
甘南州不同退化程度高寒草甸植被及土壤特性的演化规律
2022-10-13 11:17李雪萍许世洋李敏权王建喜郭致杰漆永红荆卓琼李建军谢志军
生态学报 2022年18期
李雪萍,许世洋,李敏权,王建喜,郭致杰,漆永红,荆卓琼,李建军,谢志军
1 甘肃省农业科学院植物保护研究所, 兰州 730070 2 甘肃农业大学草业学院, 兰州 730070 3 甘肃省农业科学院畜草与绿色农业研究所, 兰州 730070
青藏高原高寒草甸是世界高寒草甸和草地生态系统的重要有机组成,在生物多样性保护、生态系统可持续发展、全球气候调节和碳循环等过程中功不可没,仅东南部高寒草甸面积占全球48%以上[1—2]。地处青藏高原东部地区的甘南州高寒草甸物种资源种类及数量多、生产潜力大,是我国重要的畜牧业生产基地[3—4]。然而,随社会经济发展、人类活动和全球气候变暖等因素影响,草地发生逆行演替,草地“三化”、生物基因库锐减、水土流失、生产力下降等问题相继涌现,“生态屏障”、“生态服务”等功能衰退严重,甘南州18.08%草地仍呈持续性的恶化趋势[5—7]。
草地生态系统功能依赖于地上和地下生物群落之间的相互关系,在高寒草甸长期自然演替、地质及生物活动进程中,“植被-土壤-微生物”这一自然历史复合体是贯穿高寒草甸生态系统的一条主线,三者间不断地相互作用和反馈机制是研究草地退化规律的重要依据[8—10]。高寒草甸植被状况直接反映了其演替方向和进程,是表征气候、土壤肥力等生态因子波动重要指标[11—12]。研究发现,高寒草甸退化过程中草毡层逐渐剥离形成斑块[13],草地覆盖度、生物量、多样性下降,杂草类植被增多[14],优势种呈现出由禾草向杂草演化的趋势[15]。同时,对土壤生物多样性及功能产生负面影响[16],包括土壤结构、理化性质、土壤养分、土壤酶活性和土壤微生物数量等均发生显著改变[17—19]。针对甘南不同退化程度高寒草甸植被及土壤特性的演化规律,王小燕等[20]从季节尺度上对甘南州“黑土滩”型退化草甸研究发现,草地退化引起土壤脲酶活性降低,土壤速效磷、速效钾含量和碱性磷酸酶活性增加。姚宝辉等[21]对甘南州碌曲县高寒草甸进行研究发现,随退化程度加剧,土壤pH值增大,全氮和全磷含量降低。以及本课题组[22]对甘南州碌曲县高寒草地研究发现,随草地退化程度加深,植物种类减少,优势种发生变化,植被高度、盖度、鲜草产量下降,土壤细菌、放线菌含量减少,真菌含量增多等。
不同区域草地空间具有异质性,加之甘南州高寒草甸区位环境特殊、生态脆弱、气候敏感[23—24],致使草地植被及土壤特性因草地类型、退化程度等不同而存在差异。目前,甘南州不同区域退化高寒草甸植被及土壤特性差异不明,以及甘南州退化高寒草甸的植被及土壤特性整体演化规律研究不够深入。因此,本研究通过在甘南州退化高寒草甸3个主分布区(碌曲县、夏河县、合作市)多点采样,并分析不同区域、不同退化程度的植被及土壤特性的演化规律,以期为甘南州高寒草甸退化机理、生态修复的研究提供理论支撑。
1 研究区概况
研究区位于甘肃省甘南藏族自治州,是中国东亚季风区、西北干旱区和青藏高原高寒区三大自然区交汇处,其地势西北高、东南低,多数区域海拔在3000 m以上,年均气温4 ℃,年降雨量约600—810 mm,年平均日照时长2200—2400 h,属高原大陆季风性气候。研究区碌曲县、夏河县、合作市是甘南州高寒草甸主分布区,其植被包含草类植物种类繁杂,达30—40种/m2,无明显优势种群,但由于近年放牧压力大、鼠害严重和气候变化等,其退化严重,并导致优质牧草丰富度、产量下降严重,毒杂草滋生,土壤退化明显[23—24]。如表1所示,根据《高寒草甸退化、沙化、盐渍化的分级指标》(GB19377—2003),将研究区退化高寒草甸分为轻度退化、中度退化和重度退化3种类型[25]。
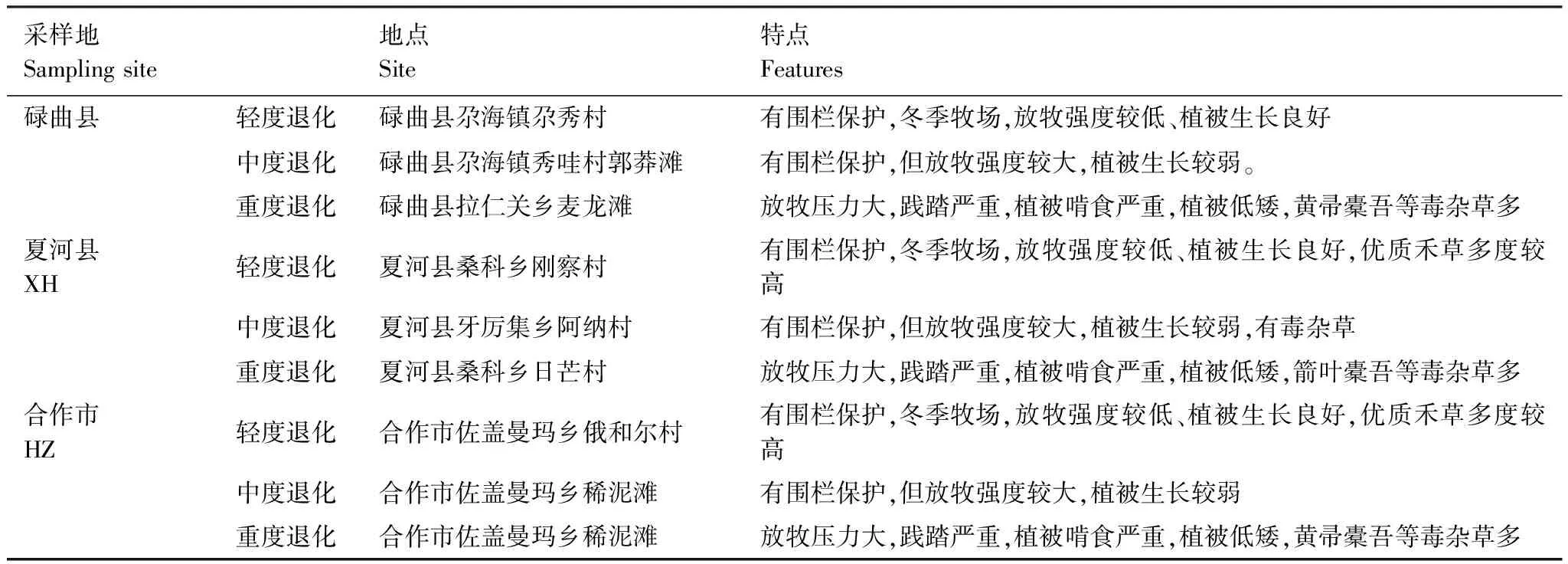
表1 样地概况
2 研究方法
2.1 草地调查与样品采集
于2019年8月底,赴甘南藏族自治州碌曲县、夏河县、合作市对不同退化程度高寒草甸的植被进行调查并采集土壤样品。每个退化程度草地调查面积约5000 m2,随机选取5个样方,样方大小为0.5 m×0.5 m。植被高度采用直尺测量,每样方随机测量10次,取平均值;植被的盖度采用针刺法测量[26];同时记录植被种类和数量,刈割样方内植物地上部分,估算草产量。土样采集深度为20 cm,每样方内用5点法取样,低温运输至实验室。
针对植物的种类及数量,计算Simpson多样性指数(D)、Shannon-Wiener多样性指数(H′)、均匀度(Evenness)、Brillouin指数(H)和Mcintosh指数(DMc)。
(1)
(2)
(3)
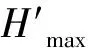
(4)
(5)
式中:n1为抽样中第1个物种的个体数量,n2为抽样中第2个物种的个体数量,n3为抽样中第3个物种的个体数量,如此类推;ni为抽样中第i个物种个体数量;N为抽样中所有物种的个体总和,s为物种数。
2.2 土壤理化性质测定
测定内容包括全氮含量、全磷含量、全钾含量、pH值、全盐含量、容重、孔隙度和土壤颗粒组成。采用半微量凯氏定氮法测定全氮含量,钼锑抗比色法测定全磷含量,火焰光度法测定全钾含量,pH值用PHS- 3C型测定计测定,重量法测定全盐含量,环刀法测定容重和孔隙度[27],采用Malvern公司生产Mastersizer2000激光粒度仪测定颗粒组成并依照美国农业部(USDA)制订标准进行划分:粒径<0.002 mm(黏粒)、0.002—0.05 mm(粉粒)、0.05—2 mm(砂粒)[28]。
2.3 土壤酶活性测定
采用靛酚比色法测定脲酶活性、3,5-二硝基水杨酸比色法测定蔗糖酶活性、磷酸苯二钠比色法测定磷酸酶活性、容量法测定过氧化氢酶活性[29]。
2.4 土壤微生物数量测定
采用平板计数法测算微生物数量:细菌采用牛肉膏蛋白胨平板,在30 ℃培养24 h计数;真菌采用马丁-孟加拉红平板,在25 ℃培养72 h计数;放线菌采用改良得高氏一号平板,在28 ℃培养96 h计数[30]。
2.5 数据分析
采用 Excel 2007、DPS v15.10进行整理数据、绘图及分析,采用中的Duncan新复极差法进行二因素统计分析及差异显著性分析,不同样地处理分别用LQ(碌曲县)、XH(夏河县)、HZ(合作市)表示,不同退化程度处理分别用L(轻度退化)、M(中度退化)、S(重度退化)表示,样地与退化程度二因素互作分别为LQL、LQM、LQS、XHL、XHM、XHS、HZL、HZM、HZS等9个处理;采用Pearson多变量相关进行相关性分析。
3 结果与分析
3.1 植被特性
如表2所示,草地高度受样地及退化程度的影响均较大,碌曲(LQ)草地平均高度最高,为16.58 cm,夏河(XH)草地平均高度最低,为12.70 cm;轻度退化草地(L)平均高度为21.06 cm,大于中度退化草地(M)平均高度(14.51 cm)及重度退化草地(S)高度(9.28 cm),差异显著(P<0.05);在不同样地及不同退化程度双重因素的影响下,各处理草地高度差异均显著(P<0.05),碌曲重度退化草地(LQS)高度较轻度退化草地(LQL)高度高出约10倍。不同样地的草地盖度和草产量部分差异显著(P<0.05),碌曲(LQ)草地盖度与其他两地差异均显著(P<0.05),为85.83%,夏河(XH)草地与合作草地盖度差异不显著(P<0.05),仅相差2%;碌曲(LQ)草产量与合作(HZ)草产量差异不显著(P<0.05),在2700 kg/hm2之上,夏河(XH)平均草产量最低,为2541.67 kg/hm2,与其他两地草产量差异显著(P<0.05)。不同退化程度的草地盖度及草产量差异均显著(P<0.05),轻度退化草地(L)盖度为94.33%,重度退化草地(S)的平均盖度仅为61.67%;轻度退化草地(L)的草产量为3708.33 kg/hm2,重度退化草地(S)的草产量仅为1883.33 kg/hm2,较轻度退化草地草产量降低了约2000 kg/hm2。在样地和退化程度的双因素影响下,各处理草地盖度及草产量差异均显著(P<0.05),草地盖度最高为碌曲轻度退化草地(LQL),达98%,最低为夏河重度退化草地(XHS),仅56%;草产量最高为碌曲轻度退化草地(LQL),为3825 kg/hm2,最低为夏河重度退化草地(XHS),仅1725 kg/hm2。
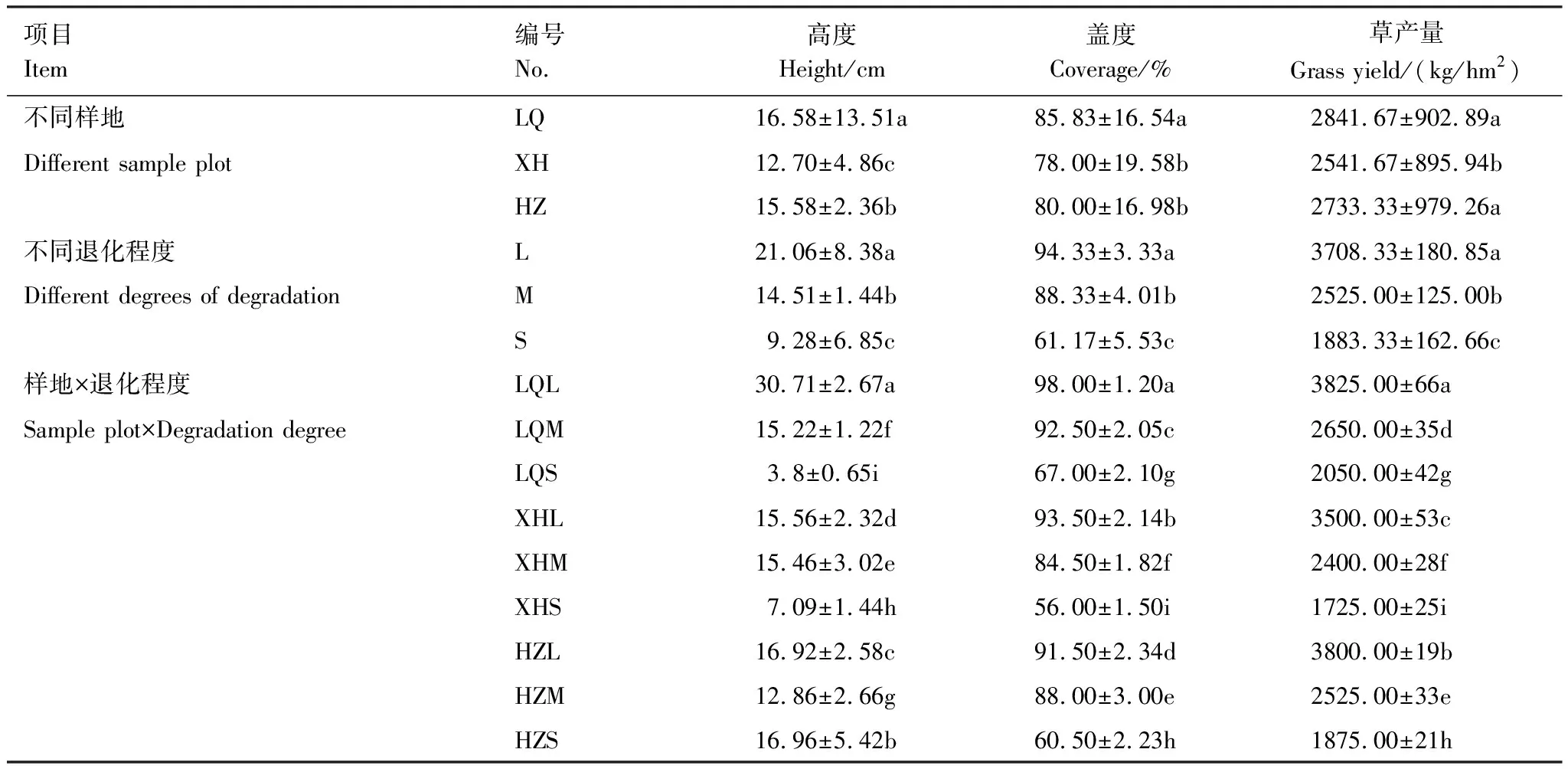
表2 不同样地不同程度退化程度高寒草甸植被特征
不同样地不同退化程度草地植被优势种差异较大,无明显规律性,碌曲县轻度退化草地(LQL)优势种为紫羊茅(Festucarubra)、甘青蒿(Artemisiatangutica)、四川嵩草(Kobresiasetschwanensis)、早熟禾(Poaannua),中度退化草地(LQM)优势种为细叶亚菊(Ajaniatenuifolia)、矮嵩草(Kobresiahumilis)、兰石草(Lanceatibetica)、火绒草(Leontopodiumleontopodioides),重度退化草地(LQS)优势种为鹅绒委陵菜(Potentillabifurca)、火绒草(Leontopodiumleontopodioides)、蚤辍(Arenariaserpyllifolia)、早熟禾(Poaannua);夏河县轻度退化草地(XHL)优势种为矮嵩草(Kobresiahumilis)、早熟禾(Poaannua)、披碱草(Elymusdahuricus),中度退化草地(XHM)优势种为莓叶萎陵菜(Potentillafragarioides)、乳浆大蓟(Euphorbiaesula)、雀麦(Bromusjaponicas),重度退化草地(XHS)的优势种为矮嵩草(Kobresiahumilis)、唐松草(Thalictrumaquilegiifolium)、披碱草(Elymusdahuricus);合作市轻度退化草地(HZL)优势种为鹅观草(Roegneriakamoji)、藨草(Sclrpuscernuus)、披碱草(Elymusdahuricus)、洽草(Koeleriamacrantha)、丝叶毛茛(Ranunculus tanguticus),中度退化草地(HZM)的优势种为细叶亚菊(Ajaniatenuifolia)、蚤缀(Arenariaserpyllifolia)、火绒草(Leontopodiumleontopodioides)、老鹳草(Geraniumwilfordii)、甘青蒿(Artemisiatangutica),重度退化草地(HZS)的优势种为早熟禾(Poaannua)、甘青蒿(Artemisiatangutica)、细叶亚菊(Ajaniatenuifolia)、白花地丁(Violapatrinii)、鹅绒萎陵菜(Potentillabifurca)。基于植被种类及各物种数量进行多样性分析发现(表3),合作草地(HZ)的各指数均高于其他样地,Simpson指数为0.92,与夏河草地(XH)植被Simpson指数差异显著(P<0.05),Shannon-Wiener指数为3.14,均匀度指数为0.82,Brillouin指数为4.37,McIntosh指数为0.76,与碌曲草地(LQ)和夏河草地(XH)的差异均显著(P<0.05);其中,夏河草地(XH)与碌曲草地(LQ)Brillouin指数和McIntosh指数差异不显著(P<0.05),合作草地(HZ)与碌曲草地(LQ)的Simpson指数差异不显著(P<0.05),由此可见,各样地间物种多样性无明显规律性。但不同退化程度的草地植被多样性指数规律性明显,Simpson指数、Shannon-Wiener指数、均匀度、Brillouin指数及McIntosh指数均随退化程度的加重而降低,且不同退化程度间各指数差异显著(P<0.05),如轻度退化草地(L)均匀度指数为0.89,重度退化草地(S)的均匀度指数降低至0.69,轻度退化草地(L)的McIntosh指数为0.81,重度退化草地(S)的McIntosh指数仅为0.64。在样地和不同退化程度的双重因素的影响下各指数均表现为合作轻度退化草地(HZL)的最高,无其他明显规律性。
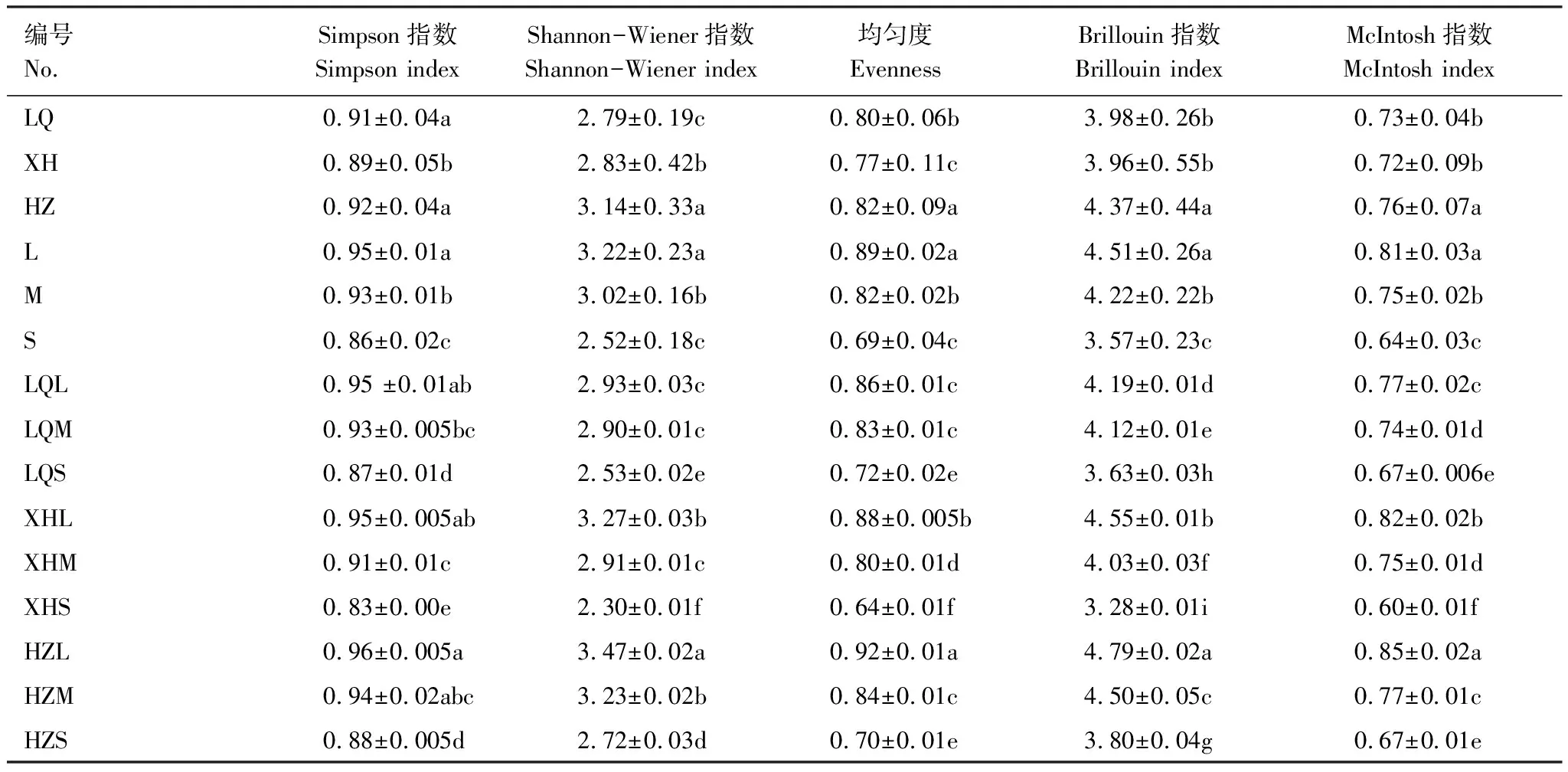
表3 不同样地不同程度退化程度高寒草甸植被物种多样性指数
3.2 土壤理化性质
如表4所示,不同样地(LQ、XH、HZ)草地全氮、全磷、全钾、土壤pH值、全盐、容重、孔隙度、颗粒组成均有所差异,且多数处理间差异显著(P<0.05),无明显规律性。不同退化程度(L、M、S)的各土壤理化性质指标变化规律性明显,随着退化程度的加重,除土壤砂粒变化不明显外,土壤全氮、全磷、全钾、孔隙度及粉粒呈降低趋势,土壤pH值、全盐、容重、黏粒呈升高趋势,且各退化程度间差异显著(P<0.05)。在不同样地和不同退化程度的双重因素影响下,合作市轻度退化草地(HZL)土壤全氮含量最高,为1.84 g/kg,且与中度(HZM)和重度退化草地(HZS)的含氮量差异显著(P<0.05)。土壤全磷含量则是夏河县轻度退化草地最高(XHL),为1.67 g/kg,且随退化程度加深下降程度最为明显,由轻度至重度降幅为0.67 g/kg,差异显著(P<0.05)。土壤全钾含量则是合作市轻度退化草地(HZL)最高,可达1.98 g/kg,且与其重度退化草地(HZS)全钾含量差异显著(P<0.05),由轻度至重度退化草地土壤全钾含量降幅达0.62 g/kg。土壤pH值除碌曲轻度退化草地(LQL)、合作轻度(HZL)及中度退化草地(HZM)呈弱酸性外,各样地其余不同退化程度草地土壤均呈碱性。此外,各处理草地全盐含量均较高,最高达2.27 ms/cm;土壤颗粒组成呈现出粉粒>砂粒>黏粒的特点,大部分退化草地土壤砂粒在30%以上。
3.3 土壤酶活性
如表5所示,碌曲草地(LQ)除过氧化氢酶活性与夏河草地(XH)相同之外,其他土壤酶活性均高于夏河草地(XH)和合作草地(HZ),且差异显著(P<0.05),夏河草地(XH)和合作草地(HZ)间酶活性的变化无显著规律性。但随着退化程度加重,脲酶活性和蔗糖酶活性显著降低(P<0.05),过氧化氢酶则呈现出先升高再降低的趋势,碱性磷酸酶活性亦呈现出随退化程度的加重而降低的趋势,但中度退化草地(M)与轻度退化草地(L)差异不显著(P<0.05)。在样地与退化的双重因素影响下,合作市重度退化草地(HZS)较其轻度退化草地(HZL)而言蔗糖酶活性降幅最大,达0.45 mg/g/24h。碌曲轻、中度退化草地(LQL、LQM)及夏河(XHL、XHM)土壤碱性磷酸酶活性差异不显著,重度退化草地(LQS、XHS)土壤碱性磷酸酶活性较其显著降低(P<0.05)。
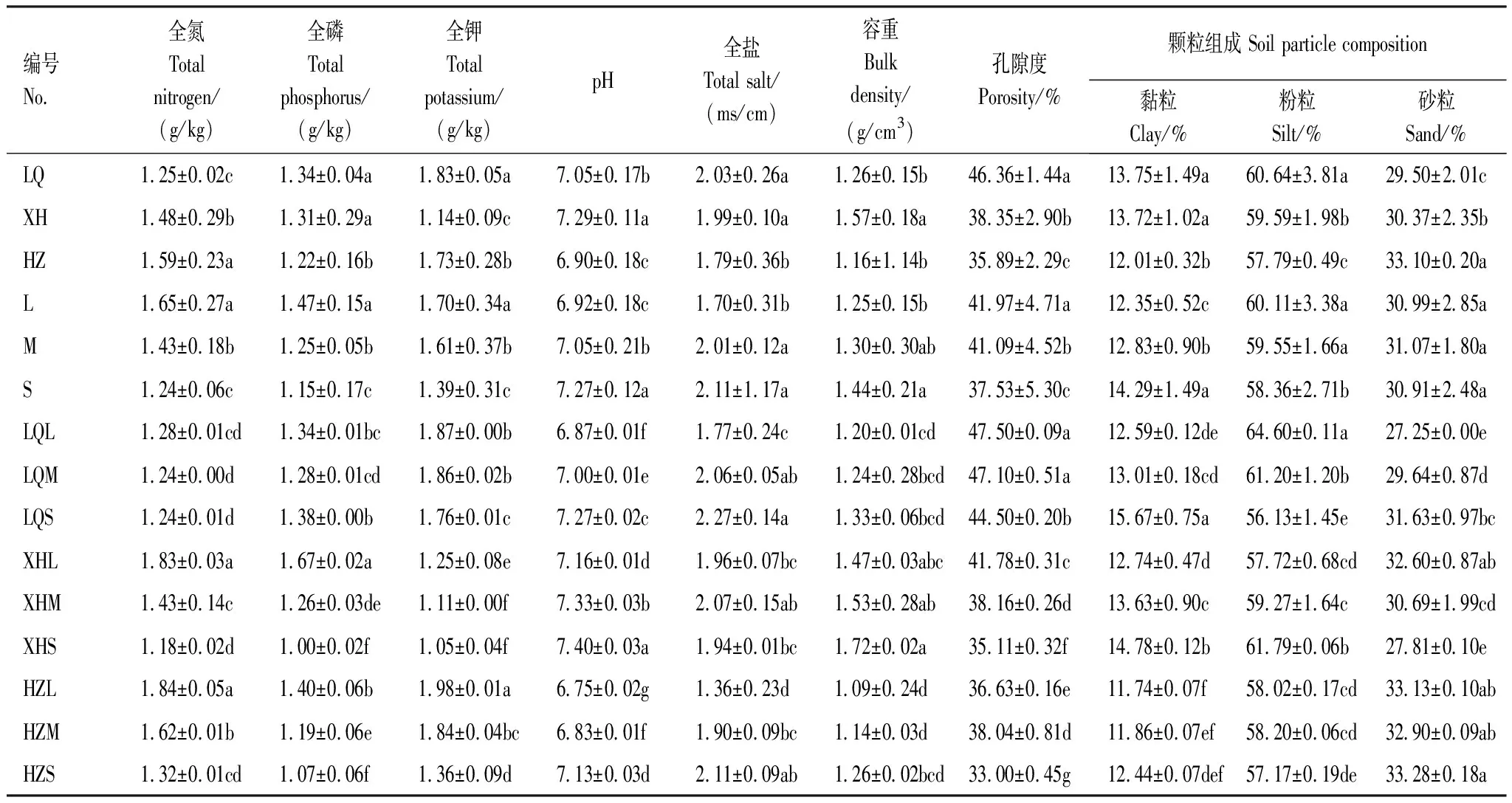
表4 不同样地不同退化程度高寒草甸土壤理化性质
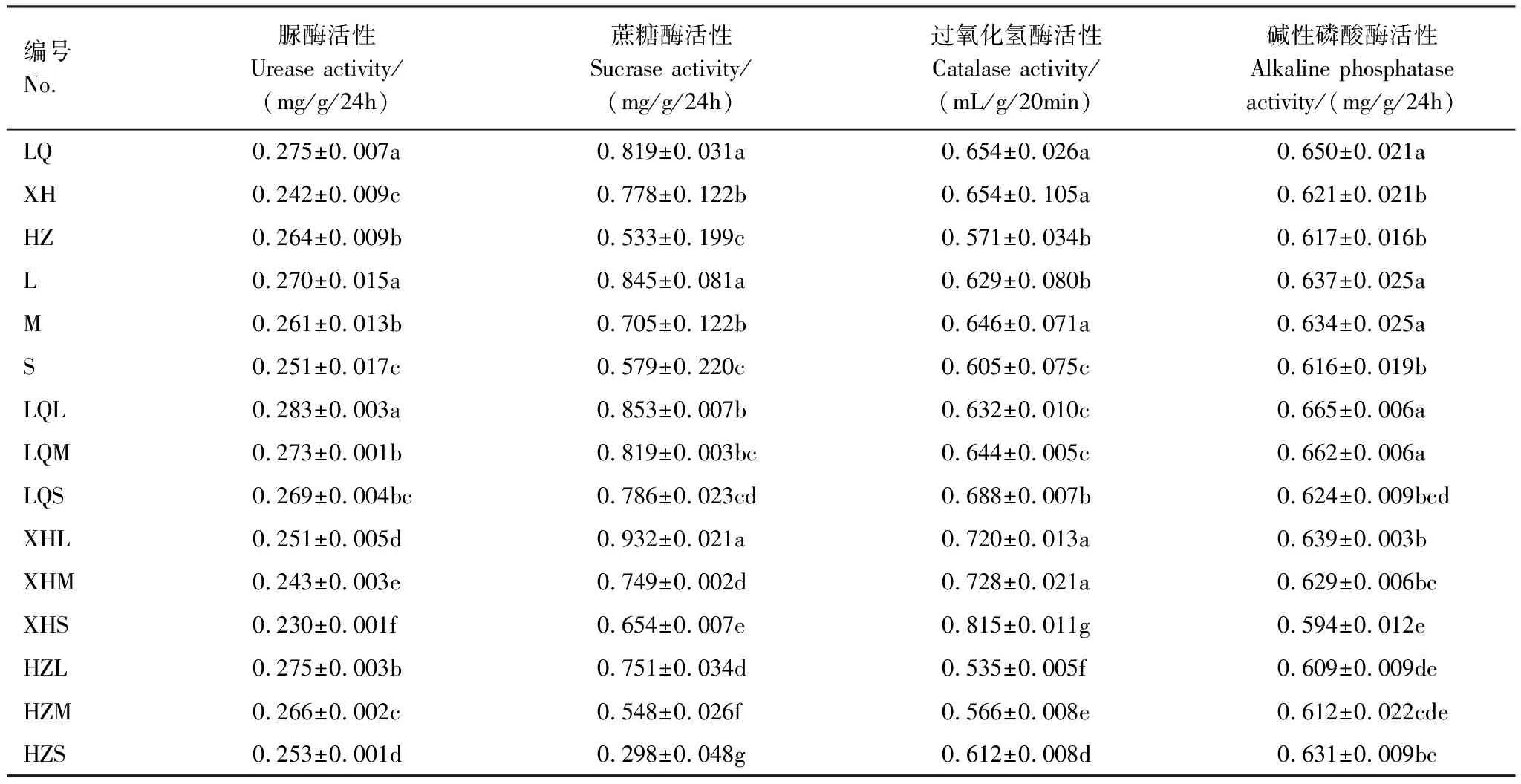
表5 不同样地不同退化程度高寒草甸土壤酶活性
3.4 土壤微生物数量
如表6所示,土壤微生物数量因样地不同均有所差异,夏河草地(XH)土壤细菌数量最高,达5.98×106cfu/g,合作草地(HZ)细菌数量最低,为2.56×106cfu/g,同时,合作草地(HZ)真菌数量及放线菌数量均最高,真菌数量为4.947×103cfu/g,放线菌数量为7.561×105cfu/g,与夏河草地(XH)和碌曲草地(LQ)的差异显著(P<0.05)。土壤微生物数量随着退化程度加重表现出明显的规律性,土壤细菌和放线菌数量均随草地退化程度加重而减少,轻度退化草地(L)土壤细菌数量为4.77×106cfu/g,重度退化草地(S)土壤细菌数量仅为1.33×106cfu/g;轻度退化草地(L)放线菌数量为8.391×105cfu/g,中度退化草地(M)放线菌数量为5.784×105cfu/g,重度退化草地(S)放线菌数量为3.063×105cfu/g;土壤真菌的数量则表现出相反的变化趋势,即随退化程度的加重土壤真菌数量增加,轻度退化草地(L)数量为3.304×103cfu/g,中度退化草地(M)数量为4.354×103cfu/g,重度退化草地(S)数量为5.758×103cfu/g,不同退化程度草地间各微生物数量差异均显著(P<0.05)。在样地和退化双重因素影响下,碌曲轻度退化草地(LQL)细菌数量及放线菌数量均最高,细菌数量达8.69×106cfu/g,放线菌数量为1.25×106cfu/g,合作重度退化草地(HZS)细菌数量最低,为1.04×106cfu/g。土壤真菌数量则为合作重度退化草地(HZS)最高,达7.343×103cfu/g。多个处理间土壤放线菌数量差异不显著(P<0.05)。

表6 不同样地不同退化程度高寒草甸土壤微生物数量
3.5 各因子间的相关性
通过对各因子的相关性分析发现(表7),各因子除砂粒和过氧化氢酶外,其他因子均与草地退化程度相关系数均大于0.92,且多在0.95之上,说明草地退化与植被及土壤特性的变化息息相关,其规律与前文分析结果一致。植被特性各因子间(高度、盖度、草产量及多样性指数)、除砂粒和过氧化氢酶外各土壤特性各因子间、土壤微生物各因子间,以及植被特性各因子与土壤特性(砂粒和过氧化氢酶除外)、土壤微生物各因子间相关性均较大,相关系数最低为0.82,多数因子间的相关指数均在0.90之上,0.99或0.98占比较大。植被特性各因子与土壤全氮、全磷、全钾、孔隙度、粉粒、脲酶活性、蔗糖酶活性及碱性磷酸酶活性均呈正相关关系,与土壤pH,全盐、容重及黏粒呈负相关关系,与土壤微生物细菌和放线菌数量呈正相关关系,与真菌数量呈负相关,相关系数在0.95以上,甚至达到1,即百分百完全相关的程度。土壤全氮、全磷、全钾、孔隙度、粉粒、脲酶活性、蔗糖酶活性及碱性磷酸酶活性与土壤细菌及放线菌数量亦呈正相关,与真菌数量呈负相关,土壤pH,全盐、容重及黏粒则与细菌和放线菌数量呈负相关,与真菌数量呈正相关,且相关系数最低为0.92,多为0.99甚至1。
4 讨论与结论
高寒草甸植被群落物种生长特征、优势种组成和结构等是揭示草地生态系统退化及演替规律的重要依据[13—15, 31],本研究中不同地区高寒草甸高度、盖度、草产量以及植被多样性指数均随退化程度加深显著降低,且呈现出牧草类植物的优势地位逐渐被毒杂草类代替的可能,这与李媛媛等[32]、邓燕等[33]研究结果一致;在该过程中,当退化由中度转为重度,降幅最大,与前人研究一致[34—35]。不同地区不同退化程度优势种组成差异较大,分析其原因可能一是不同地区不同退化程度高寒草甸气候、土壤、降水等生态因子不同所致,如土壤营养物质淋溶、挥发及青藏高原高寒草甸气候整体干热化变化趋势等影响,部分科属植株一旦死亡便很难再生[36];二是随着草地退化加剧,部分植被吸收利用营养物质的能力严重衰减,致使在较深退化程度下早熟禾、披碱草等禾本科优质牧草与嵩草属、委陵菜属杂草类植物同时占据优势地位;三是由于过度放牧,植被根部被啃食严重,部分植被生长受抑或死亡。
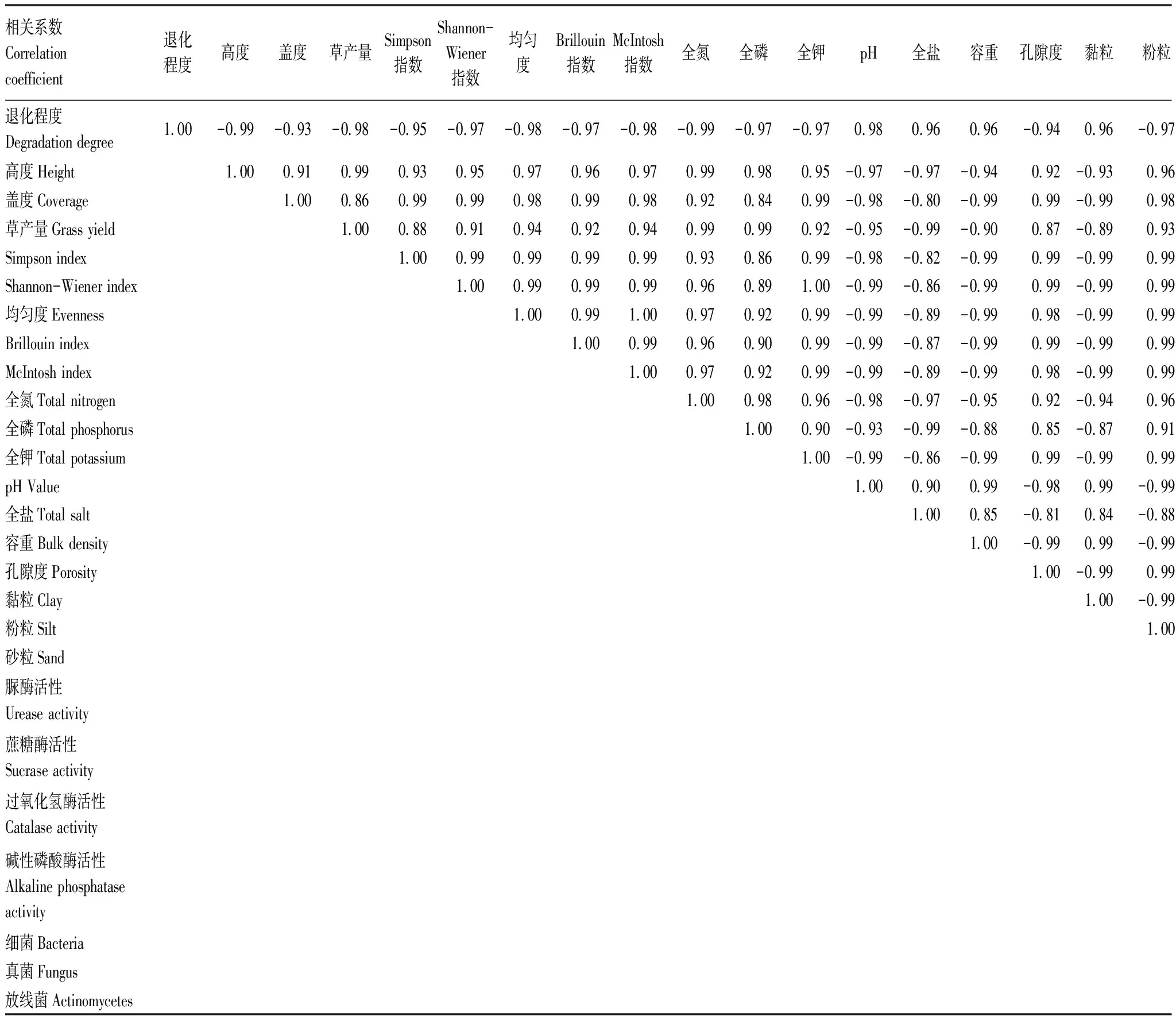
表7 甘南州高寒草甸植被及土壤特性各因子间的相关性
草地生态系统中,土壤理化性质能反映高寒草甸的退化状态和趋势[37]。本研究发现,随退化程度加深,土壤全氮、全磷、全钾、孔隙度及粉粒含量降低,土壤pH值、全盐、容重、黏粒含量升高,说明随着草地退化加剧,土壤肥力衰退、逐渐盐碱化、向荒漠化次生演替,与魏茂宏等[38]、杨馥铖等[39]研究结果一致。土壤酶则是植物根际微生态调节、运转的重要活性驱动因子,对土壤养分活化、迁移和供应等具有催化作用,土壤脲酶可催化尿素酰氨碳氮键水解并生成氨,蔗糖酶可将土壤中的多糖水解为小分子葡萄糖或果糖为植物及微生物提供营养,磷酸酶则对磷的转化起促进作用[40—42]。本研究发现,随高寒草甸退化加重,脲酶、蔗糖酶、碱性磷酸酶活性显著降低,表明在高寒草甸逆行演替进程中含氮、含碳、含磷营养物质转化受到严重抑制[43]。土壤微生物是土壤微生态系统的重要组成,是高寒草甸生态系统变化的敏感指标,其数量、组成波动直接反映了土壤健康状况,并间接反映了高寒草甸植被和土壤的演替方向[9,14]。本研究发现,随着草地退化加剧,土壤细菌和放线菌数量显著降低,真菌数量增多,与Sokol等[44]、王永宏等[45]研究结果一致。
综上,通过对甘南州不同退化程度高寒草甸植被及土壤特性的演化规律的研究发现,甘南州高寒草甸随着退化程度加重,植被高度、盖度、草产量、多样性指数降低,土壤全氮含量、全磷含量、全钾含量、孔隙度含量、粉粒占比、脲酶活性、蔗糖酶活性、碱性磷酸酶活性、细菌数量、放线菌数量降低,土壤pH值、全盐含量、容重、黏粒占比及真菌数量升高,土壤砂粒差异不显著(P<0.05),过氧化氢酶则先升高再降低。而“植被-土壤-微生物”这一整体演化动态是掌握高寒草甸演化规律及其退化修复的关键,本研究通过对植被、土壤及微生物各因子进行相关性分析发现,除个别因子外,各因子之间及各因子与草地退化程度之间相关性均较强,说明草地在退化过程中,“植被-土壤-微生物”三者之间相互依存、密不可分、协同演化。土壤作为联系植被与微生物的纽带,随着草地退化,其理化性质的演化趋势朝着不利于植被、微生物生存的方向进行,土壤中N、P、K等养分含量及有关能量转化、物质循环的酶活性逐渐降低,土壤向盐碱化、荒漠化演替。土壤微生物作为退化土地修复的变局者,其活性在生物地球化学循环中发挥着重要作用,是有机物形成、分解的重要介质及全球碳平衡的直接管控者[46],其群落结构失调,则反作用于植被及土壤理化性质及酶活性,致使退化进一步加剧。
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
科学技术创新(2022年30期)2022-10-21
课外生活·趣知识(2021年2期)2021-05-24
海外文摘·文学版(2021年3期)2021-05-19
学校教育研究(2021年24期)2021-03-28
西部资源(2018年3期)2018-11-06
时代金融(2017年23期)2017-09-13
中国市场(2017年7期)2017-04-01
中学生英语·外语教学与研究(2017年2期)2017-02-08
延河·绿色文学(2016年8期)2016-05-14
