
鹅掌楸属种间杂交子代生长动态分析
2022-05-13 02:14蔡桁徐红梅蒋祥娥曹健董梅胡超杨寒管兰华
湖北林业科技 2022年2期
蔡桁 徐红梅 蒋祥娥 曹健 董梅 胡超 杨寒 管兰华
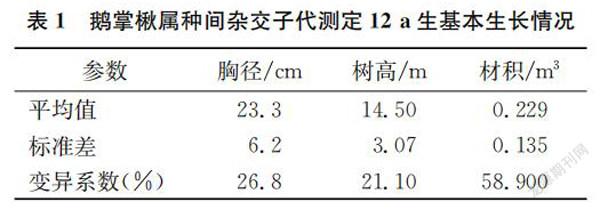


摘要:以12 a生鹅掌楸属种间杂交子代测定林为对象,利用连年生长数据进行了生长差异性分析、相关分析和生长动态分析。结果表明:12 a子代测定林平均胸径23.3 cm,平均树高14.50 m,平均材积0.229 m3,杂交组合之间连年生长差异极显著。造林后第6年与第11年胸径、树高和材积的相关系数分别为0.903、0.911、0.914,相关密切,早期选择具有较高的可靠性。杂交子代测定林的速生期为第2年~第8年,胸径、树高生长数量成熟在第8a和9a,此时材积生长仍然保持较高水平,其生长数量成熟要晚于第11年。
关键词:鹅掌楸;子代测定;生长动态分析;数量成熟
中图分类号:S792.21文献标识码:A文章编号:1004-3020(2022)02-0013-06
Dynamic Growth Analysis of Interspecific Hybrid Progenies of Liriodendron chinense
Cai Heng Xu Hongmei Jiang Xiang Cao Jian Dong Mei Hu Chao Yang Han Guan Lanhua
Abstract:In this paper,the growth difference analysis,correlation analysis and growth dynamic analysis were carried out based on the growth data of 12yearold interspecific hybrid progeny test of Liriodendron chinense. The results showed that the average DBH,tree height and volume of trees were 23.3 cm,14.50 m and 0.229 m3,respectively. The correlation coefficients of DBH,tree height and volume were 0.903,0.911 and 0.914 respectively between 6th year and 8th year after afforestation,indicating that early selection had high reliability. The rapid growth period of the hybrid progeny test was at 2~8 year. Quantitative maturity of DBH and tree height were at 8th and 9th year,at this time,the volume growth still maintained a high level,and the growth matured quantitatively later than 11th years.
Key words:Liriodendron chinense;progeny test;growth dynamic analysis;quantitative maturity
鵝掌楸属Liriodendron现仅存两个种[1],鹅掌楸L. chinense和北美鹅掌楸L. tulipifera ,亚美马褂木Liriodendron×sinoamericanum 为鹅掌楸与北美鹅掌楸的人工杂种的统称[2]。亚美马褂木具有很强的杂种优势,适应性强,生长迅速,树形美观,作为珍贵用材及园林绿化等树种具有十分广阔的推广应用前景。20世纪60年代以来,在鹅掌楸属种间杂交及杂种优势分析[36]、无性繁殖[711]、引种与栽培技术[1219]、胁迫反应[2023]、木材特性[2426]、基因克隆[2730]等方面开展了大量研究。但以往选育的亚美马褂木存在亲本来源少,杂种子代遗传基础狭窄等缺陷[31],同时鹅掌楸属树种的生长特性等方面的研究较少[3234],为了揭示鹅掌楸属种间杂种亚美马褂木生长性状的遗传变异、生长动态等特性,本研究利用鹅掌楸属杂交子代测定林进行了亚美马褂木生长分析,期望了解亚美马褂木生长特性,为人工林培育提供技术支撑;通过早期选择研究,评价对成年目的性状进行早期预测和选择的可行性,从而达到缩短育种周期的目的,以便及时将优良材料用作生产群体的亲本或作高世代育种材料[35]。
1材料与方法
1.1试验地概况
试验地位于湖北省咸宁市咸安区林业科学研究所(114°23′E,29°59′N),亚热带季风性湿润气候,四季分明,气候温和。年均气温16.8 ℃,最冷月1月平均气温4 ℃,最热月7月平均气温29.3 ℃。降雨量年均1 531.4 mm,无霜期年均258 d。
1.2试验材料
鹅掌楸属种间杂交于2007年在湖北省京山市虎爪山林场鹅掌楸属种源试验林内进行[6], 2008年在湖北省林木育种中心苗圃分家系育苗(湖北林业局林木种苗管理总站内),2009年移植到湖北省林木种苗场(湖北省咸宁市咸安区横沟桥镇)再培养1 a。2010年杂交子代测定林营建于湖北省咸宁市咸安区林业科学研究所,移植再培养1 a实生苗造林,平均苗高0.8 m左右,面积4 hm2,随机完全区组设计,3次重复,10株小区,株行距3 m×4 m,63个杂交组合,以7个鹅掌楸和北美鹅掌楸母本自由授粉家系为对照[36]。
1.3数据调查
2009年底进行苗木地径、苗高及保存率调查,造林后(2010~2020年)每年生长季节结束后进行胸径、树高及其干型等性状的调查。B76C6AAF-029F-4CDF-B006-78D12E6F5DD1
1.4数据整理与分析
数据分析对象为鹅掌楸属种间杂交正反交47个组合,应用Excel 2010进行数据整理、分析和制图,材积计算以V=f×π×H×(D/2)2,式中H表示树高,D表示胸径(苗期时是地径),f是实验形数,取值为1/3[36]。变异系数cv=s/x-×100%,s表示标准差,x-表示平均值;相关系数r=cov(x·y)/var(x)·var(y),式中cov(x·y)表示x与y的协方差,var(x)表示x的方差,var(y)表示y的方差。方差分析和相关分析运用SPSS17.0进行。
2结果与分析
2.1生长性状差异性分析
鹅掌楸属种间杂交子代测定林12 a生(第1年为苗期,下同)生长数据见表1,平均胸径23.3 cm,变异系数为26.8%,平均树高14.50 m,变异系数为21.1%,平均材积为0.229 m3,变异系数为58.9%,由此可以看出杂交组合间存在较大的分化。连年生长数据方差分析结果见表2,胸径、树高和材积3个指标的方差分析结果极为相似,所有年份杂交组合间的生长性状差异极显著,杂交组合间存在广泛的遗传变异,为优良杂交组合的选择提供了基础。
2.2生长性状连年相关分析
研究生长性状各年份之间的相关状况,了解各年份之间的内在关系,为早期选择提供合理的理论依据。由表3可知,生长性状的相关趋势是相同的,年龄相差越小,相关系数越大;随着林龄的增长,早晚相关系数也越来越大。第1年与第12年胸径、树高、材积相关系数分别为0.491、0.508、0.518,到第6年、第7年与第12年胸径、树高和材积的相关系数分别为上升到0.868、0.865、0.886和0.903、0.911、0.914,表明亚美马褂木生长前几年生长不稳定,它们之间的相关性相对较小,随着林龄的增长,相关越来越密切,进行早期选择可靠性高,因此鹅掌楸属种间杂交子代可以在第7年进行早期选择。
2.3生长动态分析
2.3.1胸径生长动态分析
第1年为造林时苗期生长数据,第2年胸径生长较慢,第3年进入速生期,第6年达到高峰,随后慢慢回落(表4)。连年生长量和平均生长量在第9年交叉(图1),表明在株行距3 m×4 m种植条件下,胸径生长在第9年达到数量成熟。虽然达到了数量成熟,但仍然保持较高的生长速度,连年生长量超过1 cm/a。
2.3.2树高生长动态分析
树高生长动态与胸径相似,第2年生长较慢,第3年进入速生期,第7年达到高峰,随后慢慢回落(表4)。连年生长量和平均生长量在第9年、第10年之间交叉(图2),表明树高生长在第10年达到数量成熟,之后树高生长明显放缓。
2.3.3材积生长动态分析
材积生长1~3 a生长缓慢,第4年进入速生期,第10年达到高峰(表4),之后有所回落。连年生长量和平均生长量在12 a内没有交叉(图3),表明材积生长还没有达到数量成熟。由于材积是树高和胸径的3次方程,树高和胸径的微小变化将引起材积较大的变化,因此生长后期材积的增长仍然很明显。
3结论与讨论
(1)育种学家们将生长期分为若干阶段,探讨各个阶段对生长量的重要性[37],速生期是指某一基因型在生长期中处于优势的时期,具有相对意义。与传统的利用树干解析木2 a为1个龄级的方法进行生长动态分析不同,本研究利用亚美马褂木12 a连年生长数据进行分析,得出亚美马褂木速生期为第3年~第9年,胸径和树高生长第3年就进入速生期,表明亚美马褂木幼苗缓苗期只有1 a,适应环境能力较强。材积生长第5年进入速生期,相对较晚,与材积达到数量成熟较晚相吻合。郝自远等[33]对24 a生北美鹅掌楸的研究结果为胸径速生期为4~16 a,而且速生期内有2个峰值;树高生长速生期为0~16 a,速生期内有3个峰值;材积生长在20 a时出现峰值,但24 a试验期内未达到数量成熟,与本试验结果不同,可能是由于树种、立地条件、生长年限等不同所致。
(2)林木生长规律是树种生物学研究的重要内容之一,为提高鹅掌楸属林分的科学经营水平,提高森林资源的储备和林地生产力,必须准确地了解林分的生长动态规律。根据本试验结果,胸径和树高的数量成熟龄分别为第9年和第10年,而材积生长在现有试验期(12 a)没有达到数量成熟。这个试验结果与郝自远等[33]关于北美鹅掌楸胸径、树高数量成熟龄为17 a,材积数量成熟齡大于24 a不同;也与朱祥锦[34]关于鹅掌楸胸径、材积数量成熟龄超过12 a,树高数量成熟龄为11 a不同。
(3)三个生长性状连年相关具有相似的规律,早晚相关随着林龄的增长,相关系数越来越大,在速生期的末期(第7年)相关系数已经超过0.9,以后的相关系数逐渐接近1.0,表明在这个时期进行早期选择的结果具有很高的可靠性。关于鹅掌楸属树种生长性状早晚相关的报道极少,该结果与郝自远等[33]以18 a为伐期年龄进行早期选择的结果基本一致,第6年选择效率最高。李斌等[38]通过17 a鹅掌楸人工林材性研究结果表明材性性状的早期选择不应早于7 a,Ryu 等[39]对北美鹅掌楸成熟林分的生长性状综合分析评价的结果是20 a是北美鹅掌楸早期选择的最适年龄。
(4)树木速生期、数量成熟受到树种、基因型、立地条件、栽培措施、林分密度等影响[40],国内外对于鹅掌楸属的早期选择年龄、数量成熟龄等的结论尚不统一。在进行鹅掌楸属树种生长动态分析时,首先要确定目的树种、基因型,在此基础上再结合培育目标,选择尽量相同的立地条件、栽培措施,最终得出相对准确的生长规律。
参考文献
[1]王章荣.鹅掌楸属树种杂交育种与利用[M].北京:中国林业出版社,2004.
[2]向其柏,王章荣.杂交马褂木的新名称—亚美马褂木[J].南京林业大学学报(自然科学版),2012,36 (2):1-2.B76C6AAF-029F-4CDF-B006-78D12E6F5DD1
[3]产工业学院林学系育种组.亚美杂种马褂木的育成[J].林业科技通讯,1973(12):10-11.
[4]叶金山.鹅掌楸杂种优势的生理遗传基础[D].南京:南京林业大学,1998.
[5]李周岐,王章荣.鹅掌楸属种间杂交可配性与杂种优势的早期表现[J].南京林业大学学报,2001,25 (2):34-38.
[6]汪建亚,宋开秀,王健,等.鹅掌楸属杂交育种研究初报[J].湖北林业科技,2012(5):1-6,42.
[7]杨志成.杂种马褂木扦插试验初报[J].林业科学研究,1994,7(6):697-700.
[8]叶金山,季孔庶,王章荣.杂种马褂木无性系插条生根能力的遗传变异[J].南京林业大学學报,1998,22 (2):71-74.
[9]陈金慧,施季森,诸葛强.杂交鹅掌楸体细胞胚胎发生与植株再生[J].植物生理学通讯,2002(6):7-12.
[10]蒋祥娥,蔡桁,汪健亚,等.鹅掌楸组织培养技术初探[J].江西林业科技,2004(4):24-26.
[11]袁金伟,孙笃玲.杂交马褂木嫁接技术[J].林业科技开发,2004,18(3):66-67.
[12]赵书喜.杂交马褂木的引种与杂种优势利用[J].湖南林业科技,1989(2):20-21.
[13]武慧贞.杂交马褂木引种试验[J].湖北林业科技,1990(3):16-18.
[14]李建民,封剑文,谢芳,等.鹅掌楸人工林的丰产特性[J].林业科学研究,2000,13(6):622-627.
[15]胡少瑛,芮义忠,冯必胜.杉木与鹅掌楸混交林效益调查研究[J].安徽林业科技,2001(6):14.
[16]李斌.北美鹅掌楸引种驯化与良种选育[C].第14次全国树木引种驯化学术研讨会暨第3届全国灌木学术研讨会,2012,
[17]何长虹.亚美马褂木施肥效应研究[D].南宁:广西大学,2015.
[18]王章荣,高悍东.亚美马褂木在我国丘陵山区的造林示范与推广[J].林业科技开发,2015,29(5):1-4.
[19]王晓东,唐良民,徐红波.造林密度对亚美马褂木生长的影响[J].南方林业科学,2018,46(3):28-30.
[20]张晓平,方炎明,陈永江.淹涝胁迫对鹅掌楸属植物叶片部分生理指标的影响[J].植物资源与环境学报,2006,15(1):41-44.
[21]孙志勇,季孔庶.干旱胁迫对杂交鹅掌楸无性系叶片内源激素含量的影响[J].安徽农业科学,2010,38 (31):17362-17364.
[22]成铁龙,彭冶,施季森,等.低温胁迫对杂交鹅掌楸幼苗活性氧和活性氮代谢的影响 [J].江苏农业科学,2017,45(24):99-102.
[23]周艳威,李美平,鲁路,等.杂交鹅掌楸苗期NaCl胁迫响应研究 [J].分子植物育种,2017,15(2):735-743.
[24]徐朝阳,潘彪.人工林杂种鹅掌楸木材性能及其应用[J].中国木材,2004(3):20-22.
[25]季孔庶,王章荣,温小荣.杂交鹅掌楸生长表现及其木材胶合板性能[J].南京林业大学学报:自然科学版,2005,29(1):71-74.
[26]潘彪,徐朝阳,王章荣.杂交鹅掌楸木材解剖性质及其径向变异规律[J].南京林业大学学报(自然科学版),2005,29(1):79-82.
[27]徐嘉娟,李火根.鹅掌楸LcPAT8基因的克隆及功能初步分析[J].林业科学,2017,53(9):45-54.
[28]成彦丽,王曦,司卫杰,等.鹅掌楸属GST家族基因的克隆与表达分析[J].西北植物学报,2018,38(3):445-455.
[29]刘换换,杨立春,张成阁,等.北美鹅掌楸CPP转录因子家族LtTCX2基因的克隆与分析[J].广西植物,2020,40(7):9981009.
[30]张成阁,刘换换,宗亚仙,等.美鹅掌楸LtuFPPS1基因克隆与组织表达分析[J].广西植物,2021,41(6):961-969.
[31]王章荣.鹅掌楸属(Liriodendron)杂交育种回顾与展望[J].南京林业大学学报(自然科学版),2003,27(3):76-78.
[32]蔡伟建,杨丽芳,高捍东,等.杂交马褂木人工林生长规律及趋势研究[J].安徽农业科学,2011,39(15):945794-59.
[33]郝自远,李火根,康昊,等.北美鹅掌楸人工林生长规律及早期选择可行性探究[J].林业科学研究,2017,30(5):8788-85.
[34]朱祥锦.鹅掌楸人工林生长最优模型选择研究[J].亚热带植物科学,2020,49(1):46-51.
[35]Bruce L G,Nuno M G B,Carolyn A R. CA.Early selection in eucalypt breeding in Australia – optimum selection age to minimise the total cost of kraft pulp production[J].New Forests,2003,25(3):201-210.
[36]管兰华,胡兴焕,曹健,等.鹅掌楸属杂交测定初步分析研究[J].湖北林业科技,2015,44(3):16-19.
[37]Wu H X. Study of early selection in tree breeding. 2. Advantage of early selection through shortening the breeding cycle.[J].Silvae Genetica,1999,48(2):78-83.
[38]李斌,顾万春.鹅掌楸主要木材性状早期选择可行性研究[J].林业科学,2002,38(6):43-48.
[39]Ryu K O,Hang M S,et al. Ageage correlation on volume growth of yellow poplar(Liriodendron tulipifera L.)[J].Journal of Agriculture & Life Science,2014,48(3):1323.
[40]Mckeand S E. Optimum age for family selection for growth in genetics tests of lobolly pine[J].Forest Science,1998,34(2):400-411.
(责任编辑:唐岚)
基金项目:湖北省林业科技支撑重点项目“湖北省优良乡土珍稀树种选育体系建设”([2017]LYKJ04)。
作者简介:蔡桁(1964~),女,高级工程师,主要从事组织培养和林木育种工作。
管兰华为通讯作者。B76C6AAF-029F-4CDF-B006-78D12E6F5DD1
