
琥珀酸脱氢酶B亚基对卵巢癌细胞线粒体DNA拷贝数影响的研究*
2022-03-23 02:48陈立兰
现代妇产科进展 2022年3期
陈立兰,狄 文,2,3
(1.上海交通大学医学院附属仁济医院妇产科,上海 200127;2.上海市妇科肿瘤重点实验室,上海 200127;3.癌基因及相关基因国家重点实验室,上海 200127)
卵巢癌是死亡率最高的妇科恶性肿瘤[1],代谢异常在肿瘤的发生、发展过程中发挥重要作用[2]。琥珀酸脱氢酶(succinate dehydrogenase,SDH)是参与三羧酸循环-氧化磷酸化的重要酶,在肿瘤代谢中发挥重要作用[3]。研究发现,SDH在多种肿瘤中异常低表达[4]或发生基因突变[5-6],因此认为SDH及其变异可能是影响肿瘤进展的关键分子。SDHB为SDH亚基之一,SDHB在结肠癌耐药细胞和非霍奇金淋巴瘤(non-Hodgkin's lymphoma,NHL)Ramos耐药细胞中低表达[7-8]。在肠相关神经内分泌肿瘤中,SDHB低表达与肿瘤转移患者的预后相关[9],均提示SDHB在恶性肿瘤中发挥重要作用。人线粒体DNA(mitochondrial DNA,mtDNA)是独立于核DNA之外的双链闭合环状分子,线粒体能量代谢的改变与mtDNA拷贝数密切相关[10]。研究发现,线粒体DNA拷贝数与神经胶质瘤患者的预后呈正相关[11]。线粒体DNA拷贝数增加可通过增强结肠癌微卫星病灶线粒体氧化磷酸化促进肿瘤进展[12],提示线粒体DNA拷贝数在肿瘤中发挥重要作用。本课题组前期研究证实,SDHB在人卵巢癌组织中低表达,并且SDHB低表达显著促进卵巢癌细胞的增殖、侵袭和迁移能力[13]。但是,目前卵巢癌中SDHB与mtDNA拷贝数的关系尚不清楚。本文拟探讨人卵巢癌细胞中SDHB对mtDNA相对拷贝数的影响及机制,为卵巢癌的诊疗提供新思路。
1 材料与方法
1.1 主要材料与试剂 人卵巢癌腺癌细胞株A2780、SKOV3(中科院上海细胞所)。选取2011年7月至2012年6月就诊于上海交通大学医学院附属仁济医院的6例良性卵巢肿瘤患者和14例上皮性卵巢癌患者的病理组织标本,患者均签署知情同意书。上海艾博思生物科技公司设计合成人SDHB siRNA序列(siSucA 5'-GATTAAGAATGAAGTTGACTC-3',siSucC 5'-GCTCAGAGCTGAACATAATT-3'),阴性对照(Negative Control)(NC:UUCUCCGAACGUGUCACGUTT)。SDHB和内参β-actin引物序列由上海生工合成;转染试剂Lipofectmin 2000、Trizol(Invitrogen);RPMI 1640细胞培养基、胎牛血清FBS(Hyclone);SDHB多克隆抗体、β-tubulin单克隆抗体(Epitomics),β-actin抗体(Abmart),p38抗体、p-P38抗体、γ-H2AX抗体(Epitomics),羊抗兔(鼠)远红外荧光二抗(LI-COR生物科技);SDS-PAGE胶试剂盒(碧云天生物);SYBR实时荧光定量PCR试剂盒及反转录试剂盒(TAKARA);ABI Step one plus实时荧光定量仪;ABI反转录仪;远红外荧光扫描仪(Odyssey);基因组DNA提取试剂盒(碧云天),CCK-8检测试剂盒(碧云天),顺铂(山东齐鲁制药)。
1.2 实验方法
1.2.1 Trizol法提取卵巢组织和细胞中RNA 从-80℃冰箱中取出组织,取豆粒大组织于1mL Trizol中,利用液氮研磨。将混合物置1.5mL离心管,室温静置10min,加氯仿200μL/管(国产分析纯),混匀。室温静置10min,4℃ 12000r/min,离心10min。水相转移至新的EP管,分别加和水相等体积的异丙醇混匀。室温沉淀10min,4℃ 12000r/min,离心10min,弃上清。沉淀用75%乙醇洗涤2次,4℃ 7500r/min,离心5min,弃乙醇,倒置于滤纸晾干,加DEPC水溶解RNA沉淀。进行RNA浓度测定,调整RNA浓度500ng/μL进行反转录。反转录程序为37℃ 15min,85℃ 5s。同法提取卵巢癌细胞株RNA。
1.2.2 Real-time PCR检测SDHB和线粒体转录因子A(mitochondrial transcription factor A,TFAM)水平 取上一步骤反转录cDNA。引物序列:SDHB上游5'-AAATGTGGCCCCATGGTATTG-3',下游5'-AGAGCCACAGATGCCTTCTCTG-3');β-actin上游5'-TGACGTGGACATCCGCAAAG-3',下游5'-CTGGAAGGTGGACAGCGAGG-3');TFAM上游5'-CCAAAAAGACCTCGTTCAGCTTA-3',下游5'-CTTTACAGTCTTCAGCTTTTCCTGC-3')。Real-time PCR反应条件:95℃预变性30s,95℃ 5s,60℃ 30s,40个循环,融解曲线分析程序为95℃ 0s,65℃ 15s,95℃ 0s。采用2-△CT法相对定量比较SDHB和TFAM mRNA水平。
1.2.3 细胞siRNA干扰 卵巢癌细胞株SKOV3、A2780用含10%FBS的RPMI 1640培养基培养,置37℃、5%CO2恒温培养箱孵育。取对数生长期细胞,将细胞分为转染试剂组、阴性对照组和实验组。收集对数生长期SKOV3、A2780细胞,将2×105细胞重悬于2mL含10% FBS RPMI 1640中,接种于6孔板。将细胞置37℃、5% CO2恒温培养箱孵育24h,细胞密度达40%~50%时进行转染。转染后置恒温培养箱孵育,4h后换成含10%FBS的RPMI 1640培养液继续培养24~48h。
1.2.4 Real-time PCR和Western blot法检测SDHB干扰效率 收集转染24h的SKOV3、A2780细胞,Trizol法提取RNA,并进行逆转录。实时定量PCR法检测SDHB mRNA水平,方法同前。收集转染48h的SKOV3、A2780细胞,用RIPA裂解液(加蛋白酶抑制剂PMSF 100×)于冰上裂解细胞30min,4℃ 12000r/min,离心10min,取上清,加上样缓冲液(5×,碧云天),100℃煮沸10min变性。10%SDS-PAGE胶分离蛋白,将蛋白转移至硝酸纤维素膜(NC),5%脱脂奶粉室温封闭2h,内参β-tubulin(1∶5000)或β-actin(1∶500)和目的蛋白SDHB(1∶1000)室温摇床孵育2h,TBST洗涤3次,每次5min,远红外荧光二抗分别避光室温摇床孵育1h,TBST洗涤3次,每次5min,Odyssey扫膜仪扫描显影。
1.2.5 Real-time PCR检测干扰前后线粒体相对拷贝数(mtND2/HBB)、TFAM和PRKDC变化 干扰SDHB 24h后,收集细胞,提取基因组DNA和RNA,依据基因组DNA小量抽提试剂盒(离心柱式,碧云天)方法提取基因组DNA,RNA提取方法同前。引物序列:ND2上游5'-CACAGAAGCTGCCATCAAGTA-3',下游5'-CCGGAGAGTATATTGTTGAAGAG-3');HBB(β-globin)上游5'-CAGGTACGGCTGTCATCACTTAGA-3',下游5'-CATGGTGTCTGTTTGAGGTTGCTA-3';PRKDC上游5'-CTGTGCAACTTCACTAAGTCCA-3',下游5'-CAATCTGAGGACGAATTGCCT-3'。实时荧光定量检测程序同前。HBB是核单拷贝基因,ND2是线粒体编码基因,通过mtND2/HBB比值比较细胞线粒体相对DNA拷贝数,TFAM与线粒体DNA拷贝数相关,PRKDC为DNA依赖蛋白激酶催化亚基,影响DNA损伤修复能力。
1.2.6 CCK-8检测卵巢癌对顺铂的化疗耐药性 选择对数生长期卵巢癌细胞,胰酶消化细胞,全陪重悬细胞。细胞计数,调整细胞浓度5×104细胞/mL;96孔板每孔100μL细胞悬液,细胞培养箱过夜。根据脂质体2000转染试剂说明书,siRNA干扰SDHB表达,6h换液。48h后,将不同浓度梯度顺铂加入处理后的细胞中,细胞培养箱培养24h。CCK-8试剂盒检测细胞活性。弃96孔板中培养液,按CCK-8∶RPMI 1640=1∶9的比例配置检测液,每孔加100μL,1.5~2h检测450nm波长处OD值。计算24h半数抑制浓度IC50。
1.2.7 Western blot法检测干扰SDHB前后p38磷酸化水平和DNA损伤标记蛋白γ-H2AX水平 siRNA干扰48h,收集细胞提取蛋白,Western blot法检测干扰SDHB后p-p38和γ-H2AX蛋白变化。
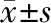
2 结 果
2.1 SDHB在人卵巢癌组织中的表达 人卵巢癌组织中SDHB mRNA水平低于卵巢良性肿瘤,后者约是前者的4.34倍(P<0.05)(图1A)。
2.2 mtND2/HBB和TFAM在人卵巢癌组织中的表达 人卵巢癌组织中mtND2/HBB和TFAM mRNA表达量均显著高于卵巢良性肿瘤组织(P<0.05),前者中含量分别是后者的3.98倍和1.80倍(图1B、C)。

图1 SDHB、mtND2/HBB和TFAM在人卵巢癌组织中的表达
2.3 SKOV3和A2780细胞株中SDHB干扰效率检测 SKOV3和A2780细胞株转染SDHB siRNA 24h,显著抑制mRNA水平:89.80%(siSucA vs NC,SKOV3),84.89%(siSucC vs NC,SKOV3),83.88%(siSucA vs NC,A2780),差异有统计学意义(P<0.001);转染48h时蛋白表达水平显著降低(图2A、B)。
图2 siRNA干扰序列显著抑制卵巢癌细胞中SDHB表达
2.4 Real-time PCR法检测干扰SDHB后卵巢癌细胞株mtDNA拷贝数和TFAM变化以及顺铂耐药性 siRNA干扰SDHB,SKOV3和A2780中mtND2/HBB和PRKDC水平显著增加(P<0.001,P<0.05),TFAM也增加,但差异无统计学意义(P>0.05)。SDHB低表达显著提高癌细胞的顺铂耐药性(图3A、B、C)。

图3 降低SDHB表达后卵巢癌细胞株mtDNA拷贝数和TFAM变化及顺铂耐药性变化
2.5 SDHB低表达对卵巢癌细胞p-p38和DNA损伤标记蛋白γ-H2AX蛋白水平的影响 SiRNA干扰卵巢癌细胞中SDHB 48h,Western blot结果显示降低SDHB时,SKOV3和A2780中p-p38蛋白水平显著增加,而总蛋白水平变化不明显,γ-H2AX蛋白水平显著降低(图4、5)。

图4 SDHB低表达对卵巢癌SKOV3细胞PRKDC mRNA水平和DNA损伤标记蛋白γ-H2AX蛋白水平的影响

图5 Western blot法检测干扰SDHB后SKOV3和A2780细胞中p-p38蛋白水平
3 讨 论
三羧酸循环在能量代谢中发挥重要作用,琥珀酸脱氢酶作为三羧酸循环的重要酶之一,在肿瘤研究中备受关注。线粒体是细胞进行三羧酸循环的主要场所,是高等动物细胞核外唯一含有DNA的细胞器。人类mtDNA基因编码13个与线粒体氧化磷酸化有关的蛋白质[14],mtDNA的功能实现不但与其结构完整性有关,还与其拷贝数有关。研究发现,mtDNA拷贝数显著促进结肠癌细胞的增殖和转移[12],mtDNA拷贝数与胰腺癌的发生呈负相关[15],表明mtDNA拷贝数在不同肿瘤中的作用及机制不同。
TFAM是影响线粒体DNA转录和复制的重要调节子,TFAM缺失会导致线粒体功能紊乱,与许多肿瘤的发生发展密切相关[16]。TFAM调节线粒体启动子的起始,因此TFAM可影响基因的表达和mtDNA的转录[17],但是由于轻链启动子处的顿挫型转录体可启动DNA的复制,TFAM对于mtDNA的复制处于次要的地位。本研究显示,人卵巢癌组织中mtND2/HBB和TFAM表达水平显著高于卵巢良性肿瘤,SDHB低表达时卵巢癌细胞株中线粒体DNA相对拷贝数和TFAM均上调,表明SDHB影响卵巢癌的线粒体DNA拷贝数,但机制不明确。
DNA依赖性蛋白激酶(protein kinase,DNA-activated,DNA-PK)是基因组DNA损伤修复过程中的关键蛋白激酶,由DNA依赖蛋白激酶催化亚基(DNA-PK catalytic subunit,DNA-PKcs,也称作PRKDC)和Ku70/80蛋白异质二聚体组成的丝/苏氨酸蛋白激酶构成[18]。PRKDC与DNA结合后被激活,当双链断裂(DSB)发生时,PRKDC被Ku70/80异源二聚体招募到DSBs,在多个丝氨酸和苏氨酸残基磷酸化后迅速被激活从而影响DNA损伤修复能力[19]。本研究发现,抑制卵巢癌细胞SDHB表达可显著降低DNA损伤标记蛋白γ-H2AX表达,增加PRKDC mRNA水平,表明SDHB低表达显著增加卵巢癌细胞的DNA损伤修复能力,而卵巢癌的化疗耐药性与DNA损伤修复密切相关[20]。推测SDHB通过影响DNA损伤修复影响线粒体DNA拷贝数,进一步影响卵巢癌的进展。本研究显示,SDHB低表达显著增加卵巢癌细胞的顺铂耐药性,表明SDHB可能通过影响线粒体DNA影响化疗效果。DNA丝裂原活化蛋白激酶(MAPK)信号通路是广泛存在于多种细胞中的信号转导途径,能将细胞外信号转导至细胞及细胞核内,通过保守的三级级联反应激活转录因子,调节基因表达,参与细胞运动、凋亡、分化及生长增殖等多种生理过程[21]。Wnt诱导的小鼠间充质干细胞C3H10T1/2向成骨细胞分化时通过ERK和p38 MAPK信号通路增加PGC-1α表达从而增加了mtDNA的拷贝数[22]。PRKDC通过p38 MAPK通路可促进乳腺癌细胞的生长[23],表明p38 MAPK通路在稳定线粒体DNA拷贝数中有一定作用。本研究结果显示,SDHB低表达显著上调p38磷酸化蛋白水平,推测SDHB可能通过p38 MAPK通路影响PRKDC影响DNA损伤修复进一步影响线粒体DNA拷贝数,影响卵巢癌细胞的化疗耐药性。
综上所述,SDHB在卵巢癌中低表达且影响线粒体DNA拷贝数,为卵巢癌进一步研究提供方向。在以后研究中将进一步明确SDHB通过影响线粒体DNA拷贝数和DNA损伤修复能力影响卵巢癌化疗耐药的具体机制。
猜你喜欢
中国癌症防治杂志(2022年4期)2022-11-24
检验医学与临床(2022年19期)2022-10-10
右江民族医学院学报(2022年2期)2022-05-19
昆明医科大学学报(2022年3期)2022-04-19
中国现代药物应用(2022年5期)2022-04-14
昆明医科大学学报(2022年1期)2022-02-28
国际放射医学核医学杂志(2021年10期)2021-02-28
江苏农业科学(2019年14期)2019-09-23
分析化学(2015年3期)2015-04-20
中国民族民间医药·下半月(2014年4期)2014-09-26
