
雨生红球藻磷脂-二酰甘油酰基转移酶基因的鉴定与表达分析
2022-03-17 12:37赵春超张宏江臧敦秀崔红利李润植
山西农业科学 2022年3期
赵春超,张宏江,臧敦秀,崔红利,李润植
(山西农业大学 农学院/分子农业与生物能源研究所,山西 太谷 030801)
来源于高等油料植物和富油微藻的植物油脂主要用于人类食用油和脂肪酸营养,亦是油脂化工和生物燃油等工业的优质原料。三酰甘油(Triacylglycerols,TAG)是真核生物储存能量的主要物质形式[1],以应对不利的环境条件[2],也在植物种子萌发过程中发生分解,为幼苗生长提供新能量。与高等油料植物相比,微藻具有光合作用强、生长周期短、繁殖速度快和油脂产量高等特点,并且被认为是蛋白质、肽、氨基酸、多糖、维生素、激素和脂肪酸等生物活性物质的优良生产者[3],市场开发应用潜力巨大。
雨生红球藻(Haematococcus pluvialis)是一种淡水单细胞绿藻,在胁迫条件下能合成和贮存大量类胡萝卜素-虾青素(Astaxanthin),被认为是获得天然虾青素最佳的生物种质。诱导雨生红球藻虾青素合成积累的胁迫条件包括高光[4]、氮缺乏[5]、高光与氮缺乏或磷缺乏结合[6]、营养胁迫[7]、盐胁迫[8]、高温[9]和化学调节剂[10]等。雨生红球藻细胞中的虾青素主要以酯的形式存在,包括虾青素单酯(70%)和虾青素双酯(25%),仅有5%的虾青素以游离(非酯化形式)的形式存在[11]。酯化后的虾青素与胞内合成的TAG一起贮藏于油滴内[12-14]。这表明虾青素的储存离不开TAG的大量合成。另有研究发现,在胁迫条件下,雨生红球藻细胞内的脂肪酸、总油脂和虾青素的积累紧密相关,并且抑制脂肪酸的合成会使虾青素的合成也受到抑制,但是抑制虾青素的合成却不会影响脂肪酸的合成[15]。因此,鉴定油脂合成途径的关键酶基因和功能将有助于解析雨生红球藻油脂合成和虾青素酯化的分子机制。
已有研究表明,陆地高等植物和微藻的TAG的生物合成途径是相似的[16]。TAG在内质网上的从头生物合成涉及甘油和酰基辅酶A,3种酰基转移酶依次将酰基辅酶A连接到甘油上,分别是甘油-3-磷酸酰基转移酶(Glycerol-3-phosphate Acvltransferase,GPAT)、溶血磷脂酸酰基转移酶(Lysophosphatidic Acid Acyltransferase,LPAAT)以及二酰甘油酰基转移酶(Acyl CoA:Diacylglycerol Acyltransferase,DGAT)[17]。然而,TAG的生物合成分为酰基辅酶A依赖途径(Kennedy途径)和非依赖途径[18]。在非依赖酰基辅酶A途径中,参与前2个反应的酶与Kennedy途径相同,唯一的区别在最后一步酰化反应,参与反应的酶不是DGAT,而是磷脂-二酰甘油酰基转移酶(Phospholipids:Diacylglycerol Acyltransferase,PDAT)。这最后一步酶促反应被认为是TAG合成的限速步骤。PDAT途径最早发现于酿酒酵母,且被认为是酵母对数生长时期TAG合成的主要途径[19]。许多高等植物[20]和莱茵衣藻[21]也发现PDAT途径。与广泛研究的DGAT酶相比,有关微藻PDAT途径的研究还鲜见报道。
为此,笔者以雨生红球藻-797株系为试材,鉴定和分子克隆编码磷脂-二酰甘油酰基转移酶(HpPDAT)的cDNA,应用组学工具检测Hp PDAT酶蛋白的理化特征,系统分析不同光质下HpPDAT的表达谱、雨生红球藻细胞的总脂合成情况,以及藻细胞生理生化参数,旨在为解析雨生红球藻油脂和虾青素的合成积累提供理论依据。
1 材料和方法
1.1 试验材料及培养条件
试验所用的雨生红球藻-797保存于山西农业大学分子农业与生物能源研究所。雨生红球藻的培养采用BG11培养基,于25μmol/(m2·s)的光照强度,12 h/12 h的光暗周期,(22±1)℃静置培养,每天摇匀2~3次。光诱导的条件设置为高白光、高蓝光、紫外和白光+紫外,高白光和高蓝光的光照强度采用10 000 lx,紫外的光照强度采用200 lx和400 lx,以黑暗条件作为对照,除光照外其他条件与正常培养相同。
1.2 雨生红球藻PDAT基因的筛选
从NCBI数据库中下载已知的莱茵衣藻PDAT基因(JN 815265.1)为探针,通过Blast搜索到普氏小球藻、栅藻、酿酒酵母、大豆、拟南芥等20个物种的PDAT同源基因。以20条不同物种的PDAT基因序列为模板,以雨生红球藻的转录组数据库为检索基础,通过本地Blast在其中检索雨生红球藻的PDAT基因序列,并对检索结果进行鉴定。
1.3 雨生红球藻PDAT蛋白的生物信息学分析
利用在线序列处理工具包(the sequence manipulation suite,SMS)将获得的HpPDAT的cDNA全长序列翻译成氨基酸序列,并通过ORF finder软件查找开放阅读框;使用在线工具ProtParam进行理化性质预测;使用在线工具TMHMM Server v.2.0进行跨膜结构域预测;使用在线工具PSORTⅡ进行亚细胞定位预测;使用在线工具SignalP-5.0进行信号肽预测;使用在线工具NetPhos-3.1进行磷酸化位点预测;Hp PDAT蛋白的二级结构和三级结构的预测利用在线工具SOPMA和SWISSMDEL进行;利用保守结构域数据库CDD(Conserved Domain Database)鉴定HpPDAT蛋白的保守域。使用工具的网址如表1所示。

表1 生物信息学工具Tab.1 Tools of bioinformatics
1.4 雨生红球藻PDAT蛋白的多序列比对和系统发育分析
选取雨生红球藻、莱茵衣藻、普氏小球藻、缺刻叶球藻、拟南芥、大豆的PDAT蛋白序列,使用软件ClustalX进行多序列比对。以表2中的PDAT序列,运用MEGA 7软件采用邻位连接法(NJ)对PDAT蛋白构建系统进化树,以泊松校正法计算进化距离,设定Bootstrap值为1 000[22]。

表2 不同物种PDAT蛋白的基本信息Tab.2 Basic information on PDAT proteins of differ ent species
1.5 雨生红球藻Hp PDAT的表达检测
以本实验室的转录组数据为基础,调取其中不同光处理条件下PDAT基因的FPKM值,计算表达量和相对表达量(黑暗条件为1),并以相对表达量绘图。同时qPCR检测Hp PDAT基因在不同光质条件下的表达谱。qRT-PCR引物采用Primer 6.0设计,内参基因Actin引物和定量引物如表3所示。RNA的提取使用RNA提取试剂盒(TaKaRa)。获得的RNA使用反转录试剂盒(TaKaRa)合成cDNA。使用2×实时荧光定量PCR Mix(TaKaRa)进行qRT-PCR。以Actin作为内参进行基因表达分析。每个样品至少进行3次生物学重复。
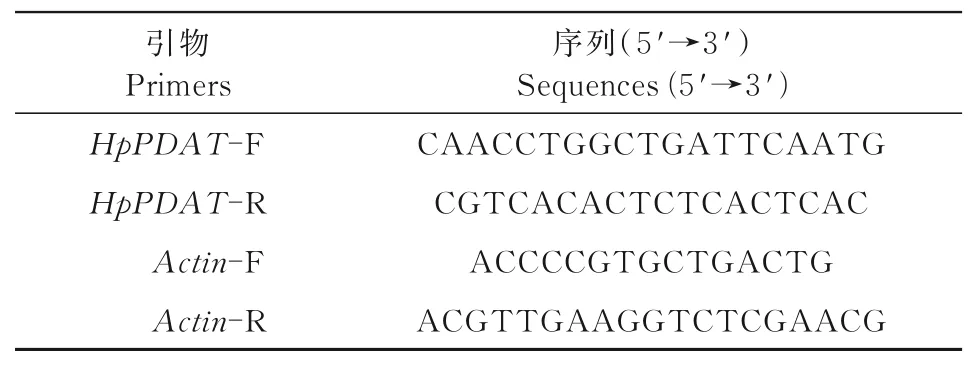
表3 PCR引物Tab.3 Primers of PCR
1.6 雨生红球藻总脂提取
将培养的藻种离心收集并冻干,研钵研磨备用。称取50 mg的藻粉,质量记为W1,倒入50 mL的离心管中,并加入7.5 mL甲醇∶氯仿(体积比2∶1),漩涡震荡5 min,于37℃摇床提取24 h后,5 000 r/min离心5 min,收集上清至新的50 mL离心管。于沉淀管中再次加入7.5 mL甲醇∶氯仿(体积比2∶1),漩涡振荡5 min,于37℃摇床提取2 h后,5 000 r/min离心5 min收集上清,与上一步的上清混合。可重复几次。向上清管中加入5 mL氯仿和9 mL 1%NaCl,保证终体系中甲醇∶氯仿∶水的体积比为2∶2∶1.8,漩涡振荡5 min后,8 000 rpm离心10 min取下清,将下清液移入干净玻璃管内(已烘干至恒质量,质量记为W2)。利用氮吹仪加速氯仿挥发,待氯仿挥发殆尽后,于105℃烘箱内烘干3 h。待玻璃管冷却后称质量,质量记为W3。
2 结果与分析
2.1 雨生红球藻Hp PDAT基因的鉴定
通过本地Blast,在雨生红球藻转录组数据库中筛选出1个具有完整开放阅读框(ORF)的候选HpPDAT基因,全长3 138 bp,5′-UTR 75 bp,3′-UTR 129 bp,ORF 2 934,编码978个氨基酸(图1)。将获得的序列利用Pfam Search(http://pfam.xfam.org/search)进行基因家族鉴定,预测蛋白包含有典型的卵磷脂胆固醇酰基转移酶功能结构域(PF02450),表明该cDNA编码的是一种PDAT酶蛋白。
2.2 雨生红球藻Hp PDAT蛋白基本理化性质分析
利用工具ProtParam对雨生红球藻Hp PDAT蛋白进行分析,结果显示,蛋白的分子式为C4437H6997O1396N1289S38,分子质量为101.95 ku,理论等电点6.15,属于酸性蛋白。HpPDAT蛋白共有978个氨基酸,其中,丙氨酸(Ala)含量最高(13.4%);其次为丝氨酸(Ser)和甘氨酸(Gly),含量分别为10.6%和10.5%;而半胱氨酸(Cys)含量最少(0.9%)。碱性氨基酸残基总数(Asp+Glu)共92个,酸性氨基酸残基总数(Arg+Lys)共80个。该蛋白的不稳定性指数(Ⅱ)为39.68,脂肪指数为74.82,属于稳定蛋白。总平均亲水系数为-0.238,为亲水性蛋白。

在线分析软件TMHMM结果表明,HpPDAT蛋白包含一个跨膜区域,是一种膜结合蛋白。通过PSORTⅡ软件对HpPDAT蛋白的亚细胞定位进行预测,结果显示,HpPDAT蛋白定位于内质网(55.6%),与TMHMM的预测相吻合。SignalP-5.0的预测结果显示,雨生红球藻Hp PDAT蛋白的序列中不包含信号肽序列。NetPhos-3.1的预测结果显示,Hp PDAT蛋白的丝氨酸激酶(Ser)的位点最多,有81个;苏氨酸激酶(Thr)的位点有29个;酪氨酸激酶(Tyr)的位点最少,仅为4个。
2.3 雨生红球藻Hp PDAT蛋白的二级结构和三级结构
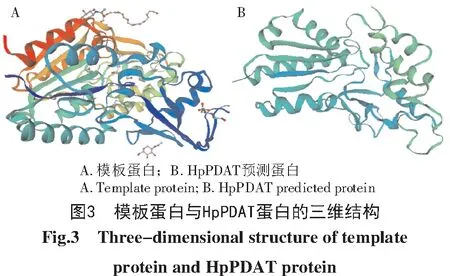
利用在线分析软件SOPMA对Hp PDAT蛋白的二级结构进行预测,预测结果如图2所示。α-螺旋所占的比例最高,为40.33%,接下来的无规则卷曲比列为37.56%,最后延伸链和β-转角的比列分别为11.77%和10.34%。表明主要由α-螺旋和无规则卷曲构成雨生红球藻HpPDAT蛋白的二级结构,该蛋白为混合型蛋白。SWISS-MODEL的三级结构预测结果如图3所示,HpPDAT蛋白与模板蛋白相似性较高,但存在一些结构上的差异,这些差异可能与HpPDAT的功能相适应。
2.4 雨生红球藻Hp PDAT蛋白的序列比对和系统进化分析
以3种藻、2种植物与雨生红球藻的PDAT共同进行序列比对,比对结果如图4所示,此比对结果进行了序列的编辑。结果显示,在非保守元件部分,各个序列之间的相似性较低,但在LCAT家族的保守元件部分,它们间的相似性保持了较高的水平。黑色方框内即为LCAT家族的保守元件,依次分别为Lid区、盐桥以及催化三联体(包含3个区域),并且其内包含的活性位点保持了高度的一致性,其中Lid区的为Trp,盐桥的为Asp和Arg,催化三联体的为Ser、His、Asp。


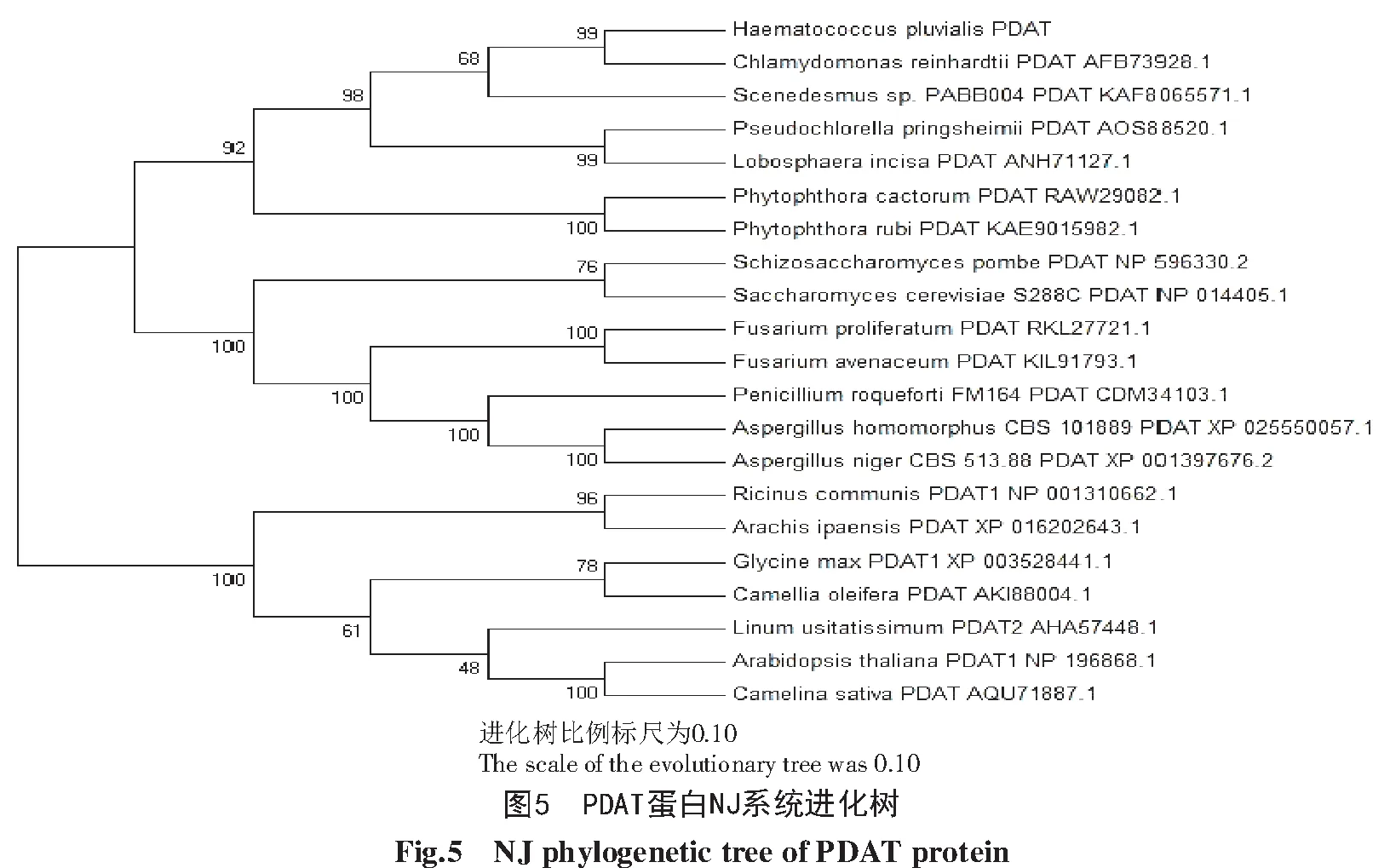
系统进化结果如图5所示,所有物种被分为了3个来源。所有的植物被分为一组,表明它们的PDAT具有相同的来源;除了疫霉属的真菌外,其他的菌类被分为一组,并且它们之间的同源性较高;藻类和疫霉属的真菌被分为一组,除了栅藻外,各个藻类之间的同源性较高。
2.5 雨生红球藻Hp PDAT的表达谱和细胞总脂积累分析
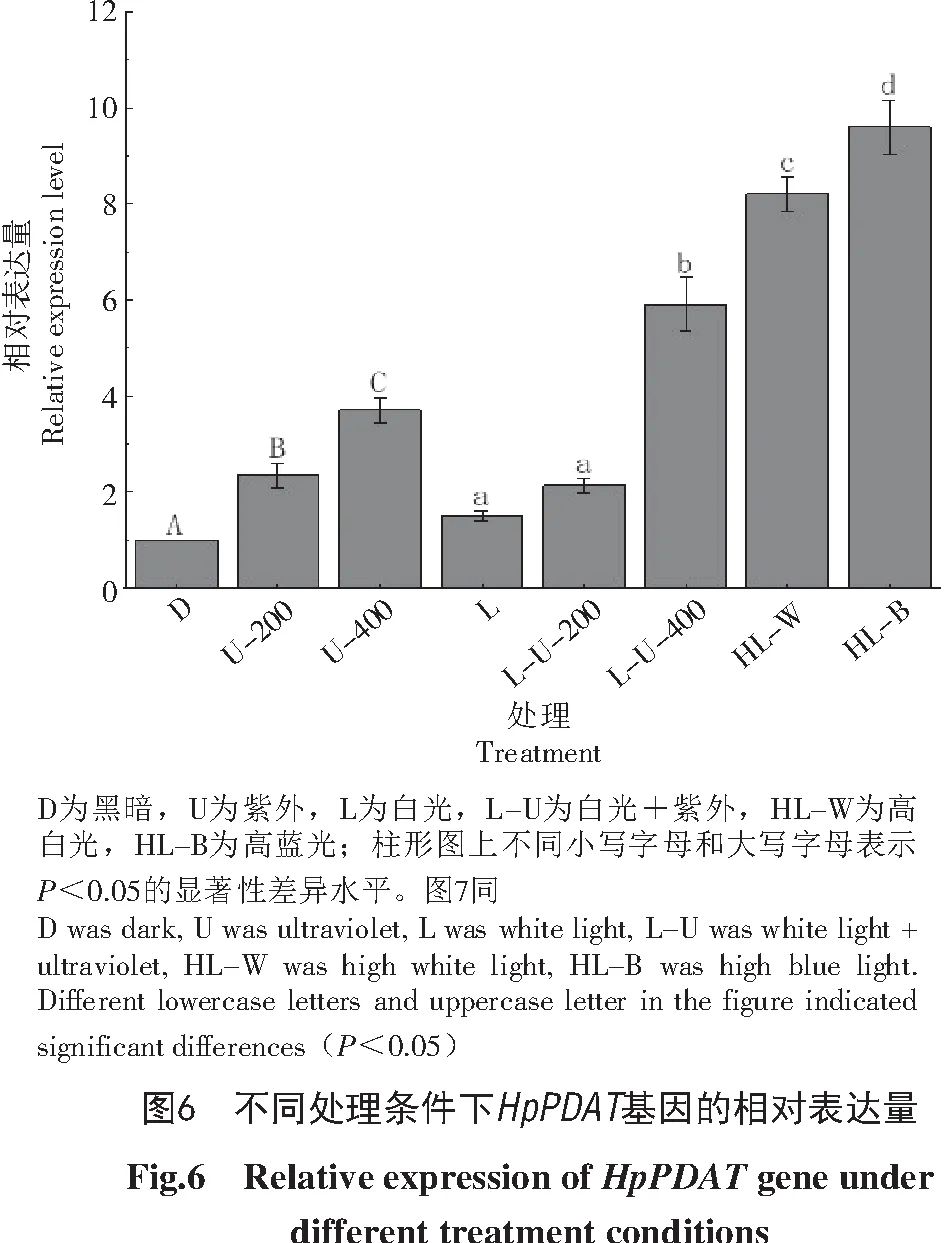
本试验选取了黑暗、白光、蓝光和紫外光作为不同的处理条件,HpPDAT的相对表达量如图6所示。总体来看,光照条件下HpPDAT的表达水平要高于黑暗条件下。无论是在黑暗还是光照条件下,紫外光的加入都会使藻细胞上调HpPDAT的表达,在黑暗条件下HpPDAT表达的上调并不明显,但在光照条件下,400 lx的紫外线使得HpPDAT的表达量显著提高,可以达到黑暗条件下的5.9倍,正常光照条件下的3.9倍。高白光(10 000 lx)诱导使得HpPDAT的表达量达到黑暗处理的8.2倍,而在高蓝光(10 000 lx)处理下则达到9.6倍;相比于正常光照条件,高白光下HpPDAT的表达量是其5.5倍,高蓝光下HpPDAT的表达量是其6.4倍。
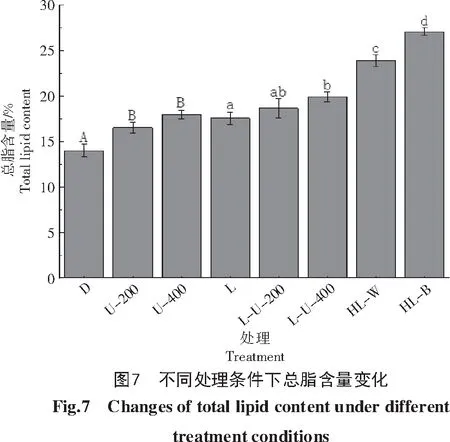
利用收集的藻粉提取并测定总脂含量,结果如图7所示。黑暗条件下细胞总脂的含量要明显低于光照条件。与对照相比,紫外光(200、400 lx)的加入会使得总脂含量增加。高光对于总脂积累的促进作用要明显好于紫外光,高白光(10 000 lx)下总脂含量达到24%,是黑暗处理的1.7倍,而在高蓝光(10 000 lx)处理下获得了最高的总脂含量,达到了27%,是黑暗处理的1.9倍。PDAT的表达量与总脂的积累量呈现出一致性,但在L-U-200处理下出现差异。此处理下细胞PDAT的表达量相较于U-200处理有所降低,但总脂含量比U-200处理的要高。推测是细胞中TAG合成的另一限速酶DGAT的参与使得其出现了差异,有待后续分析。
3 结论与讨论
雨生红球藻是一种高附加值的微藻,其细胞合成的虾青素具有显著的生物学活性[23],并且藻源TAG是生物燃油的优良原料。TAG的合成对于雨生红球藻具有重要意义,一方面其作为一种能源物质应对逆境胁迫,另一方面关系到虾青素的储存。胁迫条件下雨生红球藻会大量积累TAG和虾青素[24],两者之间的关联机制还未阐明,同时虾青素的酯化机制也不明确。雨生红球藻DGAT作为TAG合成途径中的限速酶之一已有大量的相关研究[25-27],但另一限速酶PDAT却无相关报道。本试验选取HpPDAT作为研究对象,以期为探究高光逆境条件下雨生红球藻油脂和虾青素的合成积累提供新思路。
以莱茵衣藻Cr PDAT序列为探针,分析雨生红球藻转录组的本地数据库,筛选出1个HpPDAT基因,序列全长3 138 bp,编码978个氨基酸。生物信息学分析结果显示,HpPDAT属于LCAT家族,并且具有PDAT蛋白的典型功能结构域。对Hp PDAT蛋白的进化分析结果显示,其在进化上具有较高的保守性,与微藻的亲缘关系较近,而与植物的亲缘关系较远。预测Hp PDAT是一种膜结合蛋白,并被定位于内质网,这与TAG于内质网被合成的事实相符[28-29],表明Hp PDAT蛋白被合成、修饰后锚定在内质网发挥功能。
雨生红球藻光诱导试验结果表明,与黑暗条件相比,不同光质对于HpPDAT的表达有不同程度的促进作用,施加一定强度(200、400 lx)的紫外光可以促进其表达;高光(10 000 lx)具有显著的促进作用,特别是高蓝光(10 000 lx)下,Hp PDAT的表达量可以达到黑暗条件的9.6倍,可能是雨生红球藻细胞对于蓝光具有偏好性。过量的紫外线可以造成细胞的损伤,甚至导致死亡[30],一定强度(200、400 lx)紫外光下HpPDAT的上调表达可能是雨生红球藻细胞应对紫外线损害的途径之一。同时,总脂积累分析显示,光照条件下总脂的积累情况要好于黑暗条件,并且在高蓝光(10 000 lx)处理下获得了最好的总脂积累(27%),达到黑暗处理的1.9倍。再次表明雨生红球藻细胞对于蓝光可能具有偏好性。PDAT的表达与总脂的积累呈现出一致性,但有一定的差异,出现差异可能是因为TAG的合成还涉及到DGAT的表达,因此,光照对于总脂积累的影响有待进一步研究。
本试验获得了一条雨生红球藻Hp PDAT的cDNA序列,发现其属于LCAT家族,具有PDAT蛋白的功能。系统发育结果显示,Hp PDAT与微藻的亲缘关系较近,而与植物的亲缘关系较远。光诱导试验表明,高蓝光(10 000 lx)对于HpPDAT的表达具有显著的促进作用,同时使细胞获得了最高的总脂积累。本研究为探究高光逆境条件下雨生红球藻油脂富集和虾青素酯化的分子机制提供了可靠的理论依据。
猜你喜欢
保健医苑(2022年6期)2022-07-08
中南民族大学学报(自然科学版)(2022年4期)2022-07-01
新高考·高二数学(2022年3期)2022-04-29
家庭影院技术(2021年10期)2021-11-20
落叶果树(2021年6期)2021-02-12
小学阅读指南·低年级版(2016年10期)2016-09-10
求学·理科版(2015年10期)2015-11-04
江淮论坛(2011年2期)2011-03-20
现代计算机(2009年3期)2009-12-21
中国化妆品(2009年1期)2009-02-13
