
茶尺蠖无机焦磷酸酶EoPPase672的表达与功能研究
2022-05-19 06:05傅子卓杨晓霞袁东雪毛新芳刘忠渊
昆虫学报 2022年4期
尹 恒, 傅子卓, 杨晓霞, 袁东雪, 毛新芳, 刘忠渊, *
(1.四川轻化工大学化学工程学院, 四川自贡 643000; 2.新疆大学生命科学与技术学院, 乌鲁木齐 830046)
无机焦磷酸酶(inorganic pyrophosphatase, PPase)最先在酵母中纯化得到(Kunitz, 1952),可以将多种代谢物中的焦磷酸基团水解成磷酸基团,在机体的正常物质代谢和能量传递中起到重要作用(Harold, 1966; Sivulaetal., 1999; Nelson and Cox, 2000; Poole and Reeve, 2005)。无机焦磷酸酶具有较高催化效率,可使焦磷酸盐(pyrophosphate, PPi)的水解速率提高1010-11倍,可以在短时间内降低PPi基团的积累,保证大分子持续合成,促进机体发育(Oksanenetal., 2007; 吴群峰, 2014)。无机焦磷酸酶可根据蛋白溶解性分为膜结合无机焦磷酸酶和可溶性无机焦磷酸酶。膜结合无机焦磷酸酶主要存在于植物、细菌以及寄生性生物中,可作为质子泵和Na+泵,在生物能的保存和传递中发挥重要功能(李小园等, 2004)。对水稻、细菌的无机焦磷酸酶研究表明,无机焦磷酸酶在反应过程中可以释放能量,推动热力学平衡向生物合成方向进行,对合成反应具有促进作用,在ATP缺乏的情况下可应对外界胁迫(黄园波等, 2013; 张亚芳等, 2013)。可溶性焦磷酸酶主要存在于胞质内,广泛分布于动物、植物以及真菌体内,具有调节焦磷酸基团和磷酸基团比例,维持正常生命活动的作用。朱家红等(2013)在橡胶中分离得到3个可溶性无机焦磷酸酶蛋白,证明其可以抑制PPi积累,促进橡胶的生物合成,提高橡胶的产量(Prévtetal.,1987)。对黑腹果蝇Drosophilamelanogaster焦磷酸酶研究发现其可以促进染色体的复制和转录(Gdulaetal., 1998);对线虫和蛔虫体内焦磷酸酶深入研究证明,其与幼虫发育息息相关(Islametal., 2003; Koetal., 2007)。
茶尺蠖Ectropisobliqua是一种鳞翅目(Lepidoptera)尺蠖蛾科(Geometridae)的昆虫,俗称拱背虫、造桥虫等,是茶园中发生普遍、为害严重的害虫之一(张汉鹄, 2001)。其具有暴食性,早期幼虫咬食嫩叶、嫩梢,后期取食量增加常将叶片咬食形成缺刻,大面积发生时可以将叶片全部吃光,茶园如同火烧一般,无茶可采,导致树势衰弱,对茶叶的品质和产量具有重大影响,常年造成茶叶产量损失约15%(赵磊等, 2014)。目前对于茶尺蠖体内相关酶的报道较少,仅有茶尺蠖脂肪酶合成基因EoL2的克隆(杜琴等, 2015)与利用短稳杆菌Empedobacterbrevis防治茶尺蠖后茶尺蠖中肠的血淋巴内解毒酶(AchE, CarE和GSTs)的应激情况(李良德等, 2020)的研究,更多的则是对于茶尺蠖的防治开展的一些研究(李喜旺等, 2017; 张帅琪等, 2020)。
此外,相关研究主要集中在植物和细菌中(刘海英等, 2008),昆虫无机焦磷酸酶报道较少,从NCBI中可以检索到很多鳞翅目昆虫无机焦磷酸酶基因,但是对其进行深入研究的鲜有报道。另外,已有报道表明焦磷酸酶可作为PCR增强剂(王磊, 2014)。PCR增强剂的发展源于PCR在扩增过程中会出现扩增效率低、扩增量少等问题。PCR反应一般设置30~35个循环,循环次数过多会造成大量副产物如PPi的积累,产物积累速度减慢,非特异性条带的扩增概率增加,而PCR增强剂的使用可以解决这些问题。无机焦磷酸酶具有水解PPi的特性,因此研究无机焦磷酸酶对PCR的增强效果具有重要意义。本研究通过茶尺蠖转录组测序数据库中获得无机焦磷酸酶EoPPase672的基因序列,进行体外表达和功能验证,为进一步研究无机焦磷酸酶在茶尺蠖解毒过程中的作用机制奠定基础。
1 材料与方法
1.1 供试昆虫
研究所用茶尺蠖卵来源于中国农业科学院茶叶研究所,由本实验室进行室内人工饲养和繁殖,饲养温度为22±0.5℃,相对湿度为75%±5%,光周期为14L∶10D,幼虫饲喂新鲜的茶叶枝。
1.2 材料与试剂
大肠杆菌Escherichiacoli菌株DH5α和BL21(DE3)、DNA Marker、Taq酶、pUCm-T载体和胶回收试剂盒购自生工生物工程(上海)股份有限公司;Transzol试剂、逆转录试剂和质粒DNA小量抽提试剂盒购自北京天根生物科技有限公司;ProteinFind®Anti-His Mouse Monoclonal Antibody和 ProteinFind®Goat Anti-Mouse IgG(H+L), HRP Conjugate购自北京全式金生物技术有限公司;pET32a质粒载体为实验室保存;溴氰菊酯(50 mL/瓶)购自浙江威尔达化工有限公司;其他未标明试剂均为国产分析纯。
1.3 生物学信息分析
通过茶尺蠖转录组数据(Yinetal., 2021)获得EoPPase672基因cDNA序列,利用在线预测软件对EoPPase672理化性质进行预测。 通过在线程序ORF Finder(http:∥www.ncbi.nlm.nih.gov/projects/gorf)预测基因序列的开放阅读框;使用在线软件ProtParam(http:∥web.expasy.org/protparam/)预测蛋白分子量和等电点;使用ProtScale (http:∥web.expasy.org/protscale/)预测蛋白亲疏水性;使用在线软件SignalIP 4.1(http:∥www.cbs.dtu.dk/services/SignaIP-4.1/)分析蛋白的信号肽位点。利用InterPro(http:∥www.ebi.ac.uk/interpro/scan.html)在线分析软件预测蛋白质中包含的结构域;利用PredictProtein(https:∥www.predictprotein.org/home)在线预测蛋白的二级结构。从NCBI数据库中下载昆虫无机焦磷酸酶的氨基酸序列,利用MEGA 5软件的邻接法(neighbor-joining, NJ)构建茶尺蠖EoPPase672和其他物种PPase的系统发育树。
1.4 重组蛋白EoPPase672的表达和纯化
将实验室保存的重组质粒pET-32a-EoPPase672转入大肠杆菌BL21(DE3)感受态细胞中,挑选单克隆菌落37℃ 220 r/min过夜培养,按照1∶100(v/v)转接入10 mL新鲜LB培养基中,活化培养4 h(OD600=0.6~0.8)后,加入诱导剂IPTG(终浓度为0.5 μmol/L),37℃ 220 r/min诱导蛋白表达。12 000 r/min离心1 min收集菌体。12%的SDS-PAGE检测。
将阳性单克隆菌液接入1 L新鲜LB培养基(含Amp+)进行大量诱导,离心收集菌体;用Binding Buffer (20 mmol/L Tris-HCl, 150 mmol/L NaCl, 10 mmol/L Imidazole, pH 8.0)重悬菌体,超声破碎后离心后收集上清;将上清液加入镍柱中,旋转摇床低温结合2 h。收集流穿液,用含不同浓度咪唑 (20, 25, 50和200 mmol/L)的洗脱溶液洗脱柱子,收集洗脱液,12%的SDS-PAGE检测。
在低温条件下,将纯化后蛋白洗脱液装入透析袋,在透析液(20 mmol/L Tris-HCl)中4℃透析48 h(12~16 h换1次透析液)。用10 kD的超滤管浓缩蛋白,12% SDS-PAGE检测蛋白纯度,Bradford法定量蛋白浓度。
1.5 Western bolt鉴定重组蛋白EoPPase672
将1.4节纯化的重组蛋白EoPPase672样品进行SDS-PAGE检测,将分离胶部分进行转膜(冰浴条件下,20 V恒压转移1 h),用含5%脱脂奶粉的TBST,37℃封闭NC膜4 h。用含3%脱脂奶粉的TBST按1∶5 000(v/v)稀释一抗(Mouse Anti His-Tag MonoClonal antibody),37℃孵育2 h,洗膜5次;1∶3 000 (v/v)稀释二抗(Horseradish Peroxidase-conjugated AffiniPure Goat Anti-Mouse IgG),37℃孵育1.5 h,洗膜5次;DAB染色液避光染色5 min后观察显色情况。
1.6 溴氰菊酯刺激后茶尺蠖EoPPase672的表达水平检测
通过毒力实验测定,测得溴氰菊酯对茶尺蠖的亚致死浓度(LC10=2.08 mg/L),使用(LC10)溴氰菊酯浸泡茶叶10 s后饲喂茶尺蠖2-4龄幼虫,分别在0(CK), 12, 24和48 h后取试虫,一个处理条件处理6头同龄昆虫,液氮冷冻保存,使用Transzol法提取幼虫总RNA,反转录合成cDNA模板。用Primer Premier 5设计引物序列,以18S rRNA(正向引物: 5′-GAGAAACGGCTACCACATCCA-3′; 反向引物: 5′-GCAAATGCTTTCGCTGATGTT-3′)作为内参基因,基于茶尺蠖转录组数据获得的EoPPase672基因cDNA序列的保守性设计相应引物(正向引物: 5′-CCACACAGTTTTCCGCTACA-3′; 反向引物: 5′-TATGCTCACCAAGCAGCATC-3′),qRT-PCR检测溴氰菊酯刺激后不同时间点EoPPase672的表达水平。PCR反应体系:2×SYBR Green Mix 12.5 μL,正反向引物(10 μmol/L)各2.5 μL, cDNA模板1.0 μL, RNase-Free Water 6.5 μL。反应条件: 95℃ 30 s; 95℃ 10 s, 60℃ 30 s, 40个循环。循环结束后添加溶解曲线。每个处理3个生物学重复,每个重复来自2头个体,3个技术性重复。
1.7 茶尺蠖EoPPase672酶活力测定
配制Na2HPO4(0.1 mol/L)溶液,取用不同量(0, 150, 250, 350, 450, 550, 600, 750和850 μL)加H2O定容至25 mL混匀,每份取240 μL并加入2 mL AAM[体积比为H2O∶正钼酸铵(20 mmol/L)∶硫酸(2 mmol/L)∶丙酮=0.12∶0.5∶1.25∶2]静置显色3 min后于420 nm下比色测定,制作磷含量标准曲线。实验组依次加入20 μL焦磷酸钠(0.017 mol/L), 500 μL P Buffer[0.06 g Tris-HCl, 0.003g MgSO4(1 mmol/L)定容于100 mL],50 μL H2O,1.4节原核表达纯化后的茶尺蠖EoPPase672酶液(0.28 mg/mL) 20 μL,对照组用H2O替代,65℃水浴30 min,立即加入100 μL柠檬酸(0.1 mmol/L)溶液终止反应,冷却至室温取240 μL加入2 mL AAM,静置显色3 min后于420 nm下比色测定磷酸根浓度,利用标准曲线算出溶液中无机磷酸盐(Pi)的浓度,进一步测定EoPPase672酶活力。将1.4节原核表达纯化后的茶尺蠖EoPPase672酶液稀释200倍后,按上述酶活力测定的方法测定不同pH(4.0, 4.5, 5.0, 5.5, 6.0, 6.5, 7.0, 7.5, 8.0, 8.5, 9.0, 9.5, 10.0和10.5)下的酶活力[1 个酶活力单位(U)为1 μmol EoPPase672在65℃时,1 min内转化1 μmol底物所需的酶量],制作酶活力与pH相关曲线,测定酶反应最适pH。将1.4节原核表达纯化后的茶尺蠖EoPPase672酶液稀释200倍后,按上述酶活力测定的方法测定不同温度下(25, 35, 45, 55, 65, 75, 85和95℃)酶活力,制作酶活力与温度相关曲线,测定酶反应最适温度。用不同金属离子(Ca2+, Cu2+, Mn2+和Zn2+)替换P Buffer中的Mg2+,检测添加不同金属离子与Mg2+共同作为辅因子时重组蛋白EoPPase672的活力,验证不同金属离子对酶活力的影响。
1.8 重组蛋白EoPPase672在PCR反应中对PPi的去除效果检测
为了研究PCR反应过程中产生的焦磷酸基团对PCR扩增的抑制效果,称取0.003 g焦磷酸钠配制4 mmol/L焦磷酸钠溶液,依次稀释为0, 0.25, 0.5, 1, 2和4 mmol/L后加入PCR反应中,基于EoPPase672序列设计正反向引物(5′-GCATTATCG CCGCGCGTGTT-3′; 5′-GAGATTGGAAGCGAGGTAG-3′),以pET-32a-EoPPase672质粒DNA为模板进行PCR。PCR反应体系(20 μL): Taq 9 μL,正反向引物(10 μmol/L)各0.5 μL, DNA模板1.0 μL, ddH2O 8.0 μL, PPi 1.0 μL。反应程序: 94℃ 4 min; 94℃ 50 s, 57℃ 40 s, 72℃ 40 s, 32个循环; 72℃ 10 min。1%琼脂糖凝胶电泳检测PCR结果。
以只添加不同浓度的PPi(0, 0.4, 0.8, 1.0, 1.2, 1.4, 1.6和1.8 mmol/L)为对照组,实验组中添加与对照组中相同浓度的PPi和1 μL重组蛋白EoPPase672(1.0 mg/mL),检测不同浓度的PPi对PCR扩增效率的影响以及重组蛋白EoPPase672对PPi抑制效果的影响。基于EoPPase672序列设计正反向引物(5′-GCATTATCGCCGCGCGTGTT-3′; 5′-GAGATTGGAAGCGAGGTAG-3′),以pET-32a-EoPPase672质粒DNA为模板进行PCR。PCR反应体系(20 μL): Taq 9 μL, 正反向引物(10 μmol/L)各0.5 μL, pET-32a-EoPPase672质粒DNA模板1.0 μL, ddH2O 8.0 μL, PPi 1.0 μL。反应程序: 94℃ 4 min; 94℃ 50 s, 57℃ 40 s, 72℃ 40 s, 32个循环; 72℃ 10 min。1%琼脂糖凝胶电泳检测PCR结果。
1.9 重组蛋白EoPPase672对PCR扩增效率的影响检测
在实验组中依次加入不同剂量的Taq酶(10, 15, 20, 25和30 U)和重组蛋白EoPPase672(1.0 mg/mL) 1 μL,以只含有不同浓度Taq酶(10, 15, 20, 25和30 U)的PCR作为对照组,进行PCR,引物序列同1.8节,分析重组蛋白EoPPase672对PCR扩增效率的影响。PCR反应体系(20 μL): Taq 9 μL, 正反向引物(10 μmol/L)各0.5 μL, DNA模板1.0 μL, ddH2O 7.0 μL, PPi 1.0 μL,重组蛋白EoPPase672(1.0 mg/mL) 1.0 μL。反应程序: 94℃ 4 min; 94℃ 50 s, 57℃ 40 s,72℃ 40 s 32个循环; 72℃ 10 min。1%琼脂糖凝胶检测PCR结果。
1.10 数据分析
基因相对表达量的计算采用2-ΔΔCT法,利用IBM SPSS Statistics 2分析基因相对表达量数据,采用单因素方差分析和LSD多重比较分析差异显著性。
2 结果
2.1 茶尺蠖EoPPase672序列分析
从茶尺蠖转录组数据库筛选到EoPPase672 cDNA(GenBank登录号: OM962968),其开放阅读框全长1 023 bp,编码340个氨基酸。其编码蛋白PPase672预测相对分子质量为37.8 kD,等电点为6.27,亲水性指数为-0.326,推测为亲水性蛋白。无信号肽,为分泌型蛋白。
氨基酸序列比对结果表明(图1),昆虫的PPase结构域序列保守。茶尺蠖EoPPase672与家蚕BombyxmoriBmPPase的序列一致性最高,为80%,与粉纹夜蛾TrichopiusianiTnPPase的序列一致性为76%,与斜纹夜蛾SpodopteralituraSlPPase的序列一致性为76%,与棉铃虫HelicoverpaarmigeraHaPPase的序列一致性为77%。系统发育树表明茶尺蠖EoPPase672与鳞翅目昆虫夜蛾类中的粉纹夜蛾TnPPase的亲缘关系最近,与鞘翅目的赤拟谷盗TriboliumcastaneumTcPPase亲缘关系最远(图2)。

图1 茶尺蠖EoPPase672与其他昆虫PPase氨基酸序列比对Fig.1 Amino acid sequence alignment of EoPPase672from Ectropis obliqua and PPase from other insects蛋白来源物种及GenBank登录号Origin species of proteins and their GenBank accession numbers: EoPPase672: 茶尺蠖Ectropis obliqua, OM962968; TnPPase: 粉纹夜蛾Trichopiusia ni, XP_026737068.1; SlPPase: 斜纹夜蛾Spodoptera litura, XP_022835395.1; HaPPase: 棉铃虫Helicoverpa armigera, XP_021199627.1; HvPPase: 烟芽夜蛾Heliothis virescens, PCG63030.1; BmPPase: 家蚕Bombyx mori, XP_004922937.1; VtPPase: 特美红蛱蝶Vanessa tameamea, XP_026484258.1; DppPPase: 黑脉金斑蝶Danaus plexippus plexippus, XP_032513218.1; AtPPase: 脐橙螟Amyelois transitella, XP_013184525.1; PxPPase: 小菜蛾Plutella xylostella, CAG9132134.1; GmPPase: 大蜡螟Galleria mellonella, XP_026753285.1.
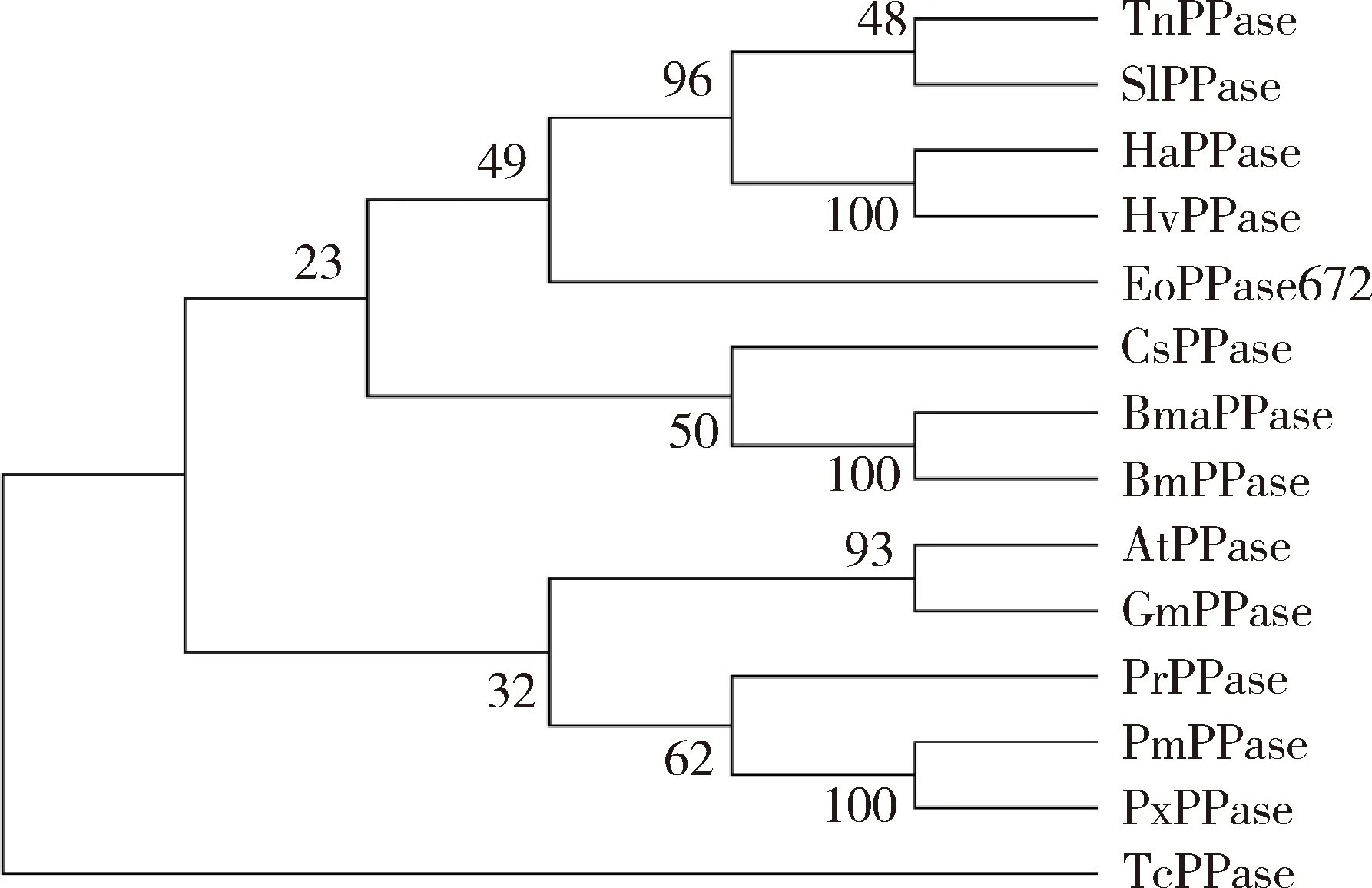
图2 邻接法构建的基于氨基酸序列的茶尺蠖和其他昆虫PPase672的系统进化树(1 000次重复)Fig.2 Phylogenetic tree of PPase672 from Ectropis obliquaand other insects by neighbor-joining method based onamino acid sequences (1 000 replicates)蛋白来源物种及GenBank登录号Origin species of proteins and their GenBank accession numbers: TnPPase: 粉纹夜蛾Trichoplusia ni, XP_026737068.1; SlPPase: 斜纹夜蛾Spodoptera litura, XP_022835395.1; HaPPase: 棉铃虫Helicoverpa armigera, XP_021199627.1; HvPPase: 烟芽夜蛾Heliothis virescens, PCG63030.1; EoPPase672: 茶尺蠖Ectropis obliqua, OM962968; CsPPase672: 水稻二化螟Chilo suppressalis,RVE43533.1; BmaPPase:野桑蚕Bombyx mandarina; XP_028027307.1; BmPPase: 家蚕Bombyx mori, XP_004922937.1; AtPPase: 脐橙螟Amyelois transitella, XP_013184525.1; GmPPase: 大蜡螟Galleria mellonella, XP_026753285.1; PrPPase: 菜粉蝶Pieris rapae, XP_022123308.1; PmPPase: 金凤蝶Papilio machaon, XP_014370251.1; PxPPase: 柑橘凤蝶Papilio xuthus, KPI99608.1; TcPPase: 赤拟谷盗Tribolium castaneum, XP_008200814.1.
2.2 EoPPase672的原核表达
SDS-PAGE结果如图3(A, B)所示,在50 kD处有一条表达量很高的条带,与目的蛋白大小一致。Western blot结果进一步表明重组EoPPase672蛋白的正确表达(图3: C)。将蛋白洗脱液经过透析、浓缩和定量,最终获得重组EoPPase672蛋白浓度为0.28 mg/mL,单一性较强,可用于后续功能验证。
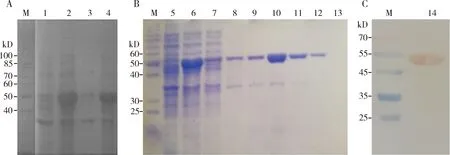
图3 重组EoPPase672蛋白表达(A)、纯化(B)的SDS-PAGE及Western blotting(C)Fig.3 SDS-PAGE detection of expression (A) and purification (B), and Western blotting (C) of the recombinant EoPPase672M: 蛋白质分子量标准Protein molecular weight marker; 1: 总菌体裂解液(未诱导)Total bacterial lysate (not induced); 2: 37℃下0.6 mmol/L IPTG诱导4 h后的总菌体裂解液Total bacterial lysate induced by 0.6 mmol/L IPTG at 37℃ for 4 h; 3: 超声上清 Ultrasonic supernatant; 4: 超声沉淀Ultrasonic precipitate; 5: 总菌体裂解液(未诱导)Total bacterial lysate (not induced); 6: 28℃ 180 r/min下0.6 mmol/L IPTG诱导4 h后的总菌体裂解液Total bacterial lysate induced by 0.6 mmol/L IPTG at 28℃ and 180 r/min for 4 h; 7: 流穿液Flow-through sample; 8-13: 分别用浓度为20, 25, 50和200 mmol/L的咪唑洗脱缓冲液洗脱获得的蛋白Protein samples washed with 20, 25, 50 and 200 mmol/L imidazole elution buffer, respectively; 14: Western blot检测的纯化重组蛋白Purified recombinant protein detected by Western blot.
2.3 溴氰菊酯刺激后茶尺蠖幼虫中EoPPase672的表达水平
结果表明(图4),溴氰菊酯(LC10=2.08 mg/L)刺激12 h后,茶尺蠖幼虫中EoPPase672表达整体情况呈现上调趋势,2和3龄幼虫中的表达量与对照组的相比显著上调(P<0.001),4龄幼虫中的小幅上调,与对照组相比无显著差异(P>0.05);24 h后,2和3龄幼虫中EoPPase672的表达量与对照组相比仍然显著上调(P<0.001),但相对于处理12 h而言,表达量呈现下调趋势。
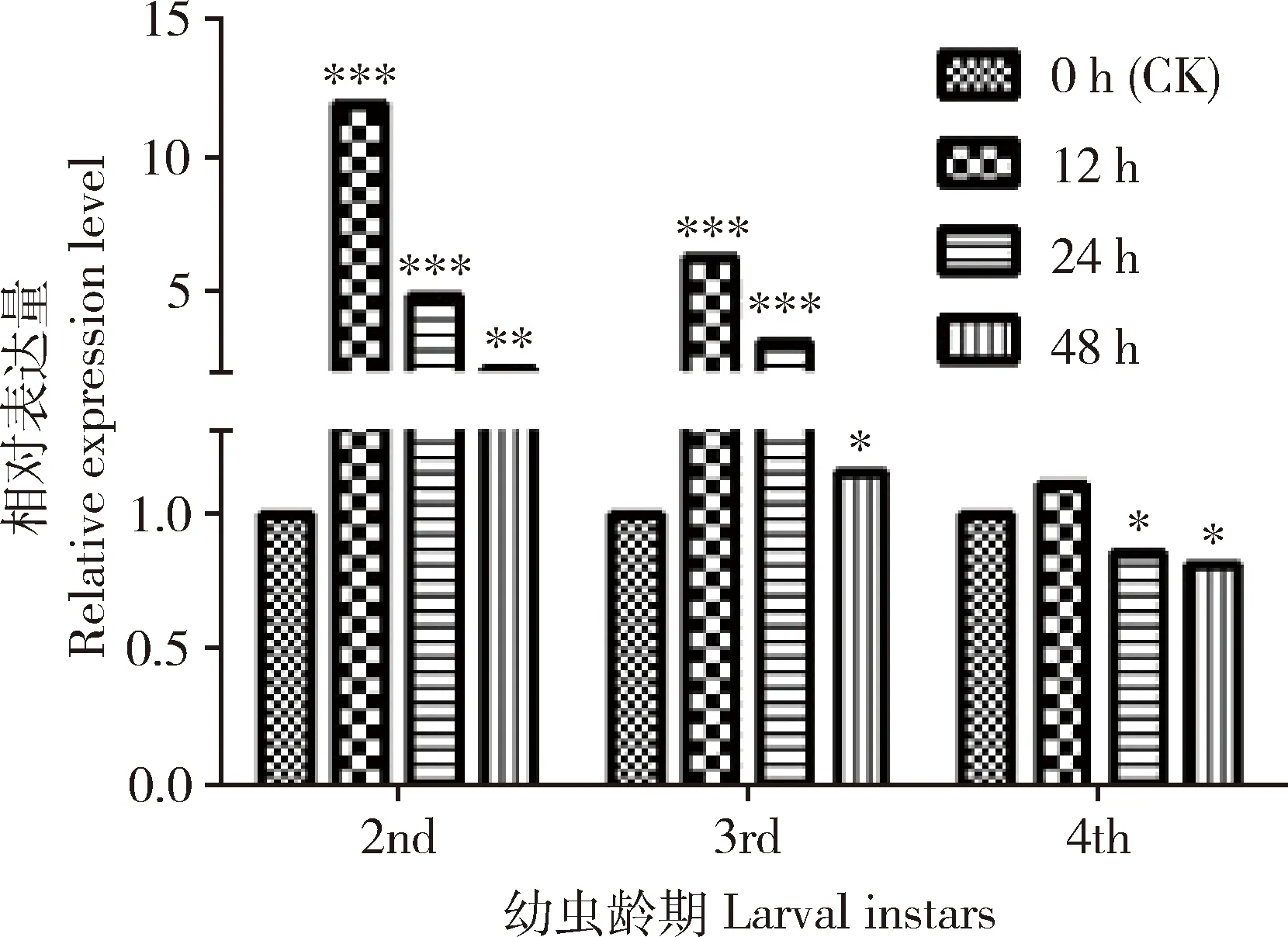
图4 溴氰菊酯(LC10=2.08 mg/L)刺激不同时间后EoPPase672在茶尺蠖不同龄期幼虫中的表达量Fig.4 Expression levels of EoPPase672 in Ectropisobliqua larvae at different instars after exposed todeltamethrin (LC10=2.08 mg/L) for different time图中数据为平均值±标准误; 柱上星号表示基因相对表达量在处理组与对照组间的差异显著性(*P<0.05; **P<0.01; ***P<0.001)(t检验)。Data in the figure are mean±SE.Asterisk above bars indicates significant difference in the relative gene expression level between the treatment group and the control group (*P<0.01; **P<0.01; ***P<0.001)(t-test).
2.4 EoPPase672酶活力
酶活力测定结果显示(图5: A),通过Pi含量标准曲线可得EoPPase672比活力为1 279.6 U/mg。pH 4-6时酶活力低,随着pH增加酶活力呈线性增加(图5: B),在靠近pH 6-7之间突然降低,推测原因可能是此pH接近蛋白质等电点,蛋白质稳定性降低,酶反应最适pH 7-8,高于pH 8之后酶活力缓慢降低,表明EoPPae672酶反应最适pH 7.5左右。热稳定性研究表明(图5: C),重组蛋白EoPPase672在20-100℃之间都有酶活力,65℃时活力最高,为酶反应最适温度,当温度低至20℃或高至95℃时相对活力都高达40%以上,表明重组蛋白EoPPase672具有很好的热稳定性。酶反应的发生需要金属离子作为辅因子,结果表明,Mg2+是最适金属离子,Zn2+和Mn2+对重组蛋白EoPPase672的酶活力具有抑制作用(图5: D)。
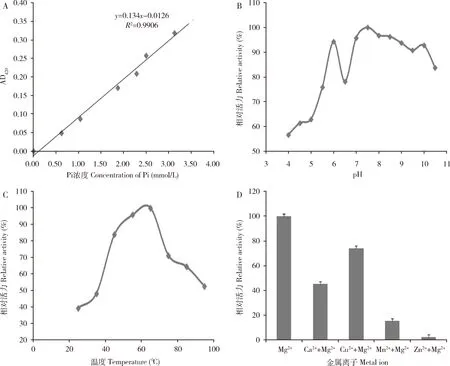
图5 重组蛋白EoPPase672的酶活力(A)以及最适pH (B)、最适温度(C)和最适金属离子(D)Fig.5 Determination of activity (A), and the optimum pH (B), optimum temperature (C)and optimum metal ion (D) of the recombinant EoPPase672利用无机磷酸盐(Pi)含量标准曲线测得酶活力。Standard curve of inorganic phosphate (Pi) content was used to determine the enzyme activity.
2.5 EoPPase672在PCR反应中对PPi去除效果
PCR反应后期,副产物的大量积累会直接影响扩增效率,为了验证PPi对PCR反应的抑制效果,外源添加焦磷酸钠到PCR反应中,结果如图6,当焦磷酸钠浓度较低(0.25和0.5 mmol/L)时,对PCR的抑制效果不明显,当焦磷酸钠浓度增加到1 mmol/L时明显抑制了扩增效率,2 mmol/L以上则完全抑制。表明当反应体系中存在大量PPi时会对扩增效率产生一定影响。在PPi浓度相同的情况下,添加1.0 mg/mL EoPPase672后泳道4(未添加EoPPase672)与3(0.4 mmol/L PPi)无显著差异。当焦磷酸钠浓度为0.8 mmol/L时,泳道6(未添加EoPPase672)扩增结果明显低于泳道5。当焦磷酸钠浓度高于 1.0 mmol/L 时,如未添加 EoPPase672的泳道8, 10, 12, 14和16所示,PCR扩增被完全抑制,添加重组蛋白EoPPase672的不受抑制,如泳道7, 9, 11, 13和15所示,扩增条带与对照组(0 mmol/L焦磷酸钠)无显著差异(图7)。说明在PCR反应中添加重组蛋白EoPPase672可以在一定程度上水解PPi,解除PPi对PCR扩增的抑制作用。
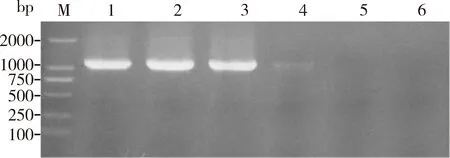
图6 不同浓度焦磷酸钠(PPi)对PCR反应的抑制作用Fig.6 Inhibition of sodium pyrophosphate (PPi) ofdifferent concentrations on PCR reactionM: DNA分子量标准DNA molecular weight marker; 1-6: 分别为0(CK), 0.25, 0.5, 1, 2和4 mmol/L PPi 0(CK), 0.25, 0.5, 1, 2 and 4 mmol/L PPi, respectively.
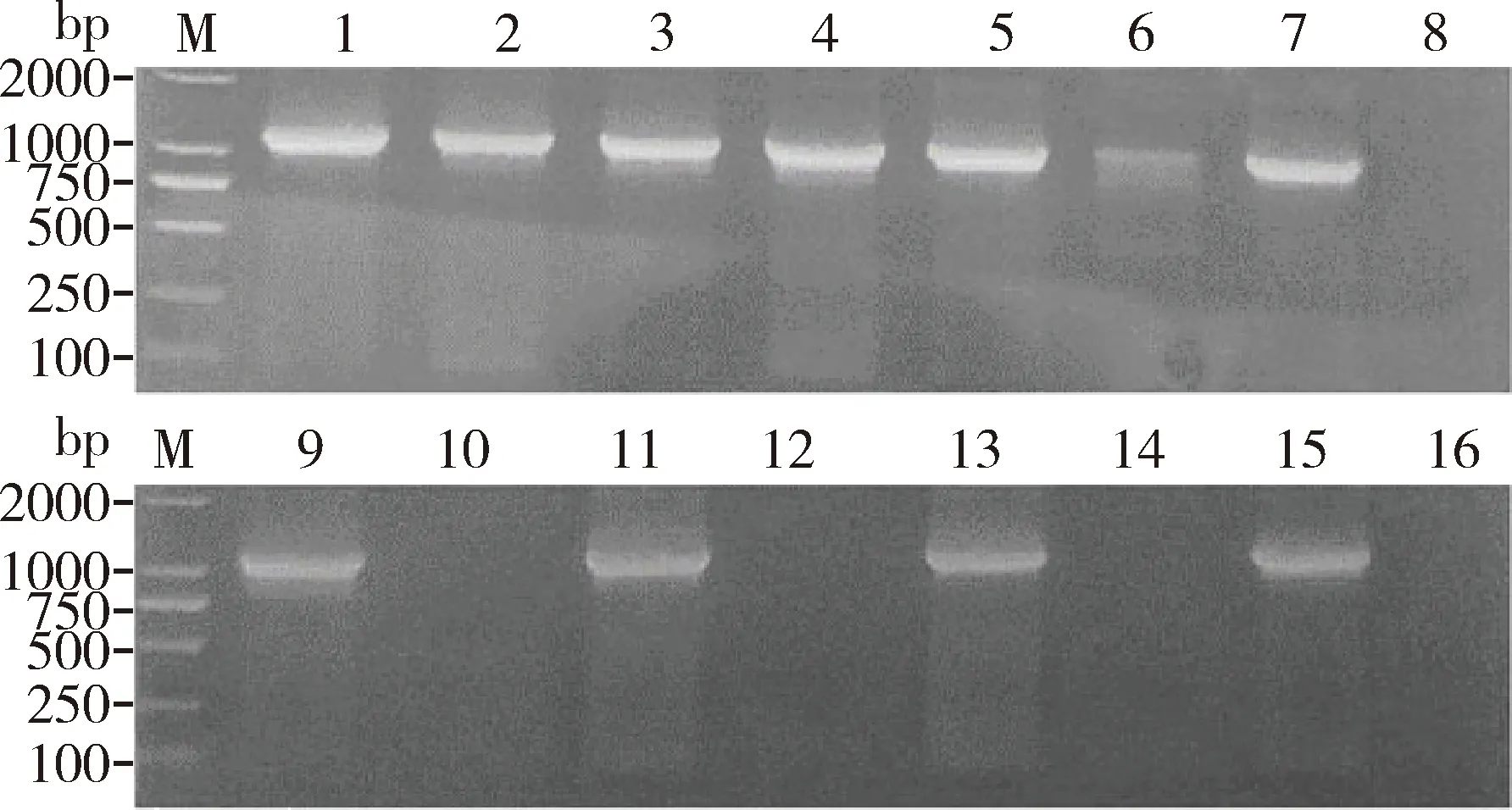
图7 重组蛋白EoPPase672 (1.0 mg/mL)在PCR反应中对焦磷酸钠(PPi)去除效果Fig.7 Effect of the recombinant EoPPase672 (1.0 mg/mL)on the removal of sodium pyrophosphate (PPi)in PCR reactionM: DNA分子量标准DNA molecular weight marker; 1: 0 mmol/L PPi+1 μL EoPPase672; 2: 0 mmol/L PPi; 3: 0.4 mmol/L PPi+1 μL EoPPase672; 4: 0.4 mmol/L PPi; 5: 0.8 mmol/L PPi+1 μL EoPPase672; 6: 0.8 mmol/L PPi; 7: 1.0 mmol/L PPi+1 μL EoPPase672; 8: 1.0 mmol/L PPi; 9: 1.2 mmol/L PPi+1 μL EoPPase672; 10: 1.2 mmol/L PPi; 11: 1.4 mmol/L PPi+1 μL EoPPase672; 12: 1.4 mmol/L PPi; 13: 1.6 mmol/L PPi+1 μL EoPPase672; 14: 1.6 mmol/L PPi; 15: 1.8 mmol/L PPi+1 μL EoPPase672; 16: 1.8 mmol/L PPi.
2.6 重组蛋白EoPPase672对PCR扩增效率的影响
结果显示(图8),泳道1为2 μL Taq酶,无明显条带,泳道2为2 μL Taq酶加1 μL重组蛋白EoPPase672,扩增结果显示有较淡条带。加入重组蛋白EoPPase672后泳道4, 6, 8和10扩增结果明显高于对照组泳道3, 5, 7和9(不加重组蛋白EoPPase672)。结果表明PCR反应体系中加入重组蛋白EoPPase672对PCR扩增效率具有增强效果。
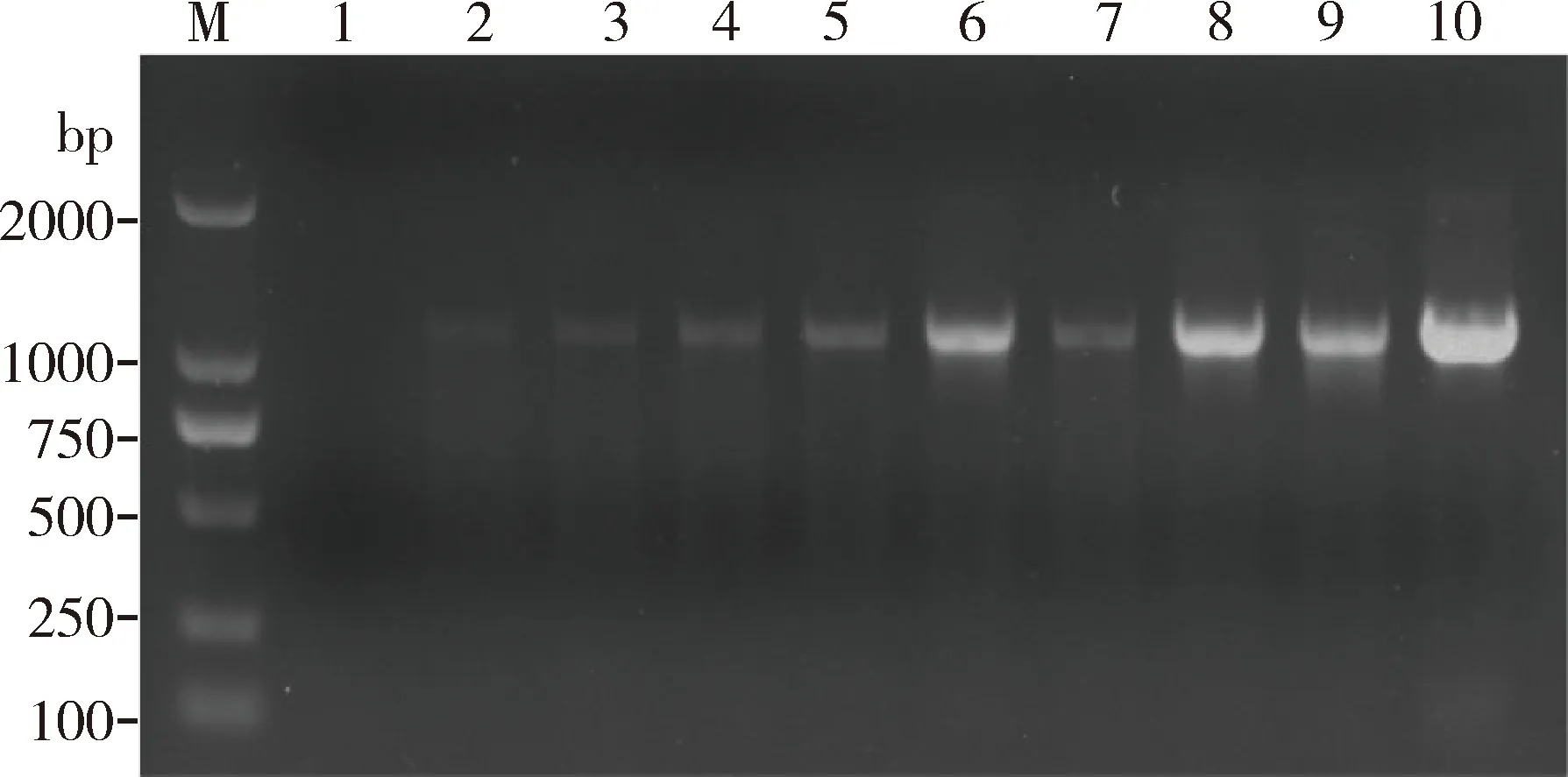
图8 重组蛋白EoPPase672 (1.0 mg/mL)对PCR扩增效果的影响Fig.8 Effect of the recombinant EoPPase672 (1.0 mg/mL)on the PCR amplification efficiencyM: DNA分子量标准DNA molecular weight marker; 1: 2 μL Taq; 2: 2 μL Taq+1 μL EoPPase672; 3: 3 μL Taq; 4: 3 μL Taq+1 μL EoPPase672; 5: 4 μL Taq; 6: 4 μL Taq+1 μL EoPPase672; 7: 5 μL Taq; 8: 5 μL Taq+1 μL EoPPase672; 9: 6 μL Taq; 10: 6 μL Taq+1 μL EoPPase672.
3 讨论
本研究从菊酯类杀虫剂处理茶尺蠖的转录组数据库中筛选到无机焦磷酸酶基因EoPPase672片段,该基因可以被杀虫剂诱导上调表达(图4)。对基因序列进行生物学信息预测,结果表明EoPPase672的ORF全长1 023 bp可编码340个氨基酸残基,推测为亲水性蛋白。通过构建原核表达载体,成功表达可溶性的蛋白(图3),在高温下稳定性强,在pH 7.5,65℃下活力最高(图5)。
在溴氰菊酯刺激茶尺蠖不同龄期幼虫后,12 h后EoPPase672表达量迅速升高(图4),推测可能昆虫对溴氰菊酯的作用反应比较迅速。比较不同龄期之间的反应程度,龄期越小的幼虫在受到外界杀虫剂入侵后更易诱导无机焦磷酸酶的表达,而龄期较高的幼虫,机体本身免疫防御能力强,在LC10浓度下对4龄幼虫的影响较低,结果提示茶尺蠖的防治在4龄幼虫之前,杀虫剂的作用效果较强,用量少。
有报道称,焦磷酸可能是辅酶α连接酶的抑制剂,这种抑制作用可以通过添加PPase来解除,该酶可以催化PPi水解无机磷酸盐(Pi),从而可以加速ATP的生成(Lelièvreetal., 2020)。此外,PPase对不同的DNA聚合酶的增强效果不同,例如对iProof, Pfu turbo, S-KT的效果明显弱于对Taq(李小园等, 2004),推测原因可能是Taq酶中的金属离子为Mg2+,在反应过程中会竞争离子,从而导致增强效果稍弱(李小园等, 2004)。金属离子是蛋白酶发挥活性的辅助因子,如果缺乏会影响到酶活力。在本研究中发现,Mg2+对EoPPase672的活力是必不可少的,而其他离子起到少量的促进或者抑制作用(图5)。当同时加入Mg2+, Cu2+或者Ca2+时,无机焦磷酸酶的活力下降,当加入Mn2+时酶活力抑制程度增强,而Zn2+几乎完全抑制。
此外,PPase在许多生物学过程中起着重要作用(Poole and Reeve, 2005; Carman and Han, 2006),并且与许多临床疾病(例如肺腺癌和结直肠癌)有关(Friedmanetal., 2004)。因此,可以建立一些简便、灵敏的PPase活力测定方法,通过测定PPase的活力,以此来预测相关的疾病。一些生物传感器的建立(Zhangetal., 2017)为PPase活力的测定带来了便利。
PCR扩增后期会进入平台期,产物积累速度减慢,酶和底物的稳定性、活力均降低,前期不断产生的焦磷酸基团大量积累,抑制扩增效率,同时存在引物二聚体或非特异性产物发生非特异性竞争,高浓度扩增产物变性不彻底和扩增产物自身复性等现象。无机焦磷酸酶可以水解PPi基团,研究发现体外表达的EoPPase672对PCR反应具有增效的作用(图8),而加入无机焦磷酸酶可以有效消除这种现象。本研究中在加入重组蛋白EoPPase672后,PCR的扩增效率得到极大的提升,说明PPase作为一种PCR扩增增强剂的可行性,其热稳定性也为发挥功能提供了保障。研究结果为茶尺蠖解毒机理的研究提供了思路与依据。
猜你喜欢
食品界(2022年10期)2022-10-19
土壤学报(2022年3期)2022-08-27
茶叶科学(2022年4期)2022-08-27
世界热带农业信息(2022年8期)2022-07-19
茶叶科学(2022年2期)2022-04-20
作物学报(2022年3期)2022-01-22
分析化学(2018年7期)2018-09-17
科学与财富(2017年12期)2017-05-16
农民致富之友·下半月(2015年6期)2015-07-09
天津农业科学(2014年12期)2014-12-11
