗木生物量的改良,对更好地指导云南松高效培育技术的实施具有重要的意义。表型可塑性差异是由基因型和环境因素共同造成的,可通过基因组学、转录组学和代谢组学的综合研究,以达到高效培育的目的。但由环境诱导的表型可塑性变异能否被遗传等问题,需进一步研究。
猜你喜欢
作物学报(2022年2期)2022-11-06
中国交通信息化(2022年7期)2022-10-27
农业工程学报(2022年10期)2022-08-22
中国现代医生(2022年21期)2022-08-22
浙江农林大学学报(2022年2期)2022-04-08
农村科学实验(2022年2期)2022-03-12
广西植物(2021年9期)2021-10-16
安徽农学通报(2020年7期)2020-05-26
三农资讯半月报(2020年2期)2020-03-09
翠苑(2019年4期)2019-11-11
