
小兴安岭原始椴树红松林与次生杨桦林土壤氮组分特征
2022-10-10 11:37刁云飞丛喜东王琪瑶
中国农学通报 2022年26期
刁云飞,张 苏,丛喜东,杜 倩,王琪瑶,刘 学,4
(1东北林业大学,哈尔滨 150040;2黑龙江省生态研究所,哈尔滨 150081;3黑龙江牡丹江森林生态系统国家定位观测研究站,哈尔滨 150081;4黑龙江省森林生态与林业工程重点实验室,哈尔滨 150081)
0 引言
土壤作为陆地生态系统中碳氮的主要储存形式,在全球碳氮循环过程中发挥着重要的作用。土壤氮素是影响碳库动态变化的重要因素之一,土壤氮循环与土壤碳矿化、碳循环密不可分,土壤氮素在动植物体、微生物体、土壤有机碳、土壤矿物质等系统中进行转化和迁移,对有机质的矿化,腐殖质形成及腐殖质稳定性都发挥着重要作用。因此,氮素也成为森林生态系统生产力的重要限制因子[1]。已有研究表明,土壤中的全氮大部分为有机态氮,该部分氮只有经土壤微生物的矿化作用转化为无机氮,才可供植物吸收和利用[2]。无机氮主要以铵态氮和硝态氮等形式存在,尽管所占比例很小,但与土壤中的能量和物质转化关系密切,较土壤全氮更能及时反映土壤质量和植物生长状况[3]。无机氮极易被土壤微生物利用或者与土壤中的有机物及盐基离子结合,直接或间接影响土壤碳的输入及输出,从而极大地干预森林生态系统碳循环和碳累积过程[4]。因此,研究不同森林植被类型及同一植被不同生长阶段土壤氮组分差异,对提高土壤氮素养分的有效性具有重要意义。
本研究以小兴安岭地区顶级森林群落原始椴树红松林中的典型群落—椴树红松林(Tilia PinusKoraiensis)成熟林与中幼龄林,及次生杨桦林(Secondary Poplar Birch)为研究对象,分析比较椴树红松林成熟林与中幼龄林以及次生杨桦林的土壤氮组分特征,探讨不同发育阶段的椴树红松林以及次生杨桦林土壤氮组分差异及其影响因素,以期为估算不同林龄的椴树红松林及次生杨桦林土壤氮库及评价东北森林生态系统的土壤碳氮耦合机制提供数据支撑。
1 研究区域与研究方法
1.1 研究区域
研究区域位于黑龙江丰林国家级自然保护区内,地处小兴安岭南坡,海拔300~683 m,该区域属于温带大陆性季风气候。地理坐标为48°02′—48°02′N,128°58′—129°15′E,年平均气温为-5℃,年降水量为 650 mm,无霜期120天左右,地带性土壤为暗棕壤。丰林国家级自然保护区是中国目前保存最完整、最典型的原始红松林保护区,地带性植被为温带针椴树混交林,主要乔木树种有:红松(Pinus koraiensis)、紫椴(Tilia amurensis)、鱼鳞云杉 (Picea jezoensisvar.microsperma)、白桦(Betula platyphylla)、蒙古栎(Quercus mongolica)、水曲柳(Fraxinus mands-hurica)、花楷槭(Acerukurunduense)等。
1.2 样地布设与土壤取样
本研究选择小兴安岭林区最具代表性的温带顶极群落椴树红松林,根据其演替规律,以保护区内处于相同坡位不同林龄的典型原始椴树红松林—椴树红松林(20世纪60—70年代火烧后行成的中幼龄林阶段)、椴树红松林(成熟林阶段)及次生杨桦林为研究对象。在每个森林类型内随机3个20 m×20 m的固定样地。分别于生长季内5、6、7、8、9月,按蛇形布设9个固定采样点,于在每个样点按凋落物层(L、F层)、0~10、10~20、20~30 cm取样,一部分土样放在冰盒中,3天内运回实验室用于有机碳、容重、pH的测定;另一部分自然风干,过筛,用于土壤全氮、速效氮、硝态氮、铵态氮的测定。

表1 样地基本信息
1.3 指标测定方法与数据分析
土壤含水率采用烘干法[(105±2)℃,12 h]测定,每个样品重复测定3次;土壤容重(Bulk density,BD)采用环刀法测定;土壤总有机碳含量(Total organic carbon,TOC)采用重铬酸钾外加热法测定。土壤全氮含量(Total nitrogen,TN)采用凯式定氮法利用全自动凯式定氮仪测量,土壤硝态氮(NO-3-N)和土壤铵态氮(NH+4-N)采用连续流动分析仪测定;全磷含量(Total phosphorus,TP)采用钼锑抗比色法利用紫外光分度计测量,pH采用pH 213型酸度计测定,水土比为2.5:1。土壤总孔隙度计算公式如式(1)所示。

使用SPSS19.0软件中的方差分析(ANOVA)判断不同林龄土壤理化性质差异显著性;使用Pearson相关系数分析土壤总有机碳、氮含量、磷含量与土壤氮组分的相关性(P=0.05)。
2 结果与分析
2.1 3种林型土壤主要理化性质
由表2可知,在垂直剖面上,3种林型土壤总有机碳含量均随深度的增加而减少。就水平方向而言,不同林型总有机碳含量在0~10、10~20 cm从大至小依次均为椴树红松林成熟>次生杨桦林>椴树红松林中幼龄林。20~30 cm从大至小依次为椴树红松林中幼龄林>椴树红松林成熟林>次生杨桦林。不同林型同一土层土壤,总有机碳含量差异均不显著(P>0.05)。
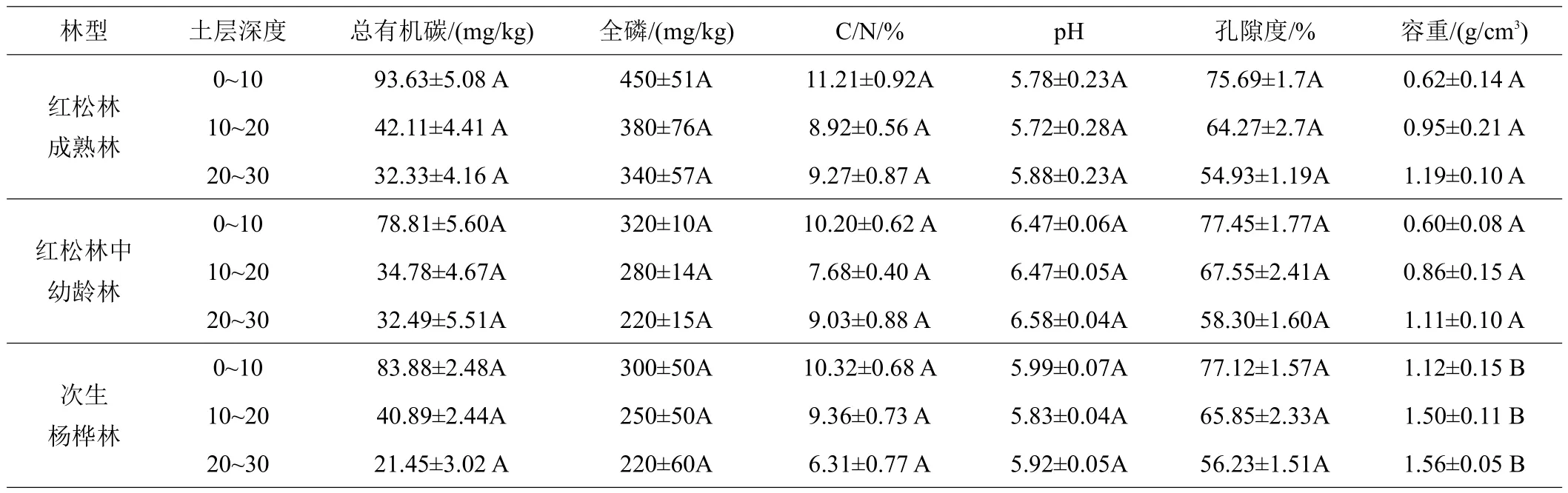
表2 椴树红松林成熟林及中幼龄林与次生杨桦林土壤理化性质
随着土层的加深,3种林型土壤含磷量均呈下降趋势。就水平方向而言,不同林型之间,全磷含量在0~10、10~20、20~30 cm 3个土层,从大至小均为椴树红松林成熟林>椴树红松林中幼龄林>次生杨桦林。不同林型同一土层土壤含磷量差异均不显著(P>0.05)。
随着土层的加深,3种林型土壤C/N变化均无一致规律性。就水平方向而言,不同林型之间,C/N在0~10 cm从大至小依次为椴树红松林成熟林>次生杨桦林>椴树红松林中幼龄林。10~20 cm,从大至小依次为次生杨桦林>椴树红松林成熟林>椴树红松林中幼龄林。20~30 cm从大至小依次为椴树红松林中幼龄林>椴树红松林成熟林>次生杨桦林。不同林型同一土层C/N差异均不显著(P>0.05)。
随着土层的加深,3种林型pH均呈稳定状态变化较小。就水平方向而言,不同林型之间,pH在0~10、10~20、20~30 cm 3个土层从大至小依次均为椴树红松林中幼龄林>次生杨桦林>椴树红松林成熟林。不同林型同一土层pH差异均不显著(P>0.05)。
随着土层的加深,3种林型土壤孔隙度均呈下降趋势。就水平方向而言,不同林型之间,孔隙度在0~10、10~20、20~30 cm 3个土层从大至小依次均为椴树红松林中幼龄林>次生杨桦林>椴树红松林成熟林。不同林型同一土层土壤孔隙度差异均不显著(P>0.05)。
随着土层的加深,土壤容重呈现逐渐增加的趋势。就水平方向而言,不同林型之间,容重在0~10、10~20、20~30 cm 3个土层从大至小依次均为次生杨桦林>椴树红松林成熟林>椴树红松林中幼龄林。不同林型同一土层,椴树红松林成熟林和中幼龄林土壤容重差异不显著(P>0.05),但二者与次生杨桦林土壤容重之间差异均为显著(P<0.05)。
2.2 3种林型土壤氮素组分比较
由表3可知,次生杨桦林与椴树红松林成熟林及中幼龄林0~10、10~20、20~30 cm 3个土层土壤全氮含量变化分别在 3.4~8.13、2.46~6.36、3.08~9.11 g/kg 之间。在垂直剖面上,3种林型全氮含量均随深度的增加而减少。3种林型含氮量0~10 cm分别与10~20、20~30 cm 2个土层差异显著(P<0.05),但10~20、20~30 cm之间差异不显著(P>0.05)。就水平方向而言,全氮含量在0~10、10~20 cm从大至小依次均为椴树红松林中幼龄林>次生杨桦林>椴树红松林成熟林。全氮含量在20~30 cm层,依次为次生杨桦林>椴树红松林中幼龄林>椴树红松林成熟林。不同林型同一土层,椴树红松林成熟林与中幼龄林之间土壤含氮量差异显著(P<0.05),但二者和次生杨桦林之间均不显著(P>0.05)。
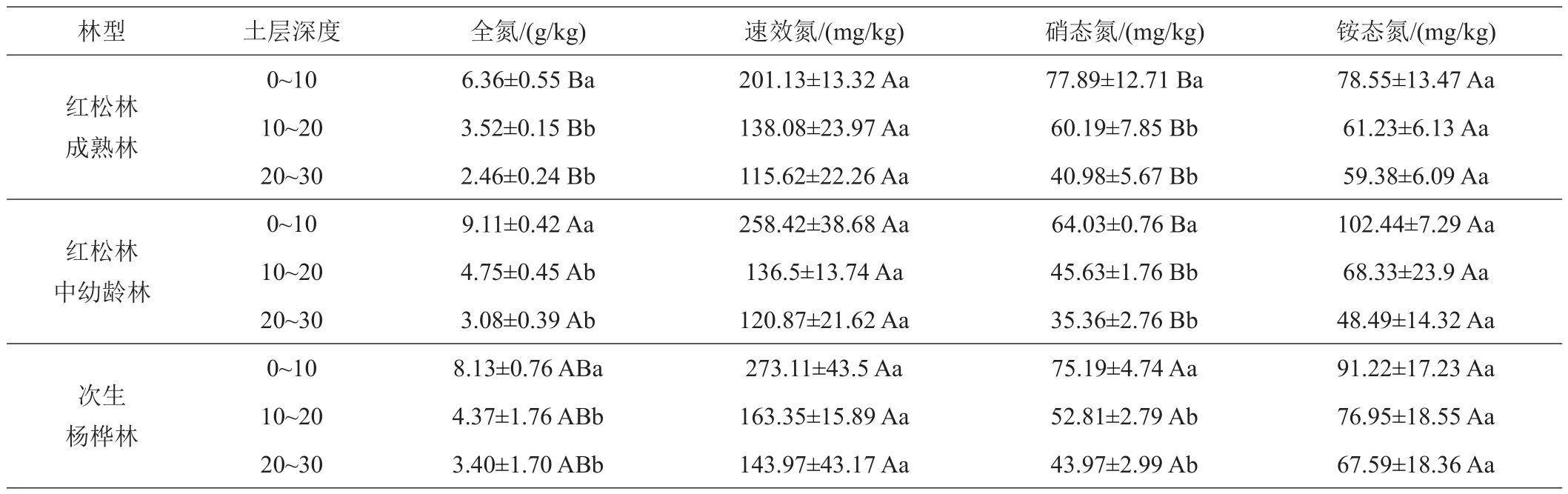
表3 次生杨桦林与阔叶红松林成熟林及中幼龄林土壤碳氮组分特征
次生杨桦林与椴树红松林成熟林及中幼龄林0~10、10~20、20~30 cm 3个土层土壤速效氮含量变化分别在143.97~273、115.62~201.13、120.87~258.42 mg/kg之间。在垂直剖面上,3种林型土壤速效氮含量均随深度的增加而减少。3种林型速效氮含量0~10 cm分别与10~20、20~30 cm 2个土层差异显著(P<0.05),但10~20、20~30 cm之间差异不显著(P>0.05)。就水平方向而言,速效氮含量在0~10 cm从大至小依次为次生杨桦林>椴树红松林中幼龄林>椴树红松林成熟林。但差异不显著(P>0.05)。速效氮含量在10~20 cm从大至小依次为次生杨桦林>椴树红松林成熟林>椴树红松林中幼龄林,但差异不显著(P>0.05)。20~30 cm速效氮含量从大至小依次为次生杨桦林>椴树红松林中幼龄林>松林成熟林,但差异不显著(P>0.05)。
次生杨桦林与椴树红松林成熟林及中幼龄林0~10、10~20、20~30 cm 3个土层土壤硝态氮含量变化分别在 43.97~75.19、40.98~77.89、35.36~64.03 mg/kg 之间。在垂直剖面上,3种林型土壤硝态氮含量均随深度的增加而减少。3种林型在0~10 cm与10~20 cm土层之间硝态氮含量差异显著(P<0.05),但10~20 cm与20~30 cm之间差异不显著(P>0.05)。就水平方向而言,硝态氮含量在0~10 cm从大至小依次为椴树红松林成熟林>次生杨桦林>椴树红松林中幼龄林。10~20 cm的变化规律均与0~10 cm一致。20~30 cm从大至小依次为次生杨桦林>椴树红松林成熟林>椴树红松林中幼龄林。不同林型同一土层,椴树红松林成熟林与中幼龄林之间土壤含氮量差异显著(P<0.05),但二者和次生杨桦林之间均不显著(P>0.05)。
次生杨桦林与椴树红松林成熟林及中幼龄林3个土层土壤铵态氮含量变化分别在67.59~91.22、59.38~78.55、48.49~102.44 mg/kg之间。在垂直剖面上,3种林型土壤铵态氮含量均随深度的增加而减少。3种林型铵态氮含量0~10 cm与20~30 cm之间差异显著(P<0.05),但10~20 cm与20~30 cm之间差异不显著(P>0.05)。就水平方向而言,铵态氮含量在0~10 cm从大至小依次为椴树红松林中幼龄林>次生杨桦林>椴树红松林成熟林。铵态氮含量在10~20 cm从大至小依次为次生杨桦林>椴树红松林中幼龄林>椴树红松林成熟林。20~30 cm铵态氮含量从大至小依次为次生杨桦林>椴树红松林成熟林>椴树红松林中幼龄林。不同林型同一土层,差异均不显著(P>0.05)。
2.3 3种林型土壤氮组分与凋落物及其他理化指标相关性分析
3种林型年凋落物与凋落物现存量见图1。如表4所示,土壤全氮、硝态氮、铵态氮与总有机碳、全磷、凋落物现存量等各项指标的相关性表明,土壤总有机碳与全磷、硝态氮、速效氮、孔隙度呈极显著正相关(P<0.01),与全氮呈显著相关(P<0.05),与含水率呈极显著负相关(P<0.01)。土壤全磷与总有机碳、硝态氮呈呈极显著正相关(P<0.01),与含水率呈极显著负相关(P<0.01)。土壤全氮与铵态氮、孔隙度呈极显著正相关(P<0.01),与总有机碳、速效氮呈显著相关(P<0.05)。土壤硝态氮与速效氮、孔隙度呈极显著正相关(P<0.01),与凋落物量现存量呈显著相关(P<0.05),与含水率呈极显著负相关(P<0.01)。铵态氮与全氮、孔隙度呈极显著正相关(P<0.01),与其他指标相关关系均不显著。速效氮与孔隙度呈极显著正相关(P<0.01),与全氮呈显著相关(P<0.05),与其他指标相关关系不显著。凋落物现存量与硝态氮显著相关(P<0.05),与其他指标相关关系不显著。含水率与总有机碳、全磷、硝态氮呈极显著负相关(P<0.01)。孔隙度与总有机碳、全氮、硝态氮、铵态氮、速效氮呈极显著正相关(P<0.01),与含水率呈显著负相关(P<0.05)。碳氮比与孔隙度呈显著相关(P<0.05),与其他指标相关关系不显著。
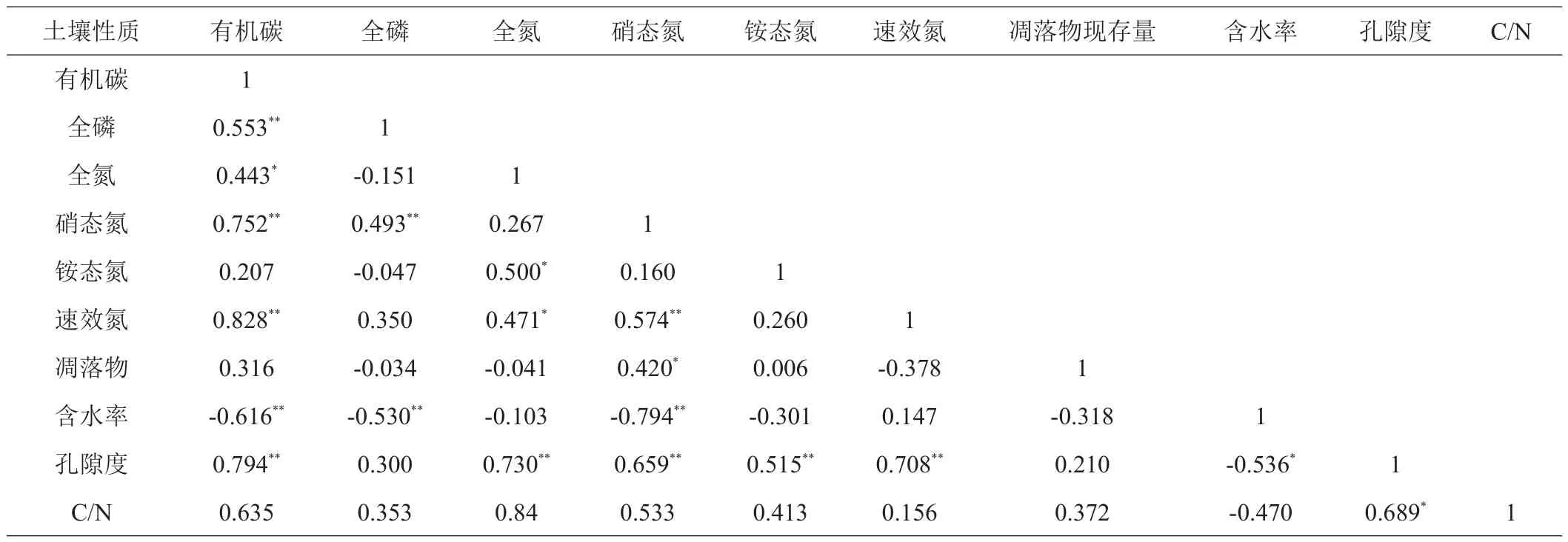
表4 土壤氮组分与凋落物及其他理化指标的相关性
3 结论与讨论
3.1 地上凋落物对土壤氮组分的影响
凋落物量是森林生态系统土壤氮的重要输入途径,决定着土壤有机氮库的大小[5]。张萍等[6]研究表明,凋落物、根系系统和不同的营养吸收方式都可能会影响微生物的组成和活性,进而影响土壤氮组分分布。马红亮等[7]的研究表明凋落物分解能增加土壤的肥力,同时也积累了大量的土壤有机碳。袁志忠等[8]研究表明凋落物的数量和质量决定着森林土壤养分性质,凋落物添加或减少能直接引起土壤养分数量上变化。凋落物的数量和质量决定着土壤微生物群落结构,进而影响土壤碳氮的循环过程[9-12]。王鑫等[13]研究表明,5年的地上凋落物数量处理并没有显著影响土壤有机碳和总氮的含量,但地上凋落物数量加倍处理相对于去除处理使10~20 cm土壤有机碳中木质素和次级脂肪酸的含量分别显著提高了28.24%和42.42%。本研究中3种林型年凋落物量和凋落物现存量依次均为次生杨桦林>椴树红松林中幼龄林>椴树红松林成熟林。3种林型的速效氮含量也同样排序。一般来说,凋落物分解,产生的养分及其残渣会进入到土壤中,改变凋落物的现存量和养分含量,也影响着土壤中的养分[14]。相关研究表明,土壤对植被供应氮素的数量,取决于土壤起始速效氮量,以及在植被生长期间土壤氮素的释放量,而生长期氮素的释放则主要受到土壤微生物数量和活性的影响。土壤中的微生物通过调节凋落物分解速率调控土壤中碳的周转,成为促使土壤中有机物和植物养分转化、循环的动力因子[8]。
因此,虽然本研究中次生杨桦林年凋落物数量明显高于椴树红松林成熟林,但由于凋落物质量和微生物群落结构以及其他土壤环境的影响,并没有明显促进次生杨桦林土壤有机碳氮的积累。凋落物对土壤有机质的影响可能更多取决于凋落物自身的特性和土壤条件,这与阮超越等[15]研究一致。此外,有研究表明,凋落物输入对土壤碳氮库的影响具有时间上的变异性,且地上地下凋落物输入对土壤碳氮库的贡献在不同的森林生态系统间表现不同。中温带森林生态系统对凋落物输入导致的土壤温湿度变化并不敏感,因此,森林生态系统的响应也相对较弱。本研究表明,地上凋落物对于土壤碳氮的贡献具有延后性,这与王鑫等[13]的研究一致。
3.2 土壤理化性质及土壤环境对氮组分的影响
土壤中碳、氮的相互关系是通过微生物连接起来的,因此土壤微生物对于土壤有机碳的分解至关重要[16]。土壤中的固氮细菌是氮元素转化的主要原因,在土壤植物根系的分布范围内最为活跃。土壤中的硝态氮和铵态氮都于无机氮,可以直接被植物利用,所以在0~10 cm土层含量较高。本研究中土壤表层0~10 cm的各种氮均高于其他剖面,这表明土壤氮也存在表层聚集现象[3]。其原因可能是在垂直方向上,氮元素被植物根系直接吸收,受生物循环的影响,使吸收的氮的营养元素归还于土壤的表层(固定与转化),致使氮元素在0~10 cm土层积累。
马寰菲等[17]研究表明,在土壤养分供应较低的生境下植物生长易受N限制,而在土壤养分供应较充足的生境下植物生长易受P限制。王泽西等[18]研究表明,川西天然次生林土壤中总氮、总磷和硝态氮含量增加会引起土壤活性养分可利用性提高,引起表层土壤微生物种群数量及其活性增加,促进土壤有机碳、氮的分解和转化,进而影响土壤有机碳氮及其活性组分含量的变化。本研究中,土壤有机碳与全磷、硝态氮、速效氮呈极显著正相关(P<0.01),与全氮呈显著正相关(P<0.05),进一步证实了这种相关性,且在不同的氮组分中,速效氮、硝态氮作为优势氮种,对小兴安岭原始椴树红松林及次生杨桦林植被生长及土壤有机碳的影响作用更显著。
研究表明[11,19-22],土壤物理性质作为土壤的基本性质之一,影响土壤结构性状和水源涵养能力,进而对不同海拔植被带土壤碳氮等矿质元素在土壤中沉积和循环产生重要影响。本研究中,土壤孔隙度与总氮、硝态氮、铵态氮、速效氮均呈显著正相关。土壤孔隙度对植物生长非常重要,一般来说,孔隙度约为55%~65%的壤土最利于植物生长,较好的土壤孔隙度有利于微生物活动、繁殖以及植物根系吸收水分,因此,3种林型中,椴树红松林成熟林相对于中幼龄林的土壤质地更为优良。然而,也有研究表明,不同的植被类型或植被群落物种多样性组合复杂多变,对土壤碳素和氮素的影响也各不相同。一般认为物种多样性高,其碳氮储量也高[23],但其影响程度仍存在不确定性[24-27],有待于更多的数据进一步研究。
综上,小兴安岭椴树红松林和次生杨桦林地上凋落物对于土壤碳氮的贡献具有延后性,地上凋落物对土壤碳氮的影响更多取决于凋落物自身的特性和土壤条件。不同的氮组分中,速效氮、硝态氮作为优势氮种,对小兴安岭椴树红松林植被生长及土壤有机碳的影响作用更显著。3种林型土壤氮组分差异是土壤理化性质、凋落物、土壤环境等多种因素综合作用的结果。
猜你喜欢
建材发展导向(2022年10期)2022-07-28
纺织标准与质量(2022年1期)2022-07-12
当代水产(2022年4期)2022-06-05
福建轻纺(2022年4期)2022-06-01
水土保持学报(2022年3期)2022-05-26
中国药学药品知识仓库(2022年8期)2022-05-09
中国瓜菜(2022年1期)2022-02-18
中国生态农业学报(中英文)(2022年1期)2022-01-15
当代陕西(2020年24期)2020-02-01
当代陕西(2020年24期)2020-02-01
