
鸣螽科(直翅目:螽亚目)昆虫研究历史与进展
2022-06-03 04:34顾俊杰徐自强王海建
环境昆虫学报 2022年2期
顾俊杰,徐自强,王海建,任 东
(1. 四川农业大学农学院,成都 611130;2. 首都师范大学生命科学院,北京 100037)
鸣螽科Prophalangopsidae属于直翅目Orthoptera螽亚目Ensifera原螽总科Hagloidea。最早的化石记录来自于中亚地区和中国北方的下侏罗统地层,在中生代中晚期广泛分布并达到生物多样性的高峰,随后逐渐衰落。在新生代时期在北美及中国西藏有少量发现。其现生类群多样性较低,分布地十分狭窄,仅在印度北部、中国西南地区、西伯利亚东部和北美西北部有发现。虽然原螽总科被认为很可能是并系群(Béthoux and Nel, 2002),但普遍认为该类群与螽斯总科Tettigonioidea有较近的亲缘关系(Gorochov, 2003; Jost and Shaw, 2006; Songetal., 2015)。鸣螽科作为原螽总科唯一的孑遗类群,物种多样性最为丰富,对理解螽亚目系统发育及演化历史有重要意义。Gorochov(2003)认为鸣螽科共包括5个灭绝亚科:阿博鸣螽亚科Aboilinae、原阿博鸣螽亚科Protaboilinae、赤峰鸣螽亚科Chifengiinae、螱鸣螽亚科Termitidiinae和异鸣螽亚科Tettohaglinae;两个现生亚科:鸣螽亚科Prophalangopsinae和圆翅鸣螽亚科Cyphoderrinae。本文将从分类学、生物学、进化生物学等角度回顾鸣螽科的研究历史、进展、问题及展望。
1 鸣螽科及相关类群的中文译名
Prophalangopsidae Kirby 1906作为螽亚目为数不多的孑遗类群,在系统演化中有重要地位,但其与Hagloidea Handlirsch 1906的中文名称的使用一直较为混乱。张海春等(2015)对Prophalangopsidae、Haglidae及Hagloidea的中文译名做过说明,但未明确各亚科及现生类群的中文名。昆虫分类名称使用不一致往往会带来一些负面效应(陈翠等, 2017),因此有必要厘清并明确Prophalangopsidae及相关类群的中文名称,并规范使用。名称使用的混乱与其系统位置多次发生变动、缺乏系统的研究、以及该类群现生种类十分稀少有一定的关系。Gorochov(1995)在对直翅目化石进行系统研究中提出Hagloidea总科包括Prophalangopsidae、Haglidae、Tuphellidae和Hagloedischiidae 4科,其中Prophalangopsidae包括4灭绝亚科及2现生亚科。但鸣螽Prophalangopsidae各现生类群(Prophalangopsinae、Cyphoderrinae)在研究当中常被归入原螽科Haglidae(Morris and Gwynne, 1979; Storozhenko, 1980; Kumalaetal., 2005; Judgeetal., 2011)。国内最早关于Haglidae和Prophalangopsidae的记载来自肖采瑜等人的译著《昆虫的分类》,书中将Prophalangopsidae译为鸣螽科,Prophalangopsinae译为鸣螽亚科,但并未指出Haglidae的中文名。之后部分学者沿用此中文名(林启彬, 1982; 王五力, 1987; 方艳等, 2007; 顾俊杰, 2012)。林启彬(1965)发表Haglidae两新种,但没有指明该科及属种的中文名。Haglidae最早的中文名由洪友崇先生译为哈格鸣螽(洪友崇, 1982a),此后多数古昆虫学文献中均沿用此中文名(王五力, 1987; 任东等, 1995; 王文利和刘明渭, 1996;孟祥明和任东, 2006)。张海春(1996)使用鸣螽科作为Haglidae的中文名。彩万志等(2009)在译著《昆虫学概论》中以及袁锋等(2005)都使用鸣螽科作为Haglidae的中文名,但其所指类群应是Prophalangopsidae科的Prophalangopsinae和Cyphoderrinae亚科。洪友崇曾分别使用原鸣螽亚科(洪友崇, 1982a, 1983),鸣螽亚科(洪友崇, 1982b)作为Prophalangopsinae的中文名。《昆虫分类》一书中将Hagloidea译为原螽总科,Prophalangopsinae沿用鸣螽亚科,并将模式种Prophalangopsisobscura译为暗鸣螽(刘宪伟和殷海生, 1999)。此外Prophalangopsidae还出现过其它中文名,如原树蟋科(张俊峰, 1993),原哈格鸣螽科(孟祥明和任东, 2006; 孟祥明等, 2006; 李连梅等, 2007a, 2007b)。中国动物主题数据库当中收录了两种现生Prophalangopsidae科昆虫AboilomimusguizhouensisLiuetal., 2009,AboilomimusornatusLiuetal., 2009并分别使用贵州亚鸣螽和丽亚鸣螽为中文名,高级阶元沿用鸣螽科Prophalangopsidae和原螽总科Hagloidea为中文名。此外,“鸣螽”一词也在一些常见的螽斯科Tettigoniidae物种的中文名中出现过,如将Uvarovitesinflatus译为鼓翅鸣螽(庞秉璋, 1999),将Gampsolceis译为鸣螽属(钱范俊, 1986),将Gampsolceisgratiosa译为锐声鸣螽或短翅鸣螽等(何忠和陈念丽, 1985; 沈钧贤, 1989),但所涉及“鸣螽”一词均未早于肖采瑜等人1959年在《昆虫的分类》中对Prophalangopsidae所使用的名称。
如上,Prophalangopsidae和Haglidae中文名变化甚多,造成了名称使用的混乱,以及现生类群与灭绝类群系统关系认识的混乱。因此本文将依据优先原则、使用习惯、文献考证、名称适宜度及昆虫形态特征和生物学特性等因素,建议使用如下中文名:Haglidae的大多数成员相较Prophalangopsidae出现在更为古老的地层,且表现出更为原始的特征,建议Haglidae使用原螽科、Hagloidea使用原螽总科、Haglinae使用原螽亚科作为中文名。Cyrtophyllitinae曾被译为弓叶鸣螽亚科,并与鸣螽亚科Prophalangopsinae一起归入原螽科(洪友崇, 1982b),此后又被译为弯翅螽亚科(顾俊杰, 2012),弓鸣螽亚科(张海春等, 2015)。依据其系统位置归入原螽科,以中脉明显弯曲为主要鉴别特征,同时避免产生与鸣螽科Prophalangopsidae的混淆,建议使用弯翅螽亚科作为亚科的中文名。根据优先原则、Prophalangopsidae现生类群特殊的通讯行为和鸣声,笔者认为Prophalangopsidae应沿用鸣螽科作为中文名,Prophalangopsinae为鸣螽亚科。Cyphoderrinae,其模式属词源“cyph”为驼背的、弯曲的之意,但为避免与弯翅螽亚科混淆,且Cyphoderrisspp.翅形相较鸣螽科其它类群更为短圆,建议使用圆翅鸣螽亚科为宜。Paracyphoderriserebeus原文献未提供词源,“erebeus”为地狱的、地下的之意,经与定名作者确认其种本名意指该种为穴居生活,故建议使用穴拟圆翅鸣螽亚科为其中文名。Aboilinae和Protaboilinae在现有中文文献中均译为阿博鸣螽亚科和原阿博鸣螽亚科,虽为音译,但并未产生使用混乱,建议沿用此中文名。Chifengiinae由洪友崇(1982b)建立,并译为赤峰鸣螽亚科,未产生使用混乱。Termitidiinae模式属为Termitidium,定名作者最初将其与白蚁(亦称螱)做比较(Westwood, 1854; Zeuner, 1939),故命名为Termitidium,因此建议译为螱鸣螽亚科。Tettohaglinae模式种中脉及径脉较为特殊,虽其亚科位置存疑(顾俊杰, 2012),但前缘域脉序基本符合鸣螽科特征,故建议使用异鸣螽亚科为中文名。
综上所述现将鸣螽科、各亚科及现生属种对应中文名罗列如下:
Hagloidea Handlirsch, 1906原螽总科
Prophalangopsidae Kirby, 1906鸣螽科
†Protaboilinae Gorochov, 1995原阿博鸣螽亚科
†Aboilinae Martynov, 1925阿博鸣螽亚科
†Chifengiinae Hong, 1982赤峰鸣螽亚科
†Termitidiinae Zeuner, 1939螱鸣螽亚科
†Tettohaglinae Gorochov, 2003异鸣螽亚科
Prophalangopsinae Kirby, 1906鸣螽亚科
ProphalangopsisWalker, 1871 鸣螽属
P.obscura(Walker, 1869) 暗鸣螽
TarragoilusGorochov, 2001 铁鸣螽属
T.diuturnusGorochov, 2001 长生铁鸣螽
AboilomimusGorochov, 2001亚鸣螽属
A.guizhouensisLiu, Zhou, Bi and Tang, 2009 贵州亚鸣螽
A.ornatusLiu, Zhou, Bi and Tang, 2009 丽亚鸣螽
A.sichuanensisGorochov, 2001 蜀亚鸣螽
Cyphoderrinae Gorochov, 1988圆翅鸣螽亚科
CyphoderrisUhler, 1864 圆翅鸣螽属
C.buckelliHebard, 1934 巴氏圆翅鸣螽
C.monstrosaUhler, 1864 怪圆翅鸣螽
C.strepitansMorris and Gwynne, 1978 噪圆翅鸣螽
ParacyphoderrisStorozhenko, 1980 拟圆翅鸣螽属
Pa.erebeusStorozhenko, 1980 穴拟圆翅鸣螽
2 鸣螽科形态特征与生物学
2.1 鸣螽科形态学特征
鸣螽体中到大型(图1),成虫体长(头至翅端)25 mm~120 mm;现生类群体色多为暗褐色,体型相对较小;灭绝类群体型多样,最大者翅展超过200 mm,最小体长不足30 mm。
头:下口式;触角丝状,着生于复眼下缘水平之上,柄节圆柱状、长于梗节,鞭节第一亚节通常长于其他鞭节亚节。眼:一般具3枚单眼;复眼卵形,通常较大,位于头前上方两侧。口器:唇基阔,上唇大;上颚形态多样,部分种类上颚粗短,切齿较钝,臼齿域发达,适于咀嚼植物组织;部分种类上颚延伸、细长,切齿尖锐,端齿长而尖、弯钩状,适于捕食,如在道虎沟生物群常见的巨大奇异阿博鸣螽Allaboilusgigantus,其上颚与现生典型的捕食性螽斯上颚类似(Rentz, 2010; 石福明等, 2013)。
胸:前胸背板发达,马鞍状,后缘通常盖住翅基部,侧叶通常发达。足:前中足基节具短刺(鸣螽亚科),捕食性种类前足胫节具粗壮刺;后足股节通常膨大,为典型的跳跃足;前足胫节基部内外侧各具1听器,椭圆形,多为开放型,偶见封闭型(图2);前中足胫节腹面多具刺,后足胫节背面两侧多具刺,胫节端部通常具距,部分捕食性种类前足胫节轻度膨大;前中后足跗节均4节,具跗垫,灭绝类群一般第一节较长,现生类群一般第4节细长,端部具一对爪,爪间具中垫;鸣螽亚科昆虫跗节第1第2节发生不同程度愈合(图3)。
翅:翅通常发达,前翅为覆翅,翅脉复杂(图4),灭绝类群前翅多具色斑,类型多样,后翅宽阔,臀区发达;前翅翅形以椭圆、卵圆形为主,一般超过腹部末端,部分现生类群翅趋向缩短,如长生铁鸣螽雌性通常翅退化,几乎被前胸背板覆盖。前翅亚前缘域较宽,ScA通常发达且横切亚前缘域;MA与RP无融合;RA、RP通常具栉状分支;雄性个体发声部较大,具发声锉与刮器,CuPb强烈弯曲,中部发育发声齿,左右翅发声器官一般同等发育;前翅色斑类型多样,以条带型为主,现生类群体色较暗,只有丽亚鸣螽翅面交错分布浅黄绿色色斑。张海春等(2015)将鸣螽翅色斑分为8种类型,并认为是鸣螽快速辐射并适应环境的结果。后翅膜质,臀区通常扩大呈扇形,铁鸣螽后翅较前翅短小,臀区无扩大。
发声器结构:包括灭绝类群在内的鸣螽发声器官分化明显,表现出了一些明显区别与螽斯总科昆虫的、较为原始的特征。从结构上看,发声锉位于CuPb脉中部,CuPb强烈弯曲,近Z字形,与相邻的CuPaβ形成一较宽阔区域,区域内布满网状或直横脉,偶见弯曲横脉,多数鸣螽种类雄性个体左右前翅刮器、腹面发声锉、发声齿同等发育且都具有发声功能,发声齿形态结构及排列方式十分多样(Guetal., 2012; 顾俊杰, 2012)。灭绝类群中除蚁鸣螽外,多数无明显的镜膜域,但CuPb与CuPa所围成的区域,以及部分种类中脉域扩增的区域很可能起到与镜膜相似的放大共鸣的作用。怪圆翅鸣螽有较明显的镜膜域,由CuPaα、handle、CuPaα和CuPaβ基半部围成的翅室形成(Chiversetal., 2017)。发声齿的数量、形态及排列方式在鸣螽科种间差异十分显著,不同的发声齿数目及排列决定了发声频率等声学特征的差异,可以在翅形态接近或特征不全的情况下辅助分类鉴定。Wang等(2018b)曾报道巨大奇异阿博鸣螽新材料,并描述了其发声器结构,但比较其发声结构与巨大奇异阿博鸣螽模式标本后发现,虽翅脉序形态特征接近,但发声齿形态、数量、排列方式差异明显,且翅长约为模式标本的一半,应为不同种。

图1 鸣螽标本照片 Fig.1 General habitus of Prophalangopsidae 注:1,穴拟圆翅鸣螽Pa. erebeus ♂;2,长生铁鸣螽T. diuturnus ♂;3,奇异曲阿博鸣螽Sigmaboilus peregrinus ♀;4,暗鸣螽P. obscura ♀;5,巨大奇异阿博鸣螽Al. gigantus ♀ (修改自Gu et al. 2010);比例尺=10 mm。Note:1,Pa. erebeus ♂; 2,T. diuturnus ♂; 3,Sigmaboilus peregrinus ♀; 4,P. obscura ♀; 5,Al. gigantus ♀ (modified from Gu et al. 2010); Scale bar=10 mm.
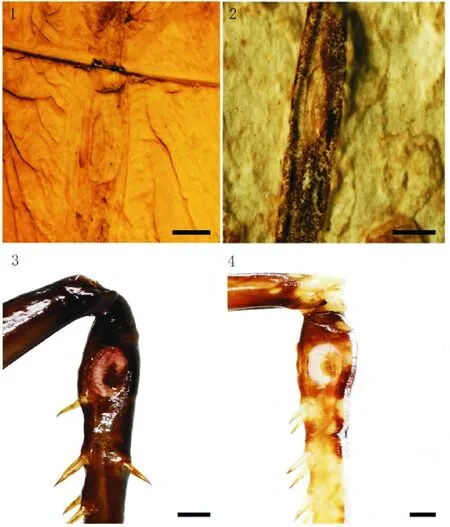
图2 鸣螽听器Fig.2 Auditory tympanum of Prophalangopsidae注:1,韩氏奇异阿博鸣螽Al. hani听器,♀;2,鸣螽未定种听器;3,长生铁鸣螽听器,♂;4,丽亚鸣螽听器,♂;比例尺=1 mm。Note:1,Tympanum of Al. hani, ♀; 2,Undetermined auditory organ of katydid; 3,Tympanum of T. diuturnus ♂; 4,Tympanum of A. ornatus ♂; Scale bar=1 mm.

图3 鸣螽跗节 Fig.3 Tarsus of Prophalangopsidae 注:1,长生铁鸣螽后足跗节;2,穴拟圆翅鸣螽后足跗节;3,丽亚鸣螽后足跗节;4,多叉新阿博鸣螽Novaboilus multifurcatus前足跗节;比例尺=2mm。Note:1,Hind tarsus of T. diuturnus; 2,Hind tarsus of Pa. erebeus; 3,Hind tarsus of A. ornatus; 4,Fore tarsus of Novaboilus multifurcatus; Scale bar=2 mm.
腹部:一般圆筒形,多数种类腹部第10节背板与肛上板融合。灭绝类群雌性产卵器发达且形态多样,多呈剑状、弯刀状等,部分种类背产卵瓣端部具齿(顾俊杰, 2012);现生类群产卵器退化。因保存关系,灭绝类群雄性生殖节结构了解较少,现生雄性肛侧板具一对钩突,下生殖板具成对针突,尾须较短。
2.2 生物学
鸣螽科现生类群分布十分狭窄。北美分布的圆翅鸣螽研究相对较深入,对其生物学、行为学有较多研究。中国西南地区及印度分布的鸣螽,种群数量较小,在物种被描述之后鲜有报道和研究。
2.2.1生殖
鸣螽科中关于圆翅鸣螽属生殖行为的研究较多。研究表明圆翅鸣螽的交配一般存在引诱和强制行为。在交配过程中,雄性腹部的阱铗(gin trap)可能起着重要的作用(Sakaluketal.,1995)。当雌性爬上雄性背部,噪圆翅鸣螽雄性会挤压腹部利用阱铗结构扣住雌性腹端,特别是当缺乏营养的雌性个体只欲进食雄性后翅而不交配时,阱铗可强制交配直至雌性接受精子,以保证交配成功率,圆翅鸣螽属其他种可能也存在相同的方式(Morris and Dodson, 1981; Dodsonetal., 1983; Eggert and Sakaluk, 1994; Sakaluketal., 1995)。对营养的需求可能是影响雌雄交配选择的主要因素之一,雄性会通过后翅诱使雌性交配,但贡献后翅对雄性影响较大,雌性通常优先选择与未交配雄性进行交配,当雄性后翅完整时,交配具更高成功率(Morris and Gwynne, 1979; Morris and Dodson, 1981; Morrisetal., 1989; Johnsonetal., 1999)。在交配季节末期,部分雄性个体后翅全部被咀嚼掉,为保存交配竞争力,雌雄单次交配中后续阶段表现为更高水平的斗争,巴氏圆翅鸣螽雌性会使用后足跗节推雄性的腹端,阻止生殖器靠拢,交配开始后雄性会阻止雌性继续取食后翅,推动雌性远离它的后翅外缘(Judge and Morris, 2011)。当精包转移后,雄性立即释放雌性,结束交配,雌性通常此后不再交配(Dodsonetal., 1983)。Judge和Morris(2011)发现经饥饿处理的雌性巴氏圆翅鸣螽比食物充足的雌性更有可能取食雄性后翅,也更有可能发生交配。而Johnson等(1999)对野外噪圆翅鸣螽未交配雄性后翅进行破坏处理,发现并未明显影响雄性成功进行交配。此外,后翅的损伤和精囊转移,会显著降低雄性个体能量,难以维持其高耗能的鸣声活动,后翅完整的雄性噪圆翅鸣螽常拥有更高交配率,且鸣叫声持续时间更长(Sakaluketal., 1987; Morrisetal., 1989; Sakaluk and Snedden, 1990),当雄性失去发声能力时,交配率明显下降(Snedden and Sakaluk, 1992)。实验室中处于饥饿状态的雄性仍能产生精包(Eggert and Sakaluk, 1994a; Johnsonetal., 1999),表明精包转移可能并不是影响交配的主要因素。除此之外,还有多种因素会影响交配行为及成功率,如雌性一般会选择鸣声更为响亮,即能提供更多的能量的雄性,但体型和鸣叫的关系仍存在争议(Mason, 1996)。Sakaluk等(1990)认为体型较大的噪圆翅鸣螽可能具有较高能量且保持较长时间鸣叫从而提升交配率,但后续实验认为它们之间可能并无关联(Sakaluketal., 1995)。在交配过程中雄性血淋巴的消耗会影响其下次交配(Sakaluketal., 2003)。此外,后翅受损导致免疫系统上调也可能会影响交配率(Lemanetal., 2009)。
螽斯产卵方式为隐蔽式,产卵地点选择与其产卵器形态,卵形态直接关联,多数螽斯产卵于土壤中,部分类群产卵于植物嫩茎、枝干缝隙、叶片或腐木中(Rentz, 2010)。鸣螽科现生类群产卵器退化,如圆翅鸣螽有可能在土壤中产卵,并且有看护、孵化行为(Gwynne, 2001)。拟圆翅鸣螽很可能产卵于土壤洞穴中。中生代鸣螽很可能产卵与土壤中(赤峰鸣螽亚科),或植物枝干、裂隙(阿博鸣螽亚科部分种类)、或植物组织中(曲鸣螽属Sigmaboilus)(顾俊杰, 2012)。
2.2.2声通讯
鸣螽科发声器官结构及行为较为独特,与其亲缘关系较近的螽斯总科差别较大。从结构上看,不论现生类群还是灭绝类群,均表现出左右翅对称的结构,即左右前翅发声锉、发声齿近同等发育。现生鸣螽如圆翅鸣螽、铁鸣螽均可通过前翅摩擦发出类似蟋蟀的乐音(律音)的鸣叫声,且前翅变换左右叠覆状态仍可摩擦发声(Morrisetal., 2002)。圆翅鸣螽鸣叫高峰时段为5-8月的夜晚,长生铁鸣螽为7-8月的晚7-10点。圆翅鸣螽的鸣声频率为12~15 kHz,高于蟋蟀(3~8 kHz)(Spooner, 1973; Morris and Gwynne, 1978; Otte, 1992; Morrisetal., 2002)。Chivers等(2017)对圆翅鸣螽前翅发声器功能形态进行了研究,认为圆翅鸣螽发声器结构功能与现代螽斯不同,处于向特化的其他现生螽斯的过渡阶段。侏罗纪时期原螽科的悦耳古鸣螽Arcaboilusmusicus被发现同样可以发出低频的、乐音的鸣叫声,频率大约为6.4 kHz(Guetal., 2012),此外,中生代鸣螽很可能在通讯行为及鸣声物理特征上均发生了明显的分化(Guetal., prepare)。研究表明巴氏圆翅鸣螽和怪圆翅鸣螽的雌虫可以通过雄虫鸣叫声中的高频衰减来区分雄虫的大小,并利用鸣声强度来判断雄虫的距离,而雄虫通过载波频率和脉冲强度来确定竞争优势(Morrisetal., 2002)。圆翅鸣螽不同种通讯行为有所差异,领地意识较强的怪圆翅鸣螽在鸣叫时会升级为物理冲突,巴氏圆翅鸣螽和噪圆翅鸣螽则持续通过声学竞争,野外巴氏圆翅鸣螽的鸣声也具明显聚集作用(Morrisetal., 2002)。怪圆翅鸣螽通讯行为会对外界温度变化做出反应(Morris and Gwynne, 1978),同时其听觉敏感与鸣声载频明显错配,对2 kHz的音频最敏感,但其鸣声主频在12 kHz附近,可能与躲避捕食者相关(Mason, 1991; Masonetal., 1999)。
2.2.3食性
古直翅目昆虫在石炭纪时期食性已经发生分化,目前已经发现植食性和肉食性的代表类群(Guetal., 2011; 顾俊杰, 2012)。现生直翅目昆虫多为植食性,也存在一些杂食性和典型的捕食性代表。鸣螽科灭绝类群食性也发生了分化。Krassilov等(1997)首次在鸣螽科化石Aboilusamplus和Aboilusdilutus肠道中发现有机物内含物的克拉梭粉属Classopollis的花粉粒。此外也有典型的捕食性种类,如巨大奇异阿博鸣螽,具发达的上颚,端部延伸具尖齿,与现生直翅目典型捕食性上颚相似(顾俊杰, 2012)。而曲阿博鸣螽Sigmaboilusspp.,阿博鸣螽属Aboilus的多数种类上颚短钝、臼齿域发达,很可能为植食性。现生鸣螽的食性报道较少,对圆翅鸣螽的研究和野外观察表明,圆翅鸣螽野外多以裸子植物及被子植物花粉、植物叶片以及虫瘿为食(Morris and Gwynne, 1978, 1979)。鸣螽亚科食性尚未见正式报道。
3 系统分类与进化
3.1 鸣螽科系统分类
鸣螽科模式种暗鸣螽发现于印度东北部,最早被命名为TarragaobscuraWalker, 1869,但因占用了Polyphagidae中1属学名而无效,之后重新定名ProphalangopsisobscuraWalker 1971(Walker 1869-1871)。Kirby(1906)据此建立新亚科鸣螽亚科,并归入Phasgonuridae (Tettigoniidae),但未有详细记述。Caudell(1911)首次对该亚科进行了描述。Handlirsch(1925)将其归入沙螽科Stenopelmatidae,随后提升为科,建立新科鸣螽科Prophalangopsidae。随后Karny和Wytsman(1937)又将其转移到蟋螽科Gryllacrididae,但未被认可。Zeuner(1939)将Haglinae、 Prophalangopsinae、 Cyrtophyllitinae、 Geinitziinae归入鸣螽科。Sharov(1968)认为科名Haglidae较Prophalangopsidae优先,应包括Haglinae、Prophalangopsinae、Cyphoderrinae、Cyrtophyllitinae、Isfaropterinae等亚科。鸣螽属以及圆翅鸣螽属长期以来被认为是鸣螽科的现生代表(Zeuner, 1939; Ragge, 1955; Vickery and Kevan, 1983)。但Otte(1997)认为圆翅鸣螽属及拟圆翅鸣螽属应归入原螽科,鸣螽属应归入鸣螽科。Gorochov(1995)对螽亚目昆虫进行了系统回顾,认为原螽总科应包括鸣螽科、原螽科、Tuphellidae和Hagloedischiidae;其中鸣螽科以发达的ScA脉横切亚前缘域、RP与MA无融合或小脉相连为特征,由4灭绝亚科及鸣螽亚科和圆翅鸣螽亚科2现生亚科组成。此后,Gorochov(2003)对鸣螽科的进化、生物学、组成及各亚科特征进行了阐述,并建立鸣螽科新亚科—异鸣螽亚科。但异鸣螽亚科只有1属1种,且基于1块雄性标本建立。依据其前翅线条图、前缘域及亚前缘域呈现了典型的鸣螽特征,但径脉后支至肘脉区域与鸣螽,乃至原螽总科差异巨大,不排除端半部存在化石形变或翅折叠、或者个体变异等因素,因此需要检视模式标本才能做出进一步判断该亚科是否成立。目前为止,现生鸣螽总计发现 5属 9种,分属于两个亚科。Morris(1978)描述了北美分布的圆翅鸣螽第3种,噪圆翅鸣螽。Storozhenko(1980)报道了发现于西伯利亚东部地区的穴拟圆翅鸣螽,这是该亚科首次在亚洲大陆发现。Gorochov(2001a, 2001b)先后发表了发现于中国四川的现生鸣螽新种:蜀亚鸣螽和长生铁鸣螽。Liu等(2009)描述了发现于贵州赤水和四川天全的两个新种:贵州亚鸣螽和丽亚鸣螽,并在西藏墨脱首次发现了之前只在印度有记录的暗鸣螽。直翅目物种库网站(http://orthoptera. speciesfile.org/)将贵州亚鸣螽和丽亚鸣螽视为蜀亚鸣螽的亚种,但缺乏详细数据和引文(Ciglianoetal., 2020)。
相对于物种多样性较低的现生类群,化石鸣螽曾广泛分布于各大陆块的早侏罗世至早白垩世地层中,目前直翅目物种库网站共收录95种(Ciglianoetal., 2020)。阿博鸣螽亚科在鸣螽科中多样性最高,但其单系性存疑(Béthoux and Nel, 2002; 顾俊杰, 2012)。赤峰鸣螽亚科以退化的ScA脉、MA和MP区域基部窄,后明显增宽为典型特征,目前共描述4属8种。原阿博鸣螽亚科模式种为ProtaboiluspraedictusGorochov, 1988,任东和孟祥明(2006)曾发表5新种并归入该亚科,但随后被认为应归入阿博鸣螽亚科,其中Pr.liniRen and Meng, 2006被认为是层状阿博鸣螽Ab.stratosus的同物异名(Guetal., 2010)。Gorochov(2003)认为原阿博鸣螽亚科是鸣螽科最原始的类群,其CuPb-CuPaβ间横脉弯曲区别于其他所有鸣螽科类群。但原阿博鸣螽亚科的单系性在支序系统学分析中并未得到确认(Béthoux and Nel, 2002)。蚁鸣螽亚科目前发表6属10种,其中1属2种存疑。除了宁城拟阿博鸣螽Pseudaboilusningchengensis发现于中国外(Wangetal., 2018a),其余各种均发现于英国普尔拜克地区(Purbeck)侏罗纪至白垩纪地层。该亚科翅形窄长,鸣器区域相对较小,亚前缘域较窄,前翅臀区向后缘方向扩大等。异鸣螽亚科目前仅发现1属1种,且位置存疑。Gu等(2010)对中国发现的鸣螽进行了回顾,发现新异名9属12种并分析了鸣螽种内前翅脉序形态的差异。中华木螽Hylophalangopsischinensis发现于西藏羌塘盆地古新世牛宝组(Lin and Huang, 2006),被认为属于鸣螽科,且认为模式标本为发声器特化的雄性前翅。但Wang等(2019)认为中华木螽纵脉近平行,CuPb与AA1很可能到达翅的端半部,因此应该归属与沙螽科。但通过化石标本照片可以看出模式标本后缘很可能存在挤压变形的情况,肘脉与臀脉的走向很可能受到挤压变形,因此将其转移至沙螽科的证据并不充分。
3.2 系统发育与进化
关于鸣螽科的系统位置尚未有统一的认识。多数学者都认为鸣螽科与螽斯科亲缘关系较近,或为姐妹群关系(Ander, 1939; Zeuner, 1939; Ragge, 1955; Gwynne, 1995; Songetal., 2020)。Gorochov(2003)基于翅形态特征分析认为鸣螽科是螽斯总科和沙螽总科Stenopelmatoidea的祖先类群。Jost和Shaw(2006)认为鸣螽科与螽亚目其他类群为姐妹群关系。Desutter-Grandcolas(2003)基于外部形态、翅形态、消化道、呼吸、神经系统结构等特征的支序系统学研究认为鸣螽科(鸣螽属、圆翅鸣螽属)可能与螽斯科及丑螽科Anostostomatidae亲缘关系较近。Song等(2015) 基于线粒体全基因组和4个核基因分子标记的系统发育研究认为,原螽总科(圆翅鸣螽属+铁鸣螽属)与沙螽总科互为姐妹群关系,但该研究并未涉及灭绝类群。研究通过化石校准,对螽亚目各类群分歧时间进行了估算,但并未选择中生代分布最为广泛的鸣螽科化石代表,且在估算时使用了错误的地质年代表,导致对各类群推测的分歧时间偏晚。Zhou等(2017)基于线粒体基因组数据的分析结果支持原螽总科与沙螽总科的姐妹群关系。基于翅形态特征的,以灭绝类群为研究对象的系统发育分析表明,鸣螽科的单系性可以确认,但原螽总科及原螽科单系性均未得到确认,其关系尚不明确(Béthoux and Nel, 2002)。Kumala等(2005)基于外部形态和行为特征对圆翅鸣螽亚科进行了系统发育分析。综上所述,鸣螽科、原螽总科以及原螽总科与螽亚目的系统发育关系还需进一步研究,特别是整合形态、分子、现生、化石数据的系统发育研究尤为必要。
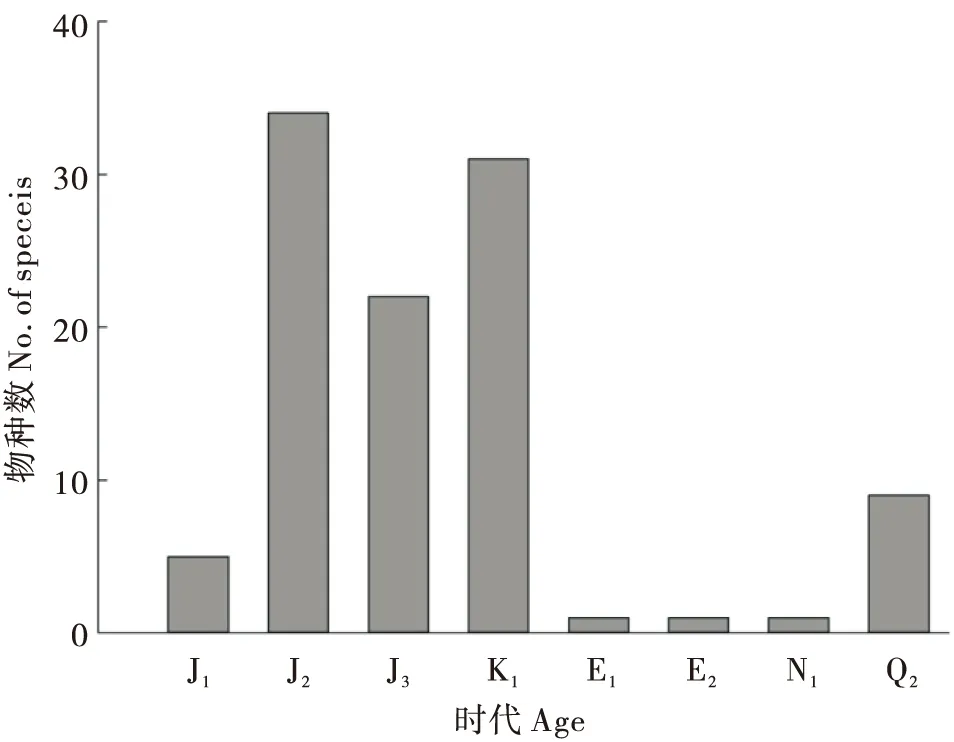
图5 各地质历史时期鸣螽科物种数Fig.5 The species number of Prophalangopsidae in geological time
4 地质历史与分布
原螽科昆虫在中三叠世就已经出现。最早的鸣螽发现于中亚地区和我国早侏罗世地层中,在中侏罗世鸣螽辐射扩散广布于各大陆(图5),达到多样性高峰。晚侏罗世鸣螽科昆虫的分异度在全球范围内明显下降。到早白垩世,分异度又有所回升,并达到该科演化过程中的又一次高峰(张海春等, 2015)。此后鸣螽科分异度急剧下降,未见化石记录。张海春等(2015)认为鸣螽科昆虫的衰落可能从晚白垩世早期就已经开始了,不太可能是因白垩纪末的生物大灭绝事件而引起的,很可能与银杏类植物的迅速衰落有关。新生代时期化石记录较少,仅在加拿大阿尔伯特及中国西藏羌塘古新世地层中有零星发现(Kevan, 1981; Lin and Huang, 2006),且分类位置存在争议(Wangetal., 2019)。
现生鸣螽分布地十分狭窄,目前仅在北美地区西北部、西伯利亚东部(圆翅鸣螽亚科)以及印度北部,西藏、四川、贵州(鸣螽亚科)有记录。圆翅鸣螽属分布在北美海拔1 500~3 100 m的高山,其生境以裸子植物为主。铁鸣螽属目前仅在四川石棉栗子坪保护区有记录。亚鸣螽分布相对较广,在贵州赤水、四川天全和映秀均有记录,一般分布在海拔900~2 600 m,湿度较大,温度较低的近水环境中,多在地表或灌木上活动。结合鸣螽现有分布格局及生境特点,鸣螽在我国西南地区及秦岭地区可能存在更多潜在的分布区。
5 问题与展望
鸣螽作为螽亚目目前已知唯一的从早侏罗世延续到现在的科,对理解螽亚目系统发育、演化辐射及行为演化均有重要的研究意义。由于雌雄二型等因素,目前鸣螽科灭绝类群多数基于雄性前翅标本建立,也有一部分仅仅基于单一雌性前翅标本建立,只有少数几种是基于雌雄标本建立或者发现了同一种的雌雄个体(Guetal., 2010)。化石鸣螽往往结构保存不完整,且标本相对稀缺,在分类过程中往往容易产生对一些如翅脉等形态特征在种内变异认识的不足。因此,现有的已发表鸣螽科化石种级单元的有效性、有多少潜在同物异名、性二型在分类工作中的认识等是值得关注的问题。此外,目前昆虫学者在研究螽亚目系统发育关系时很少考虑灭绝类群,只有鸣螽科的个别现生类群偶尔出现在他们的研究对象中,除了化石标本形态信息缺失较多的原因之外,翅脉形态在系统发育关系研究中的作用长期以来被学者们忽视,由于大多数化石标本只保留前翅,因此翅脉是联系化石类群和现生类群的唯一的、也是最有效的“纽带”。翅在漫长的演化历史过程中其功能以及形态上发生了巨大的变化,虽然翅形态存在一定平行、逆转现象,但同时也保留了昆虫大量的演化和亲缘关系的信息,对系统发育研究具有重要的意义。因此有必要明确翅脉形态在螽亚目昆虫中的分类学价值,建立起化石与现生螽斯昆虫分类之间的联系。
鸣螽的生物学还知之甚少,只有圆翅鸣螽属得到了较多的室内和野外研究。鸣螽亚科及拟圆翅鸣螽的生活史、食性、生殖行为、通讯行为、生态习性、生境和分布等问题还未得到充分研究。笔者经过多年的野外考察认为四川西南及北部山区很可能是鸣螽亚科的分布及演化中心,但其对生境要求非常高,特别是对温度湿度的要求,且迁移能力较弱,分布区域狭窄且不连续。此外,经历了多次地震、泥石流、洪水等自然灾害的影响,如长生铁鸣螽原有的生境已经遭到部分破坏,野外种群分布已发生了一定变化。因此,当前应加大研究力度,明确其生活史、生殖行为等生物学问题,提出相应的保护措施。
致谢:感谢审稿专家提出的宝贵意见。感谢首都师范大学田河博士、中国科学院上海昆虫博物馆朱卫兵博士、王瀚强博士协助拍摄标本照片,感谢河北大学石福明教授,俄罗斯科学院远东分院Storozhenko S.Yu.博士在文献和图片资料方面提供帮助。
猜你喜欢
农业科技与信息(2020年14期)2020-12-18
武夷科学(2019年2期)2019-12-20
绿色科技(2019年14期)2019-11-19
环球时报(2019-10-24)2019-10-24
江苏农业科学(2019年5期)2019-09-02
绿色科技(2018年19期)2018-11-29
