
番茄潜叶蛾卵黄原蛋白受体基因TaVgR在生殖发育调控中的作用
2022-07-13 06:55宋佳慧李仁贵张毅波杨文佳
昆虫学报 2022年6期
闫 欣, 杨 洪, 宋佳慧, 李仁贵, 张毅波, 杨文佳,,*
(1. 贵州大学昆虫研究所, 贵州省山地农业病虫害重点实验室, 贵阳 550025; 2. 贵阳学院生物与环境工程学院,贵州省山地珍稀动物与经济昆虫重点实验室, 贵阳 550005; 3. 中国农业科学院植物保护研究所,植物病虫害生物学国家重点实验室, 北京100193; 4. 贵州大学烟草学院, 贵阳 550025)
昆虫卵黄原蛋白受体(vitellogenin receptor, VgR)属于低密度脂蛋白受体(low density lipoprotein receptor, LDLR)家族,其通过介导内吞作用把卵黄原蛋白(vitellogenin, Vg)转运至卵母细胞的膜结合小泡上,为胚胎发育提供营养物质,在昆虫卵巢发育过程中发挥重要作用(Tufail and Takeda, 2008; 陈利鹏等, 2016)。LDLR家族蛋白具有5个经典保守结构域,包括配体结合域(ligand binding domains, LBD)、表皮生长因子前体同源域(epidermal growth factor precursor homology domain, EGF)、O-联糖功能域(O-linked sugar domain, OLSD)、胞质尾域(cytoplasmic domain, CPD)和跨膜域(transmembrane domain, TMD)(Tufail and Takeda, 2008, 2009)。研究发现,黑腹果蝇DrosophilamelanogasterVgR发生突变后会引起卵母细胞无法累积卵黄多肽,从而导致雌性成虫不育(Schonbaumetal., 1995);家蚕BombyxmoriVgR突变可引起Vg不能顺利进入卵巢,导致卵体积缩小且颜色失常(Shuetal., 2011);通过RNAi技术沉默棉铃虫Helicoverpaarmigera、桔小实蝇Bactroceradorsalis、褐飞虱Nilaparvatalugens等昆虫的VgR后,会引起Vg在血淋巴中大量沉积、卵巢发育受阻和雌性不育等现象(Congetal., 2015; Luetal., 2015; Zhangetal., 2016)。上述研究结果表明,VgR在昆虫卵黄生成和卵母细胞发育过程中起着关键作用。因此,开展昆虫VgR的功能研究有助于深入了解该基因调控昆虫卵巢发育的分子机理,对削弱害虫或提升益虫的繁殖力均具有重要的实际意义(Roth and Khalaila, 2012)。
番茄潜叶蛾Tutaabsoluta隶属于鳞翅目(Lepidoptera)麦蛾科(Gelechiidae),又名番茄潜麦蛾,是一种极具毁灭性的世界检疫性害虫,主要危害番茄、茄子、马铃薯等茄科植物,对番茄表现出明显偏好性。番茄潜叶蛾雌成虫繁殖力强,单头产卵量最多可达260粒左右,主要散产于植物叶片、嫩茎、顶芽和果实上,幼虫一经孵化便钻蛀植物组织潜食危害,造成叶片皱缩枯萎、顶芽枯死、未熟果实腐烂或脱落,危害严重时可导致番茄产量损失高达80%~100%(Desneuxetal., 2010; Zhangetal., 2020)。番茄潜叶蛾原产于南美洲的秘鲁(Biondietal., 2018),2006年入侵西班牙后迅速扩散至整个非洲-欧亚大陆(Zhangetal., 2020)。2017年8月,张桂芬等于新疆维吾尔自治区伊犁哈萨克自治州发现了番茄潜叶蛾,证实该害虫已入侵我国,推测其会对我国的番茄生产造成巨大危害(Zhangetal., 2020)。番茄潜叶蛾具有气候环境适应性强、定殖速度快的特性,其暴发危害会大幅增加化学农药使用量,导致番茄市场价格升高等一系列不利市场效应(Zappalàetal., 2013)。本研究在番茄潜叶蛾转录组数据的基础上,克隆卵黄原蛋白受体基因TaVgR,解析其在番茄潜叶蛾不同发育阶段和成虫不同组织中的表达模式,并采用RNAi技术解析其在番茄潜叶蛾生殖发育过程中的调控作用。研究结果不仅有助于阐明昆虫生殖调控机制,也可为番茄潜叶蛾的绿色可持续治理提供新思路和新靶标。
1 材料与方法
1.1 供试虫源
供试番茄潜叶蛾种群于2018年采自云南省玉溪市,在实验室阳光房中连续饲养至今。番茄潜叶蛾幼虫以新鲜番茄幼苗为食,成虫供以10%的蜂蜜水。 饲养条件为温度26±1℃,相对湿度60%±5%,光周期16L∶8D。
1.2 主要试剂
总RNA提取试剂TransZolUp、基因组DNA去除及反转录试剂盒TransScript®One-Step gDNA Removal and cDNA Synthesis SuperMix、PCR试剂盒TransTaq®DNA Polymerase High Fidelity、大肠杆菌Escherichiacoli感受态细胞Trans5α、qPCR试剂盒TransStart®Top Green qPCR SuperMix (北京全式金生物公司);胶回收试剂盒HiPure Gel Pure DNA Kits(Magen);载体pGEM-T Easy Vector (Promega);RNAi试剂盒TranscriptAid T7 High Yield Transcription Kit (Thermo Scientific)。
1.3 总RNA提取与cDNA第1链的合成
按照TransZolUp试剂盒说明书提取羽化后第3天番茄潜叶蛾雌成虫总RNA,使用NanoDrop 2000核酸蛋白浓度测定仪(Thermo Scientific)检测所提RNA的浓度和OD260/OD280值(1.8~2.2之间),通过1%琼脂糖凝胶电泳检测总RNA的完整性。以1 μL优质RNA样品为模板,按照TransScript®One-Step gDNA Removal and cDNA Synthesis SuperMix说明书合成cDNA第1链,存于-20℃冰箱备用。
1.4 番茄潜叶蛾TaVgR基因的克隆
基于番茄潜叶蛾转录组获得的VgR基因的unigene序列,使用Primer Primier 6.0软件设计特异性引物(表1),进行RT-PCR,扩增TaVgR基因的开放阅读框序列。PCR反应体系:TransTaq®HiFi DNA Polymerase 1 μL, 10×TransTaq®HiFi Buffer Ⅱ 5 μL, dNTPs (2.5 mmol/L) 4 μL, cDNA模板2 μL, 上下游引物(10 μmol/L)各1 μL, ddH2O补至总体积50 μL。反应程序: 94℃预变性5 min; 94℃变性30 s, 55℃退火30 s, 72℃延伸5 min, 共35个循环;最终72℃延伸10 min。PCR产物通过1%琼脂糖凝胶电泳分离鉴定,按照HiPure Gel Pure DNA Kit说明书回收目的条带,连接至pGEM-T Easy Vector进行TA克隆,经蓝白斑筛选挑取单个白色菌落扩大培养,PCR鉴定出阳性克隆后,将其送至成都擎科梓熙生物技术有限公司进行测序。

表1 引物信息Table 1 Primer information
1.5 序列分析
采用DNAMAN 6.0 (Lynnon Biosoft)对TaVgR基因序列进行编辑和分析;利用BLAST工具(http:∥www.ncbi.nlm.gov/BLAST/)对推导的氨基酸进行同源性比对分析;利用SignalP 6.0 Server (http:∥services.healthtech.dtu.dk/services.php?SignalP)预测信号肽;利用ProtParam(http:∥web.expasy.org/)分析理化性质;利用NetNGlys 1.0 server (http:∥www.cbs.dtu.dk/services/NetNGlyc/)预测N-糖基化位点;利用NCBI在线工具conserved domains (https:∥www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)查找保守区域;利用TMHMM Server 2.0 (http:∥www.cbs.dtu.dk/services/TMHMM)分析跨膜域;使用MEGA 6.0软件中的邻接法(neighbor-joining)构建系统进化树,树中各分支均进行1 000次的重复检验(Tamuraetal., 2013)。
1.6 番茄潜叶蛾TaVgR基因的时空表达模式测定
选取同一批孵化的番茄潜叶蛾幼虫,以长势一致的番茄苗为寄主植物进行饲养,分别收集1-4龄幼虫、1-7日龄雌蛹和雌成虫样品,每个发育时期30~50头试虫为一生物学重复,设置3个生物学重复。在雌成虫羽化第5天,使用微量CO2麻醉后进行解剖,分别获得头、体壁、前肠、中肠、后肠、卵巢、脂肪体和马氏管共8种组织样品,每组织解剖50头试虫为一生物学重复,设置3个生物学重复。将所有样品经液氮速冻后保存于-80℃冰箱,按照1.3节方法分别提取上述材料总RNA并合成cDNA。根据本实验室前期对番茄潜叶蛾内参基因稳定性的评估结果,选取TaEF1a(GenBank登录号: MZ054826)作为内参基因(Yanetal., 2021)。qPCR扩增在CFX96TMReal-Time System实时定量PCR仪(Bio-Rad)中进行,反应体系(20 μL):TransStart®Top Green qPCR SuperMix 10 μL, ddH2O 7 μL, cDNA模板1 μL, 上下游引物(10 μmol/L)各1 μL。qPCR反应程序: 95℃预变性5 min; 95℃变性15 s, 60℃退火30 s, 72℃延伸30 s, 共40个循环;最后于60~95℃进行熔解曲线分析。每样品重复测定3次。采用2-ΔΔCT方法计算TaVgR在番茄潜叶蛾不同发育阶段和雌成虫不同组织中的相对表达量(Livak and Schmittgen, 2001)。
1.7 番茄潜叶蛾TaVgR的RNAi实验
根据番茄潜叶蛾TaVgR和绿色荧光蛋白基因GFP的cDNA序列,在其5′端添加T7启动子序列,设计双链合成引物(表1),参照TranscriptAid T7 High Yield Transcription Kit说明书合成dsRNA。合成的dsRNA经纯化后,用RNAi注射缓冲液稀释至1 μg/μL备用。随机选取大小一致、发育整齐的番茄潜叶蛾4日龄雌蛹,利用Nanoliter 2010全自动显微注射仪(World Precision Instruments)进行dsRNA注射,每头试虫的注射量为500 ng,同时注射等量的dsGFP作为阴性对照,设置3个生物学重复,每个重复注射30头。注射后的蛹放入无菌土壤中正常饲养,分别收集dsRNA注射12, 24, 48和72 h后的试虫,采用RT-qPCR技术(方法同1.6节)检测RNAi对靶标基因的沉默效率。dsRNA注射24 h后观察蛹的存活情况并记录,待羽化并与同天羽化的雄蛾以1∶3的比例放入单独饲养装置(含新鲜番茄叶片)中,雌、雄成虫正常交配并产卵,每24 h更换一次新叶片,计算单雌1-10 d每日产卵量和10 d总产卵量,以及后代卵的孵化率。选取羽化后第1天的雌成虫进行解剖,观察卵巢发育情况,测量卵巢管长度和卵粒长度,并用VHX-2000超景深三维显微观察系统(基恩士)拍照记录。
1.8 数据分析
实验结果采用平均值±标准误表示,基因表达量数据采用SPSS 23.0软件的单因素方差分析法进行差异显著性分析,RNAi后处理组和对照组之间进行差异显著性分析采用独立样本t检验进行分析。利用GraphPad Prism 8.0.1软件制图。
2 结果
2.1 TaVgR基因的克隆与序列特征
利用RT-PCR扩增获得番茄潜叶蛾TaVgR基因(GenBank登录号: MZ682118)的cDNA全长序列,扩增产物经1%琼脂糖凝胶电泳检测后得到与预期条带大小一致的目的片段。TaVgR开放阅读框长5 496 bp,编码1 831个氨基酸,理论蛋白质分子量为206 kD,等电点为5.17。TaVgR编码的蛋白N端前18个氨基酸残基MAYQHLFFLFLCLMSCSA为信号肽,有16个糖基化位点(NXT和NXS)。跨膜域分析结果表明,第1 699-1 721位氨基酸为跨膜域,第1 722-1 831位氨基酸为胞质尾域,其余氨基酸部分为胞外区域。序列比对结果显示,番茄潜叶蛾VgR与其他22种昆虫VgR的结构高度相似,其保守区域及保守基序(LBD和EGF)位置较为一致,表明TaVgR是LDLR家族成员,具有5个典型的功能域(LBD, EGF, OLSD, CPD和TMD)。同源性序列比对发现,番茄潜叶蛾VgR与蓖麻蚕Samiaricini的VgR(GenBank登录号: AYA29178)和绿尾大蚕蛾Actiasselene的VgR (GenBank登录号: AFV32171)的氨基酸序列一致性均为47%,与甜菜夜蛾Spodopteraexigua的VgR (GenBank登录号: AOX13593)、棉铃虫Helicoverpaarmigera的VgR (GenBank登录号: AGF33811)和柞蚕Antheraeapernyi的VgR (GenBank登录号: AEJ88360)氨基酸序列一致性均为46%。
2.2 TaVgR的系统发育
系统发育分析结果显示,不同目的昆虫VgR各自聚成小类群,表明昆虫VgR在进化上具有较好的保守性;番茄潜叶蛾VgR和鳞翅目昆虫VgR聚为一支,且与螟蛾昆虫柑橘卷叶蛾Amyeloistransitella和粉斑螟蛾Cadracautella最为近缘(图1)。

图1 邻接法构建的基于氨基酸序列的番茄潜叶蛾与其他昆虫VgR的系统发育树(1 000次重复)Fig. 1 Phylogenetic tree of VgRs from Tuta absoluta and other insects constructed by neighbor joining method (1 000 repetitions)based on amino acid sequence蛋白来源物种Origin species of proteins: Ad: 排蜂Apis dorsata; Am: 西方蜜蜂Apis mellifera; Em: 兰花蜜蜂Eufriesea mexicana; Hl: 东南蓝莓蜂Habropoda laboriosa; Mr: 苜蓿切叶蜂Megachile rotundata; Ap: 豌豆长管蚜Acyrthosiphon pisum; Bt: 烟粉虱Bemisia tabaci; Nl: 褐飞虱Nilaparvata lugens; Nc: 黑尾叶蝉Nephotettix cincticeps; Aa: 埃及伊蚊Aedes aegypti; Dm: 黑腹果蝇Drosophila melanogaster; Md: 家蝇Musca domestica; Bd: 桔小实蝇Bactrocera dorsalis; Sr: 蓖麻蚕Samia ricini; Se: 甜菜夜蛾Spodoptera exigua; Sl: 斜纹夜蛾Spodoptera litura; Ha: 棉铃虫Helicoverpa armigera; Bm: 家蚕Bombyx mori; As: 绿尾大蚕蛾Actias selene; At: 柑橘卷叶蛾Amyelois transitella; Cc: 粉斑螟蛾Cadra cautella; Mv: 豆荚螟Maruca vitrata.
2.3 TaVgR基因的时空表达模式
番茄潜叶蛾不同发育阶段和雌成虫组织中TaVgR基因的表达模式解析结果(图2)显示,从幼虫期至蛹期,TaVgR基因的转录水平随着龄期的增加而升高,在雌蛹和雌成虫中的表达水平显著上升(P<0.05),且在雌成虫中达到峰值,较雌蛹期的TaVgR转录水平提高了4.57倍(图2: A)。TaVgR在番茄潜叶蛾雌成虫的卵巢中特异性高表达,表达水平显著高于其他组织中的(P<0.05),在其余组织中的表达水平则无显著差异(P>0.05)(图2: B)。
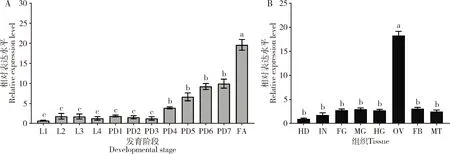
图2 番茄潜叶蛾TaVgR在不同发育阶段(A)和雌成虫组织(B)中的相对表达量Fig. 2 Relative expression levels of TaVgR in different developmental stages (A) and female adult tissues (B) of Tuta absolutaL1-4: 分别为1-4龄幼虫1st-4th instar larvae, respectively; PD1-7: 分别为1-7日龄雌蛹1-7-day-old female pupae, respectively; FA: 雌成虫Female adult; HD: 头Head; IN: 体壁 Integument; FG: 前肠Foregut; MG: 中肠Midgut; HG: 后肠Hindgut; OV: 卵巢Ovary; FB: 脂肪体Fat body; MT: 马氏管Malpighian tubules. 图中数据为平均值±标准误;柱上不同字母表示基因表达量在不同发育阶段和成虫不同组织间存在显著差异(P<0.05, LSD)。Data in the figure are mean±SE. Different letters above bars represent significant difference in the gene expression level between different developmental stages and adult tissues (P<0.05, LSD).
2.4 干扰TaVgR基因表达的生物学效应
利用RNAi技术沉默番茄潜叶蛾TaVgR基因,发现TaVgR的mRNA表达量在dsTaVgR注射后12, 24, 48和72 h较对照(注射dsGFP)分别下降了62.04%, 72.55%, 70.02%和70.15%(P<0.01),表明该基因的表达被有效抑制(图3: A)。待试虫羽化后按照雌雄比1∶3进行单独饲养,统计单头雌成虫1-10 d的产卵量。结果发现,对照组和处理组雌成虫每日产卵量均呈现先升高后降低的变化趋势,羽化第2天为产卵高峰期,此时处理组产卵量较对照组降低了61.71%(P<0.001)(图3: B)。测定注射后单雌1-10 d单日产卵量、10 d总产卵量及后代卵孵化率,发现注射dsTaVgR的番茄潜叶蛾的10 d单雌总产卵量为90粒,而注射dsGFP组的10 d单雌总产卵量可达到255粒,二者差异极显著(P<0.01)(图3: C),但二者的后代卵孵化率无显著差异(P>0.05)(图3: D)。
由于番茄潜叶蛾羽化后24 h内卵巢发育已经成熟,解剖此时的雌成虫卵巢,测量并记录卵巢管和成熟卵粒长度,发现注射dsTaVgR组平均卵巢管长度为2 846.11 μm,较注射dsGFP组(4 633.49 μm)下降了38.58%(P<0.05)(图3: E)。注射dsGFP组番茄潜叶蛾成熟卵粒较多,平均卵粒长355.43 μm,而注射dsTaVgR组成熟卵粒较少,平均卵粒长258.17 μm(图3: F)。注射dsTaVgR组番茄潜叶蛾卵巢管明显缩短,成熟卵粒数和卵粒大小明显小于注射dsGFP组的,大部分卵粒卵黄蛋白沉积较少,呈空瘪状态(图3: G)。
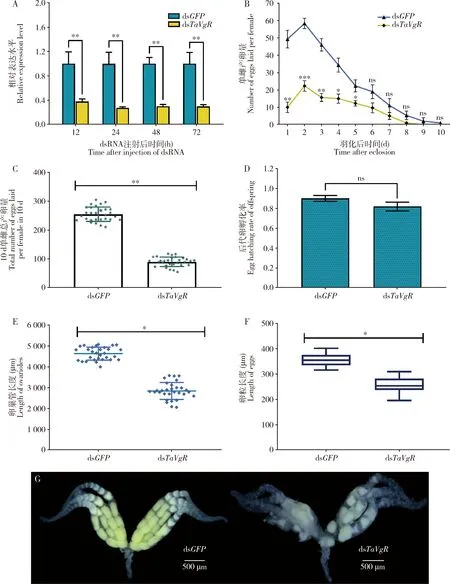
图3 TaVgR RNAi对番茄潜叶蛾卵巢发育及繁殖力的影响Fig. 3 Effect of RNAi of TaVgR on the ovarian development and fecundity of Tuta absolutaA: 注射dsRNA不同时间后TaVgR的表达量Expression levels of TaVgR at different time after injection of dsRNA; B: 注射dsTaVgR后1-10 d的单雌产卵量Number of eggs laid per female at 1-10 d after injection of dsTaVgR; C: 注射dsTaVgR后的10日单雌总产卵量Total number of eggs laid per female in 10 d after injection of dsTaVgR; D: 注射dsTaVgR后后代卵的孵化率Egg hatching rate of offspring after injection of dsTaVgR; E: 番茄潜叶蛾雌蛹注射dsTaVgR后成虫羽化24 h时卵巢管长度变化Changes in the length of the ovarioles of the 24 h-old adults after injection of dsTaVgR into the female pupae of T. absoluta; F: 番茄潜叶蛾雌蛹注射dsTaVgR后成虫羽化24 h时成熟卵粒长度变化Changes in the length of mature eggs of the 24 h-old adults after injection of dsTaVgR into the female pupae of T. absoluta; G: 番茄潜叶蛾雌蛹注射dsTaVgR后成虫羽化24 h时的卵巢形态变化Morphological changes of the ovaries of the 24 h-old adults after injection of dsTaVgR into the female pupae of T. absoluta. 向番茄潜叶蛾雌蛹注射dsRNA(500 ng/头)进行RNA干扰试验。图中数据为平均值±标准误;星号表示对照组(注射dsGFP)和处理组之间存在显著差异(*P<0.05;**P<0.01;***P<0.001)(t检验),而ns代表无显著性差异(P>0.05, t检验)。RNAi experiment was performed by injecting dsRNA (500 ng/individual) into the female pupae of T. absoluta. Data in the figure are mean±SE. The asterisk indicates significant difference between the control group (injected with dsGFP) and the treatment group (*P<0.05;**P<0.01;***P<0.001) (t-test), while ns represents no significant difference (P>0.05, t-test).
3 讨论
研究表明,VgR同一保守结构域在不同昆虫中的拷贝数及内部重复基序有所差异。本研究表明,番茄潜叶蛾TaVgR包含2个LBD,其中LBD1有4个重复基序,而其他昆虫的LBD1与之不同,如褐飞虱、黑尾叶蝉、埃及伊蚊、黑腹果蝇和家蝇具有5个,蜜蜂仅有3个(陈利鹏等, 2016; Liuetal., 2016)。番茄潜叶蛾VgR的LBD2与多数已知鳞翅目昆虫相同,包含了7个LDLa基序,但同为鳞翅目的小菜蛾Plutellaxylostella则有8个(王加伟, 2016)。两个LBD后面均有1个EGF,不同昆虫的EGF均具有富含半胱氨酸残基的B型重复基序和YWTD基序,不同昆虫间LDLb重复基序数量不尽相同,这可能与其分子功能和VgR的内吞作用效应有关。番茄潜叶蛾VgR与西方蜜蜂、黑腹果蝇、埃及伊蚊等昆虫一样,含有1个OLSD,然而粘虫Mythimnaseparata、斜纹夜蛾、棉铃虫VgR均不含OLSD,其具体功能尚不明确(李杰, 2018)。番茄潜叶蛾仅有1个TMD,而烟粉虱Bemisiatabaci、豌豆蚜Acyrthosiphonpisum和地中海实蝇Ceratitiscapitata等昆虫的VgR具有2个TMD (郭建洋, 2010; Congetal., 2015)。不同物种VgR间的结构差异是否会产生功能多效性,是否会影响其与配体的识别作用等仍不清楚,还有待深入研究。
昆虫VgR在不同发育时期的表达量存在明显差异,大多数物种的表达峰值出现在成虫期,如粘虫VgR表达量随着成虫日龄的增加而升高,在羽化第5天达到峰值(李杰, 2018)。红火蚁Solenopsisinvicta、家蚕、棉铃虫和斜纹夜蛾的结果与之基本相似,在初羽化的雌成虫中VgR表达水平较高,且在卵黄发生时期持续升高(Chenetal., 2004; Tufail and Takeda, 2005; Zhangetal., 2016)。这与本研究中番茄潜叶蛾TaVgR基因的表达情况相符,随着卵黄发生过程其表达量逐渐升高(图2)。本研究进一步说明昆虫VgR与卵黄发生过程密切相关,并在前期表现较高表达水平,而在整个卵黄发生过程中呈现持续上升的趋势。组织特异性研究结果表明,番茄潜叶蛾TaVgR基因在雌虫卵巢中特异性高表达(图2: B),这与其他大部分昆虫的结果一致。卵巢是昆虫重要的生殖器官,表明VgR参与昆虫生殖发育的调控(Linetal., 2015; Luetal., 2015)。但也有研究表明,不同昆虫VgR组织表达模式存在差异,如粘虫、柞蚕和野桑蚕Bombyxmandarina幼虫脂肪体内VgR高表达,可能通过影响昆虫的能量和营养代谢调控幼虫的生长发育(Liuetal., 2011; 付维维, 2013; 李杰, 2018)。西方蜜蜂VgR在头部、咽下腺和中肠有较高的表达,推测其在种群的信息交流、行为活动和社会分工等生命活动中发挥着重要作用(Guidugli-Lazzarinietal., 2008)。
为了探索TaVgR基因在番茄潜叶蛾生殖发育中的作用,本研究选择在番茄潜叶蛾TaVgR基因表达启动前期(4日龄雌蛹)注射dsTaVgR,发现dsTaVgR注射后TaVgR的表达量较对照组(注射dsGFP)明显下调,羽化后雌蛾产卵量显著降低,卵巢管变短且成熟卵粒体积变小,卵母细胞中沉积的卵黄含量显著减少(图3),导致其生殖力明显下降。在斜纹夜蛾蛹期,VgR表达的抑制破坏了卵母细胞对Vg的摄取,使血淋巴中的Vg积累,最终引起卵巢管发育受阻,产卵量显著降低(Shuetal., 2011)。通过对家蚕蛹第1, 4和7天注射dsVgR,发现雌蛾能正常羽化,但其中VgR沉默率较高的家蚕不能产卵,解剖可观察其卵巢管偏短,无卵子产生;而VgR沉默率较低的家蚕能交配产卵,但所产卵粒多为小型卵,且颜色偏白(Linetal., 2013);在褐色桔蚜Aphiscitricida中,AcVgR基因表达量的下降导致其若虫-成虫过渡延迟、生殖前期延长和生殖期缩短,从而导致其卵巢发育减慢且后代数量明显减少(Shangetal., 2018)。
综上所述,本研究利用RT-PCR技术克隆获得番茄潜叶蛾TaVgR基因的cDNA全长序列,该基因在番茄潜叶蛾雌成虫和卵巢中表达量较高。进一步抑制TaVgR基因的表达后,番茄潜叶蛾雌虫的卵巢发育受阻,其卵黄含量明显减少,卵巢管变短,且产卵量显著下降,表明TaVgR基因在番茄潜叶蛾的卵巢发育和繁殖过程中发挥了重要作用。
猜你喜欢
资源导刊(信息化测绘)(2022年4期)2022-11-24
家庭百事通·健康一点通(2019年7期)2019-08-16
植物保护(2018年1期)2018-05-14
家庭科学·新健康(2017年12期)2018-01-09
现代农业科技(2017年13期)2017-08-09
小溪流(画刊)(2017年6期)2017-07-10
农业科技与装备(2017年1期)2017-06-09
现代园艺(2016年7期)2017-01-09
妇女之友(2016年10期)2016-12-27
现代园艺(2016年9期)2016-07-16
