
二氢杨梅素抑制黄曲霉菌生长的机制
2022-07-29 02:40乔苏瑞谢岩黎
食品科学 2022年13期
李 倩,赵 颖,乔苏瑞,谢岩黎
(河南工业大学粮油食品学院,河南省粮油食品安全检测与控制重点实验室,河南 郑州 450001)
黄曲霉菌(,AF)属子囊菌门、丛梗孢科、曲霉属,是一种常见的腐生型好氧真菌。AF易侵染多种粮食作物,如小麦、玉米、花生,不仅损害其品质,而且影响经济效益。据报道,我国每年因霉菌污染而造成的粮食损失高达2 100万 t,占全国粮食总产量的4.2%,是每年新增粮食产量的4 倍多。作为一种病原性真菌,AF极易通过呼吸道、口耳眼鼻、皮肤等部位进入人和动物体内,引起支气管炎、肺泡坏死、外耳炎、角膜炎、鼻窦炎等多种疾病,危害人畜身体健康。此外,AF可产生多种次级代谢产物——黄曲霉毒素(aflatoxin,AFT),其中黄曲霉毒素B(aflatoxin B,AFB)毒性最强,约为氰化钾的10 倍、砒霜的8 倍、二甲基亚硝胺的75 倍、三聚氰胺的416 倍,被世界卫生组织(World Health Organization,WTO)列为I类致癌物。AFT对光、热、酸较稳定,280 ℃高温下才可裂解,因此一般烹调加工温度难以破坏。
二氢杨梅素(dihydromyricetin,DMY)又名白蔹素、蛇葡萄素,属于二氢黄酮类化合物,多存在于葡萄科、杨梅科、杜鹃科、藤黄科、大戟科及柳科等植物中,其中在葡萄科藤茶中含量最高。以往研究表明,DMY具有广泛的生物活性,如抗炎、抗肿瘤、抗氧化、调节血糖血脂、保肝护肝等,因此广泛应用于药品、食品、化妆品等行业。例如,目前以DMY为功效添加成分已研制出速溶茶、显齿蛇葡萄总黄酮含片、DMY解酒保肝胶囊等保健食品。除此之外,DMY可以有效抑制食源性细菌,如金黄色葡萄球菌、枯草芽孢杆菌、大肠杆菌、副伤寒沙门氏菌和铜绿假单胞菌,DMY对其最小抑菌质量浓度(minimum inhibitory concentration,MIC)分别为0.63、1.25、0.31、0.63、0.31 mg/mL。在熊皓平等的研究中,DMY对AF、青霉、黑曲霉、毛霉、根霉等真菌也具有显著抑制效果。
作为一种天然植物提取物,DMY成本低廉、来源广泛,具有极大的开发前景。本实验首先检测DMY对AF的抗真菌活性,在此基础上通过测定细胞壁/膜完整性、细胞内容物释放量、呼吸速率、细胞外相对电导率和pH值探究DMY潜在的抗真菌机制,最后评估其在粮食中的抑菌效果,以期为天然抑菌剂的开发提供理论依据。
1 材料与方法
1.1 菌株、材料与试剂
AF菌株(CGMCC 3.6304) 中国微生物菌种保藏中心(China General Microbial Culture Collection Center,CGMCC);DMY(纯度≥98%) 上海阿拉丁生化科技股份有限公司;卡尔科弗卢尔荧光增白剂(calcofluor white,CW) 上海懋康生物科技有限公司;抗荧光猝灭剂上海生工生物工程股份有限公司;碘化丙啶(propidium iodide,PI) 北京索莱宝科技有限公司;花生、玉米购于郑州永辉超市。
1.2 仪器与设备
LDZX-50KBS立式压力蒸汽灭菌器 上海申安医疗器械厂;AL204电子天平 梅特勒-托利多仪器(上海)有限公司;101-E电热鼓风干燥箱 北京市光明医疗仪器厂;SW-CJ-2D双人单面净化工作台 苏州净化设备有限公司;PYX-DHS-40X50-BS隔水式电热恒温培养箱上海跃进医疗器械厂;THZ-98B双功能汽浴恒温振荡器上海诵诚实业发展有限公司;BM-37XBC倒置生物显微镜上海彼爱姆光学仪器制造有限公司;DFC 7000T荧光显微镜德国徕卡公司;UV-6100S紫外-可见分光光度计上海美谱达仪器有限公司;DZS-706-C多参数分析仪上海雷磁仪器有限公司。
1.3 方法
1.3.1 DMY对AF抑制作用实验
AF于(28±2)℃下在固体沙氏培养基(Sabouraud dextrose agar,SDA)(含40 g/L葡萄糖、10 g/L蛋白胨和20 g/L琼脂)上培养7 d,然后用打孔器在菌落中央打孔制得AF菌饼(直径0.6 cm)备用。
孢子悬浮液制备:向AF的SDA表面倾注5 mL生理盐水并用涂布器刮取孢子,吸取含有孢子的生理盐水并用滤纸过滤,所得滤液即为孢子原液。充分吹打混匀后用血球计数板计数,加入液体沙氏培养基(Sabouraud dextrose broth,SDB)(含40 g/L葡萄糖和10 g/L蛋白胨)稀释孢子悬浮液使孢子浓度为5×10个/mL,备用。
用无水乙醇配制不同质量浓度的DMY溶液,避光储存于4 ℃冰箱。实验所用水均为去离子水。
1.3.1.1 DMY对AF孢子萌发抑制率的测定
DMY对AF孢子萌发抑制率的测定参考文献[16]的方法并稍作修改。在96 孔板加入20 μL孢子悬浮液(5×10个/mL)和20 μL DMY溶液(终质量浓度分别为0(对照,后同)、0.5、1、2、4、8、16 mg/mL)。将96 孔板置于恒温培养箱(28±2)℃中孵育48 h,每24 h于倒置显微镜下观察孢子萌发形态,显微镜拍照后计算孢子萌发抑制率(视野中未萌发孢子数占总孢子数比值),DMY对AF孢子的最小抑菌质量浓度(minimum inhibitory concentration,MIC)指DMY完全抑制孢子萌发的最小质量浓度。
1.3.1.2 DMY对AF菌丝生长抑制率的测定
在培养皿(直径6 cm)中加入10 mL含DMY(质量浓度分别为0、2、4、8、16 mg/mL)的SDA。培养基凝固后,在培养基中央打孔(直径0.6 cm),然后嵌入AF菌饼,密封后于恒温培养箱静置72 h,每24 h测量菌落直径。72 h后菌落直径无变化且菌丝生长抑制率为100%的最小质量浓度即为DMY对AF菌丝的MIC。菌丝生长抑制率按式(1)计算。

式中:和分别代表对照组和实验组在SDA上的平均菌落直径/cm。
1.3.2 孢子活力检测和菌丝生物量分析
1.3.2.1 孢子活力检测
采用平板记数法检测孢子活力,结果以孢子存活率表征。参照1.3.1.1节方法,制备DMY终质量浓度分别为0、4、8、16 mg/mL的孢子悬浮液。然后取100 μL孢子悬浮液稀释10倍,取200 μL均匀涂布于9 cm培养皿(每个培养皿含30 mL SDA,每个质量浓度做3个重复),置于恒温培养箱48 h,根据菌落数按式(2)计算孢子存活率。

式中:为用于涂布的孢子数量;为培养48 h后记录的菌落数。
1.3.2.2 菌丝生物量分析
将100 μL孢子悬浮液(5×10个/mL)加入20 mL SDB中,于(28±2)℃、150 r/min振荡培养24 h,然后加入2 mL DMY溶液(终质量浓度分别为0、2、4 mg/mL),继续振荡培养一定时间后用滤纸过滤培养液收集菌丝,用去离子水冲洗两次以洗去培养液。0-24组代表培养24 h收集的菌丝,0-48、2-48、4-48组代表在48 h收集的DMY终质量浓度分别为0、2、4 mg/mL的菌丝。将各组收集的菌丝在80 ℃电热恒温干燥箱中干燥24 h后分别称质量,以表征菌丝生物量。
1.3.3 细胞壁完整性测定
将50 μL孢子悬浮液(5×10个/mL)加入10 mL SDB中,于恒温培养箱静置培养24 h。加入1 mL DMY溶液(终质量浓度分别为0、1/2 MIC和MIC)。再静置培养24 h,取菌丝或培养液进行细胞壁完整性、细胞膜完整性和细胞内容物释放量的分析与测定。
细胞壁完整性分析参考文献[17]的方法并稍作修改。取少量菌丝置于载玻片上,用10%(质量分数)KOH溶液和CW染色,并滴加抗荧光猝灭剂保护荧光,最后用荧光显微镜观察并拍照(放大20 倍)。
1.3.4 细胞膜完整性测定
参考文献[18]的方法进行染色。取少量1.3.3节培养的菌丝置于载玻片上,滴加10 μL PI染料(1 μL/mL)染色,避光静置20 min后,用0.01 mol/L磷酸盐缓冲液(pH 7.0)洗去未结合染料。滴加抗荧光猝灭剂并用荧光显微镜观察拍照,采用Image J软件计算PI荧光强度,以表征细胞膜完整性。
1.3.5 细胞内容物释放量的测定
细胞内容物释放量实验参考文献[19]的方法并稍作修改。取2 mL 1.3.3节培养液,5 000 r/min离心10 min,取上清液测定260 nm波长处光密度值(OD),测定时用新鲜的空白SDB校准。
1.3.6 相对电导率和pH值的测定
相对电导率和pH值的测量参考文献[20]。将500 μL孢子悬浮液(5×10个/mL)加入100 mL SDB中,(28±2)℃、150 r/min振荡培养24 h。加入10 mL DMY溶液(终质量浓度分别为0、1/2 MIC、MIC),然后再振荡培养24 h。收集0.5 g菌丝悬浮在 20 mL 纯水中,使用多参数分析仪测定0、5、10、20、40、60、80、100、120、140、160、180 min时混合液的电导率和pH值。检测完毕后立即煮沸5 min,冷却至室温后测定混合液的电导率。按式(3)计算相对电导率。

式中:和分别代表样品煮沸前后的电导率/(μS/cm);和分别代表纯水煮沸前后的电导率/(μS/cm)。
1.3.7 呼吸抑制率的测定
DMY对AF孢子呼吸代谢影响的测定参考文献[21]。向试管中加入7.2 mL磷酸盐缓冲液(0.01 mol/L、pH 7.0)和2 mL孢子悬浮液(5×10个/mL)静置5 min后,加入0.8 mL 1%(质量分数)葡萄糖,混合均匀后静置10 min,然后加入1 mL DMY溶液(终质量浓度分别为0、1/2 MIC、MIC)混匀后静置10 min,以等体积去离子水作为空白组。待溶液稳定后用溶解氧测定仪测定溶解氧浓度,30 min后再次测定培养液的溶解氧浓度,计算30 min内溶解氧浓度变化(=-)。按式(4)计算呼吸抑制率。
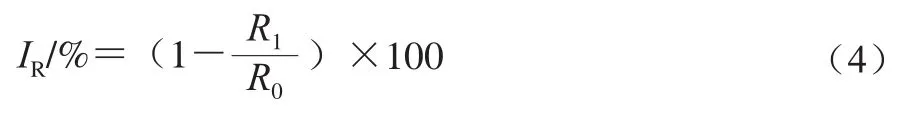
式中:和分别代表空白组和DMY样品组在30 min内溶解氧浓度变化。
1.3.8 DMY对花生和玉米籽粒储存过程中AF的体外抑菌效果分析
DMY对花生和玉米籽粒储存过程中AF的抑制效果测定参考文献[22]的方法并稍加修改。花生和玉米籽粒用无菌水洗涤,然后用紫外线照射20 min。在超净工作台短暂干燥后,将10 粒花生均匀放入直径6 cm培养皿中。将400 μL孢子悬浮液(5×10个/mL)与40 μL DMY溶液混合(DMY终质量浓度分别为0、1/2 MIC和MIC),随后均匀涂抹于花生表面(40 μL/粒)。在恒温培养箱(28±2)℃中放置72 h,每24 h统计1 次受污染花生的数量并拍照。以相同方式测定DMY对玉米籽粒的抗真菌作用。按式(5)计算花生/玉米籽粒污染率。

式中:和分别代表72 h后花生/玉米籽粒的污染个数和实验所用花生/玉米籽粒总数。
1.4 数据处理与统计分析
每组实验重复至少3 次,采用Origin 2017软件处理数据,结果表示为平均值±标准差。采用SPSS 20.0软件进行单因素方差分析,通过Duncan多重比较进行显著性分析,<0.05表示差异显著。采用Illustrator CC 2018软件作图。
2 结果与分析
2.1 DMY对AF孢子和菌丝的MIC
如图1A所示,DMY对AF孢子萌发有显著抑制作用。其中,0、0.5 mg/mL时,因DMY质量浓度过低,暂无抑制作用;当DMY为1 mg/mL时,24 h和48 h时抑制率分别为24.41%和9.05%;2 mg/mL的DMY处理48 h后80.45%的孢子未萌发;当DMY质量浓度不低于4 mg/mL时,孢子萌发抑制率达到100%,即DMY对AF孢子萌发的MIC为4 mg/mL。如图1B所示,DMY对AF菌丝生长同样有显著抑制作用。当DMY质量浓度为2 mg/mL时,24、48、72 h时菌丝生长抑制率分别为95.00%、29.88%和16.98%;当DMY质量浓度不低于4 mg/mL时,抑制率达到100%,即DMY对AF菌丝生长的MIC为4 mg/mL,高于香芹酮(MIC=1.5 mg/mL)、槲皮素(MIC=1.0 mg/mL)等,但低于罗勒烯(MIC=8.0 mg/mL),这种差异可能与AF孢子及不同菌株浓度有关。相同菌株接种浓度相同时,邻香兰素、2-羟基-4-甲氧基苯甲醛(2-hydroxy-4-methoxybenzaldehyde,HMB)、丹皮酚、辛醛、壬醛、癸醛抑制AF菌丝生长的MIC分别为0.1、0.07、0.625、1.65、1.65、4.15 mg/mL。虽然DMY的MIC接近或高于上述天然化合物,但其作为抑菌剂仍然潜力巨大,这是由于:1)醛类化合物易挥发,在实际使用过程中不可避免存在损失,而DMY稳定性较高;2)DMY在37.5 mg/mL和0.01 mg/mL质量浓度下能够分别发挥保护肝脏和抗肿瘤活性,说明DMY还具有多重功效。这些结果为后续抑菌机理的探究奠定了良好的基础。


图1 DMY对AF孢子萌发(A)和菌丝生长(B)抑制率Fig. 1 Inhibition rate of DMY on spore germination (A) and mycelial growth (B) of A. flavus
2.2 DMY处理后AF孢子存活率和菌丝生物量
为了检测DMY在MIC质量浓度及以上处理后的孢子活力,本研究测定了孢子存活率和菌丝生物量。如图2A所示,对照组孢子存活率接近100%,而4、8、16 mg/mL的DMY处理组中孢子活力下降,孢子存活率分别为80%、65%和65%,表明DMY处理从一定程度上削弱了孢子的活力,且呈现剂量依赖关系。此外,进一步对菌丝的生物量进行量化分析,如图2B所示,对照组振荡培养24 h的菌丝生物量为4.95 mg,对照组振荡培养48 h菌丝生物量达57.65 mg。与对照组振荡培养48 h相比,经2 mg/mL的DMY处理后,菌丝生物量显著下降,下降比例为55.51%,4 mg/mL的DMY对菌丝生物量的降低效果最为显著,下降比例为88.93%。同时,0-24组和4-48组的菌丝生物量无显著差异(>0.05),由此可知,4 mg/mL的DMY几乎能够完全抑制菌丝的生长。

图2 DMY处理后AF孢子存活率(A)和菌丝生物量(B)Fig. 2 Spore survival rate (A) and mycelial biomass (B) of A. flavus treated with DMY
2.3 细胞壁完整性观察结果
多糖是真菌细胞壁的主要成分,为细胞壁提供强度和硬度,如几丁质和-1,3-葡聚糖。细胞壁在向心力作用下向菌丝中心局部延伸形成隔膜,把菌丝内部分隔为不同的小室,且相邻的小室之间有细胞质的互通。已知荧光白染料可与几丁质特异性结合,而几丁质是细胞壁(隔膜)的主要成分,因此,荧光强度与细胞壁完整性有关。由图3可知,对照组菌丝隔膜完整且清晰透亮,表明几丁质含量较高。与对照组相比,1/2 MIC和MIC组隔膜数量依次减少,荧光强度依次减弱,表明DMY处理后,AF菌丝中几丁质含量减少,即细胞壁完整性被破坏。相似地,植物源天然化合物中的邻香兰素、HMB、丹皮酚等均能不同程度的溶解隔膜,在具有相似隔膜的酸腐菌中也发现,肉桂醛能够溶解隔膜,植物精油如紫苏精油能够从基因水平调控葡聚糖合成酶、几丁质酶等的表达。以上研究表明,细胞壁可能是植物源天然化合物抑菌的一个靶标,然而,哪些天然化合物能够被成功开发为靶向抑菌剂仍有待进一步研究。

图3 DMY处理后AF菌丝CW荧光染色图Fig. 3 CW fluorescence staining images of A. flavus treated with DMY
2.4 细胞膜完整性与细胞内容物释放量变化
对PI染色后的菌丝荧光强度进行量化,结果如图4A所示。随着DMY质量浓度的增加,相对荧光强度显著增加,1/2 MIC和MIC组分别是对照组的36.49 倍和257.69 倍。结果表明,DMY破坏了AF真菌细胞膜完整性,且这种破坏作用与DMY质量浓度成正比关系。已有许多研究报道醛类化合物具有疏水性,能够破坏细胞膜,导致细胞膜的通透性升高、完整性破坏。一些黄酮类化合物对真菌细胞膜的破坏作用与之相似,如乔松素和白杨素对意大利青霉以及黄芩素对烟曲霉的破坏作用。
细胞膜完整性的破坏可直接导致细胞内容物的渗出,因此,通过测定260 nm波长处的光密度值(OD)分析细胞内容物的释放情况,结果如图4B所示。当DMY质量浓度分别为0、1/2 MIC、MIC时,OD依次增加(分别为0.14、0.58和0.97)。与对照组相比,1/2 MIC组和MIC组光密度值分别增加了3.14 倍和5.93 倍。细胞内容物释放量的显著增加再次印证了细胞膜通透性的提高。
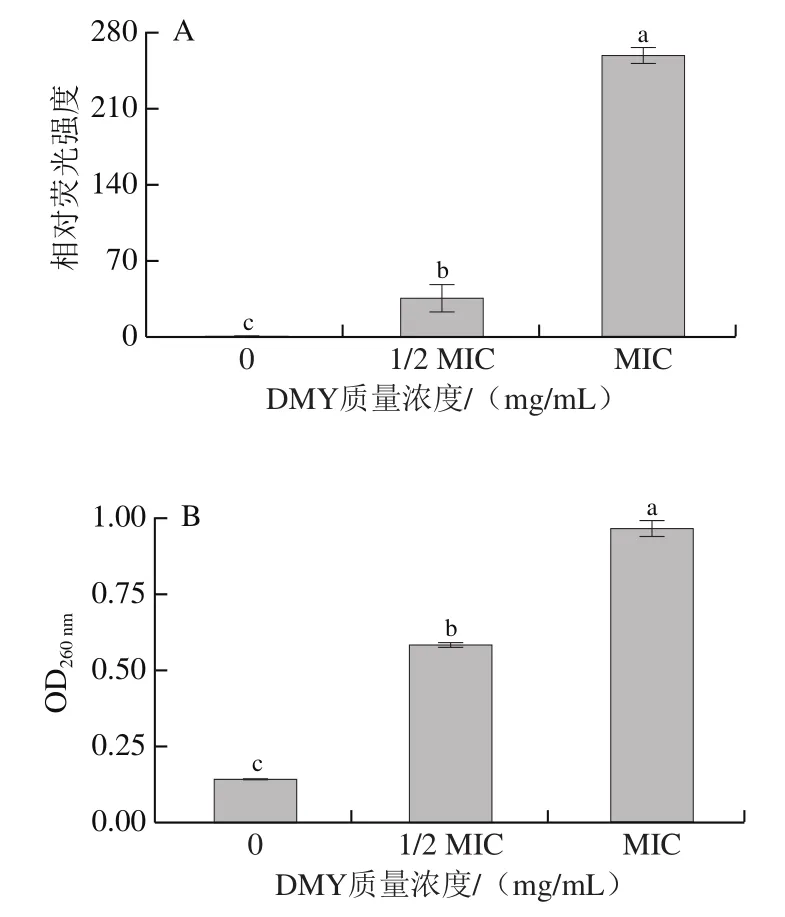
图4 DMY对AF细胞膜完整性(A)和细胞内容物释放量(B)的影响Fig. 4 Effects of DMY on the integrity of cell membrane (A) and the release of cell contents (B) of A. flavus
2.5 AF的相对电导率和pH值改变情况
由于渗出的细胞内容物具有导电性,因此,进一步通过细胞外相对电导率的变化趋势研究DMY对细胞膜破坏的程度。如图5A所示,对照组和DMY处理组均呈现了相对电导率升高的趋势。对照组的升高可能是细胞内外的渗透压导致的,1/2 MIC组的升高趋势和对照组接近,在180 min时约为43%,MIC组的升高最为显著,180 min时约为69.63%。这些结果表明细胞内的导电物质穿过完整性遭到破坏的细胞膜,且随着时间的延长,渗出的电解质逐渐增多。上述PI染色、OD及相对电导率的检测结果均间接表明了DMY处理后细胞膜受到了损伤,且该损伤呈现剂量依赖关系。
为了研究这些电解质的酸碱度,测定了不同质量浓度DMY处理后AF细胞外的pH值。如图5B所示,与对照组相比,DMY处理组细胞外pH值均升高,但随着时间变化,各组pH值均呈现略微下降的趋势,这可能与酸性物质的渗出有关。在180 min检测时间内,DMY处理组的pH值均明显高于对照组,推测处理组中有更多碱性物质渗出。pH值的变化趋势在不同天然产物处理的不同菌株中表现不同,如柠檬醛、辛醛和-松油醇3种挥发性化合物降低了柑桔地霉的pH值,HMB和丹皮酚均提高了AF菌丝胞外pH值。因此,pH值的变化趋势可能更多地与菌株类型有关。细胞膜位于真菌细胞壁和细胞质的中间,不仅是细胞的一层屏障,更能够参与调控、介导多种细胞功能,因此,细胞膜何种组分(脂质、蛋白等)、何种结构受到DMY的破坏仍有待深入研究。

图5 DMY处理下AF细胞外相对电导率和pH值的变化Fig. 5 Changes in extracellular relative conductivity and pH of A. flavus treated with DMY
2.6 DMY对AF呼吸代谢的影响
通过检测细胞外溶液中的溶解氧,能够了解AF消耗氧气的情况,进而通过计算呼吸抑制率分析DMY对AF呼吸的抑制作用。如图6所示,与对照组相比,DMY对AF的呼吸抑制率呈现剂量依赖关系,1/2 MIC和MIC组的呼吸抑制率分别为16.99%和25.82%。已有研究报道部分植物源天然化合物对真菌的呼吸代谢有抑制作用,如柠檬醛在MIC时抑制率达到21.20%,HMB在MIC时的抑制率达到33.33%,癸醛在MIC时抑制率达到58.94%。根据本实验结果虽尚无法全面揭示呼吸抑制原理,但由先前研究推测,该抑制作用可能与三羧酸循环途径或线粒体功能的破坏有关。

图6 DMY对AF的呼吸抑制率Fig. 6 Inhibition rate of DMY on A. flavus respiration
2.7 DMY对花生和玉米籽粒体外抑菌作用
花生和玉米易受AF侵染,因此,在上述人工培养液抑菌实验的基础上,本研究进一步评估了DMY对于防控AF污染的效果。AF的生长情况如图7所示,培养24 h时,除了两个MIC组,剩余花生和玉米籽粒上均观察到孢子的萌发。培养48 h时,对照组花生和玉米籽粒上均可观察到AF菌丝。培养72 h时,即便花生和玉米籽粒表面涂布有1/2 MIC的DMY,AF的生长情况也和48 h的对照组相似,而涂布MIC的DMY后花生和玉米籽粒表面则均无孢子萌发。对图7污染情况的量化结果如图8所示,培养72 h时,花生和玉米籽粒AF抑制率达到100%的均为DMY的MIC处理组。与培养液抑菌结果相同,4 mg/mL的DMY能够完全抑制AF侵染花生和玉米籽粒,因此,DMY具有开发为新型抑菌剂的潜力。

图7 DMY对花生(A)和玉米(B)籽粒的体外抑菌作用Fig. 7 In vitro antifungal effect of DMY on peanut (A) and corn (B) kernels

图8 DMY处理下花生(A)和玉米(B)籽粒的污染率Fig. 8 Contamination rates of peanut (A) and corn (B) kernels treated with DMY
3 结 论
本实验旨在研究DMY对AF的抗真菌效果及其作用机理,首先进行AF孢子萌发和菌丝生长的最小抑菌实验,然后通过CW和PI染色、细胞内容物释放以及细胞外pH值和相对电导率变化检测细胞壁和细胞膜完整性,并进行呼吸速率检测及花生和玉米籽粒体外抑菌实验。研究获得的初步结论如下:DMY对AF孢子萌发和菌丝生长的MIC均为4 mg/mL,其通过破坏细胞壁和细胞膜的完整性发挥抑菌作用。DMY处理后线粒体是否遭到破坏仍需进一步的实验证明。此外,DMY能够有效抑制AF侵染花生和玉米籽粒,未来有望将其开发为新型抑菌剂应用于粮食及农产品的储藏。
猜你喜欢
国外畜牧学·猪与禽(2022年1期)2022-04-23
食品界(2021年7期)2021-07-19
分析化学(2018年11期)2018-01-16
杂草学报(2016年4期)2017-05-31
江苏农业科学(2016年1期)2017-05-17
课程教育研究·新教师教学(2016年14期)2017-04-12
科技经济市场(2016年5期)2017-02-05
海峡科技与产业(2016年11期)2016-12-26
文理导航(2016年32期)2016-12-19
中国校外教育(上旬)(2006年5期)2006-05-25
