
共栽培植物对蒜头果幼苗的共生效应
2022-11-04 13:54陈婉东王鹏飞王俊威
江西农业大学学报 2022年5期
陈婉东,王鹏飞,普 甜,王俊威,潘 悦,王 娟
(1.西南林业大学 林学院,云南 昆明 650224;2.西南地区生物多样性保育国家林业和草原局重点实验室,云南昆明 650224;3.西南林业大学 地理与生态旅游学院,云南 昆明 650224;4.西南林业大学 绿色发展研究院,云南 昆明 650224)
【研究意义】蒜头果(Malania oleifera)属铁青树科(Olacaceae)蒜头果属(Malania),是中国特有单属种植物[1-3],仅自然分布于云南东南部和广西西部的狭窄区域,被列为国家二级珍稀濒危保护树种[4-5]。蒜头果油中含有丰富的神经酸[6],能有效促进受损神经组织修复和再生,对心血管疾病有显著疗效[7]。同时蒜头果树形优美,叶子常绿光亮,可作为绿化树种;其主根发达、耐瘠薄,也是当地石漠化治理的优良树种。蒜头果生境狭窄,种群数量有限,蒜头果果实的价格逐年攀升。为缓解蒜头果的濒危现状和为其利用提供丰富的资源,蒜头果人工种植面积逐年扩大,但造林保存率普遍偏低[8]。因此提高人工培育蒜头果的成活率,打破制约蒜头果种植产业发展的技术瓶颈刻不容缓。【前人研究进展】作为半寄生植物的蒜头果[8],其通过根部吸器从其他植物获取养分,在其天然分布区内,蒜头果能与杉木、油杉、杨梅、合欢、紫茎泽兰和鬼针草等多种植物形成寄生关系[9]。李勇鹏等[8]研究表明,蒜头果对寄主植物的种类选择具有偏好性,与吊兰共栽培后,蒜头果幼苗的生物量、叶片数和吸器数量显著高于寄主植物为华山松、洋芋和白蒿的处理。另有研究表明,蒜头果通过根部吸器获取寄主植物的营养物质,通过切片观察发现蒜头果的吸器连接寄主根并穿透其根的表皮和皮层直达维管束,该维管束鞘穿透寄主根皮层后分化为吸盘与寄主根木质部相连,以获取寄主植物的营养物质[9]。在此过程中,寄主植物体内的营养物质被摄取,导致寄主植物的生长发育受到抑制,死亡率升高[10-11]。【本研究切入点】目前,虽已有蒜头果半寄生特性的报道,但蒜头果与共栽培植物的共生效应研究仍十分有限。【拟解决的关键问题】研究以蒜头果、吊兰、艾蒿和油茶为研究对象,通过选取与蒜头果同一生境的油茶(Camellia oleifera)、艾蒿(Artemisia argyi)以及根系发达、具肉质根的吊兰(Chlorophytum comosum‘Variegatum’)为共栽培植物,分别测定蒜头果与共生植物的生长发育指标及生理指标,并通过显微观察蒜头果与共栽培植物形成的吸器结构,筛选蒜头果适宜的寄主植物种类,为提升蒜头果高效栽培技术提供参考依据和数据支持。
1 材料与方法
1.1 试验材料与地点
2019年10月从云南省文山州富宁县采集新鲜蒜头果及油茶种子,带回实验室用50%多菌灵可湿性粉剂浸泡10 min 后,清水冲洗干净,通风干燥处阴干,置于西南林业大学树木园温室的沙池中进行层积催芽。2020年3月将萌发的蒜头果及油茶种子种植于底径15 cm,高20 cm的塑料花盆,基质选用体积比为6∶2∶2的红土、椰糠和珍珠岩混配,定期浇水管护。吊兰、艾蒿采自西南林业大学树木园,将茎段剪为长度5 cm左右,用生根粉、多菌灵浸泡消毒后扦插在蒜头果周围。试验在西南林业大学树木园温室中进行,位于东经102°46′,北纬25°03′,海拔1 964 m。
1.2 试验设计
2020年8月蒜头果植株长至15 cm左右,选择长势一致的幼苗,分别与吊兰(栽培品种金边吊兰)、艾蒿、油茶共同栽培。设置4 个处理:(1)SD 处理:蒜头果与吊兰共栽培;(2)SH 处理:蒜头果与艾蒿共栽培;(3)SY处理:蒜头果与油茶共栽培;(4)S处理:蒜头果单株种植,其中共栽培的处理植株间相距3 cm,每处理重复20次。
1.3 指标测定
1.3.1 生长发育观察和生物量测定每月定期测定蒜头果及共栽培植物的株高、叶片数和分枝数。其中株高用钢卷尺测量植株茎基部到茎尖的距离,叶片和分枝数为整株幼苗的所有的叶片和分枝数量。2021 年5 月收获植株,每处理随机选取10 盆植株,将蒜头果及共栽培植物从盆中取出,洗净植物根部附着的泥土,使水分自然风干,并称量幼苗地上、地下部分的鲜重。其中,蒜头果地下部分分为主根,一级、二级侧根及须根3 部分。将植株的地上、地下部分分别在鼓风干燥箱105 ℃杀青30 min 后,65 ℃烘干至恒重后通过电子天平称量,分别称取地上、地下部分后,计算寄主利用效率。
寄主利用效率=寄生植物地上部干重/寄主植物地上部干重× 100%(1)
1.3.2 蒜头果吸器的形态及数量观察将各处理剩余10盆植株取出,清水洗净根部附着泥土,统计蒜头果主根、侧根和须根上的自吸器数量以及与寄主形成的寄生吸器数量,并计算蒜头果与共栽培植物的结合率,其中:
结合率=与蒜头果吸器相连植株数/总植株数× 100%(2)
将采集得到的吸器置于FAA 固定液(70%乙醇∶冰醋酸∶甲醛=90∶5∶5)中保存[12],使用LEICA M205 FA 体视镜观察吸器的外部形态,并采用常规石蜡切片的制片技术,在奥林巴斯BX53荧光显微镜下观察吸器的显微结构并拍照[13]。
1.3.3 生理指标测定对各处理剩余的10 盆植株,每株取茎尖以下第4~6 片新鲜叶片,擦洗干净,各指标分别称取0.1 g 鲜叶,测定植株叶片的叶绿素含量,可溶性糖含量及可溶性蛋白含量,每项指标9 次重复。其中叶绿素含量采用80%的丙酮提取,分光光度计法测定[14];可溶性糖含量采用蒽酮比色法测定[14];可溶性蛋白含量采用考马斯亮蓝G-250染色法测定[14]。
1.3.4 数据统计与分析使用SPSS 25.0 分别对蒜头果幼苗及共栽培植物的生长生理数据进行单因素方差分析(one-way ANOVA),采用Duncan法进行多重比较,使用originPro 2021绘图。
2 结果与分析
2.1 共栽培植物对蒜头果生长及生物量的影响
由图1a 可以看出,蒜头果株高净增量不同处理之间差异均达显著水平(P<0.05)。各处理中,以SY处理的株高净增量最大(5.84 cm),S处理次之(4.35 cm),SD处理最小(3.40 cm)。
由图1b可知,蒜头果叶片的净增量不同处理之间差异显著,与不同植物共栽培的蒜头果的叶片的净增量与株高呈现相同的趋势,各处理中,以SY 处理的叶片净增量最大(11.87片),S处理次之(10.00片),SD处理最小(4.63片)。
由图1c 可知,蒜头果分枝数净增量在不同处理之间以SY 处理最多,与S 处理差异不显著,但与SH处理、SD处理之间差异达显著水平。
如图1d 所示,蒜头果不同处理间的总生物量的差异均达显著水平,其中SY 处理的生物量最大(13.36 g),S 处理(12.09 g)与SH 处理(11.77 g)次之,SD 处理最低(10.56 g)。SY 处理下的地上部分干重显著高于SD 和SH 处理,与S处理差异不显著(P>0.05)。SY 与SH 处理下的蒜头果幼苗地下部分干重显著高于SD 处理。由此表明,蒜头果与油茶共同栽培下,地上、地下及总生物量均高于其它处理,表明对蒜头果幼苗的生物量积累具有明显促进效果。
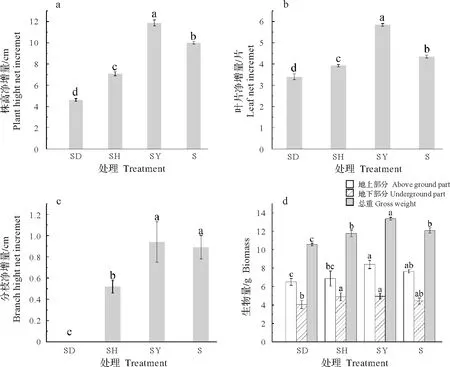
图1 共栽培植物对蒜头果生长及生物量的影响Fig.1 Effect of host plants on growth and biomass of M.oleifera seedlings
综上所述,蒜头果与油茶共栽培时,可以显著促进其株高生长、叶片数以及生物量积累的增加,但对其分枝数无显著影响。
2.2 共栽培植物对蒜头果根部吸器的影响
如表1所示,蒜头果根部自吸器分布数量由大到小依次为须根、侧根和主根,在蒜头果主根上,SD处理自吸器数量显著高于S和SH处理,蒜头果侧根上SH处理的自吸器数量显著高于其余处理,SY处理的须根及总自吸器的数量最多,与其它处理均差异显著,而S处理总自吸器数量最少。
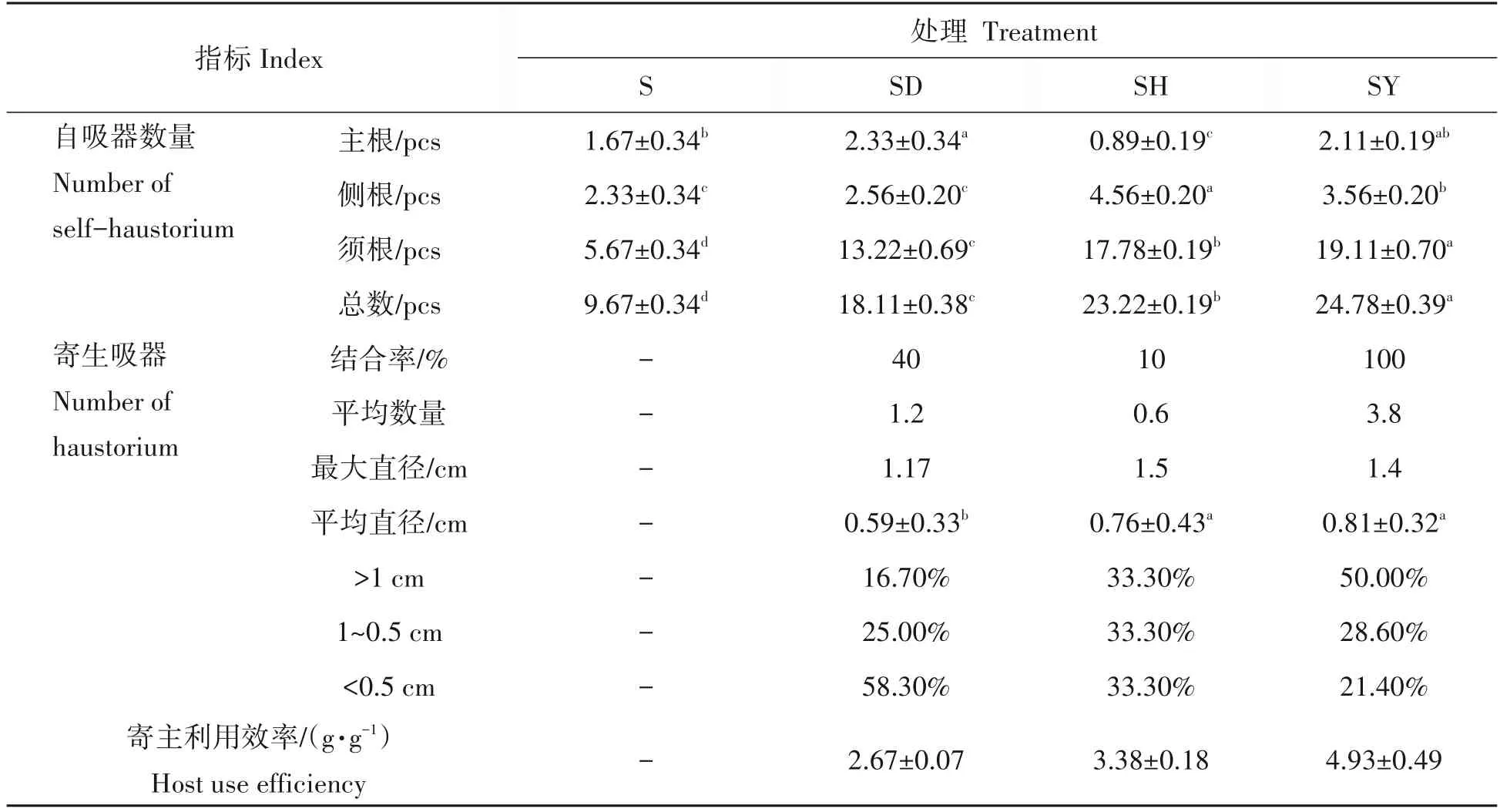
表1 蒜头果根系吸器及寄生情况Tab.1 Effects on root haustorium and parasitism of M.oleifera
蒜头果与不同共栽培植物的根系之间均有寄生吸器产生,但与不同共栽培植物的结合率不同,蒜头果与油茶的结合率最高为100%,且吸器数量最多,与吊兰(40%)和艾蒿(10%)的结合率较低,吸器数量也较少;蒜头果与不同植物根部形成的寄生吸器直径存在差异,SY 和SH 处理的吸器平均直径显著大于SD处理,SY、SH处理的寄生吸器最大直径大于SD处理。将寄生吸器按照直径大小划分等级,SY处理吸器直径大于1 cm 的占比最多。此外,SY 处理的寄主利用效率最高,其次是SH 处理,SD 处理的寄主利用效率最低。结果表明,蒜头果与不同植物共栽培均可促进蒜头果根部自吸器数量的增加,并且蒜头果与油茶结合率与寄主利用效率最高。
由图2可以看出,蒜头果与不同共栽培植物根部所形成的不同大小的寄生吸器在形态上的差异不明显,蒜头果形成的寄生吸器表面均呈不规则凸起。图3显示,蒜头果与不同共栽培植物寄生吸器的横切剖面结构有明显差异。蒜头果的寄生吸器与油茶根的结合紧密,其吸管与油茶的维管束相连,吸管前端如双手托在维管柱的外层细胞上,油茶的根皮组织被吸管破坏,寄生吸器处根的表皮细胞消失,吸管穿过根的皮层,改变了寄主根皮层组织的结构(图3a,3b);蒜头果的寄生吸器完全包裹了艾蒿根,但寄主根的各部分组织结构完整,寄生吸器处的寄主根表皮细胞和皮层细胞完整而未受到影响,吸管未穿过艾蒿根的表皮和皮层,寄生吸器与根结合不紧密,可轻易分离(图3c,3d);蒜头果的吸器侵入到吊兰根部,产生的吸管刺穿吊兰根部的皮层与中柱相连,但吸器干瘪,吸器的内部结构受损(图3e,3f)。

图2 蒜头果与不同植物形成的吸器Fig.2 Haustoria of M.oleifera with different plants

图3 蒜头果与不同植物形成吸器的横切图Fig.3 Transverse section of haustoriums of M.oleifera attached to different plants roots
由图4 蒜头果的寄主吸器与寄主根的显微观察结果可知,蒜头果与油茶根所形成的吸器有少量的维管组织分化,维管组织被周围的深色的薄壁细胞包围,寄生吸器与油茶根表皮的接触处也可见维管组织的分化,寄生吸器有明显的吸管,吸管与寄主根部相接触的细胞呈指状(图4a);蒜头果与艾蒿形成的寄生吸器未见维管组织的分化,仅见薄壁细胞(图4b);蒜头果与吊兰形成的寄生吸器可见少量维管组织分化和薄壁细胞;寄生吸器与根表皮接触处向寄主根内侵入,但根表皮相接处未见明显的维管组织分化(图4c)。

图4 蒜头果与共栽培植物形成吸器的石蜡切片横切Fig.4 Paraffin and transverse section of hemiparasitic structure of M.oleifera attached to different plants roots
2.3 不同处理对蒜头果的生理指标的影响
2.3.1 对叶绿素含量的影响如图5所示,各处理的蒜头果叶绿素a、b和总叶绿素含量表现出较为一致的趋势,其中SY 处理的叶绿素含量显著高于其他处理,不同处理之间叶绿素含量从高到低的顺序依次为SY、SH、S和SD。
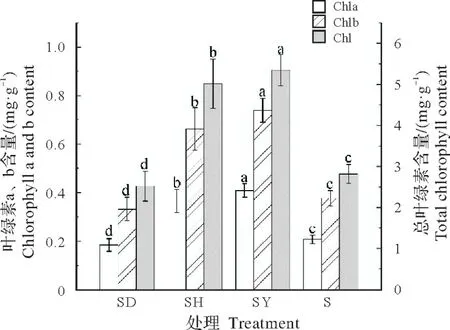
图5 不同共栽培植物对蒜头果叶绿素含量的影响Fig.5 Effects of different host plants on chlorophyll content of M.oleifera
2.3.2 对可溶性糖含量的影响由图6 可知,SD、SY 和S 处理下的蒜头果可溶性糖含量较SH 处理的可溶性糖含量显著下降,分别减少15.09%、7.12%和5.11%;单株种植较SY 处理下的蒜头果可溶性糖含量无显著差异,SD处理的可溶性糖含量显著低于其余处理。结果表明,与单独种植的蒜头果相比,与艾蒿共栽培可促进蒜头果幼苗叶片的可溶性糖含量增加,而与吊兰共栽培,会导致其可溶性糖含量下降。
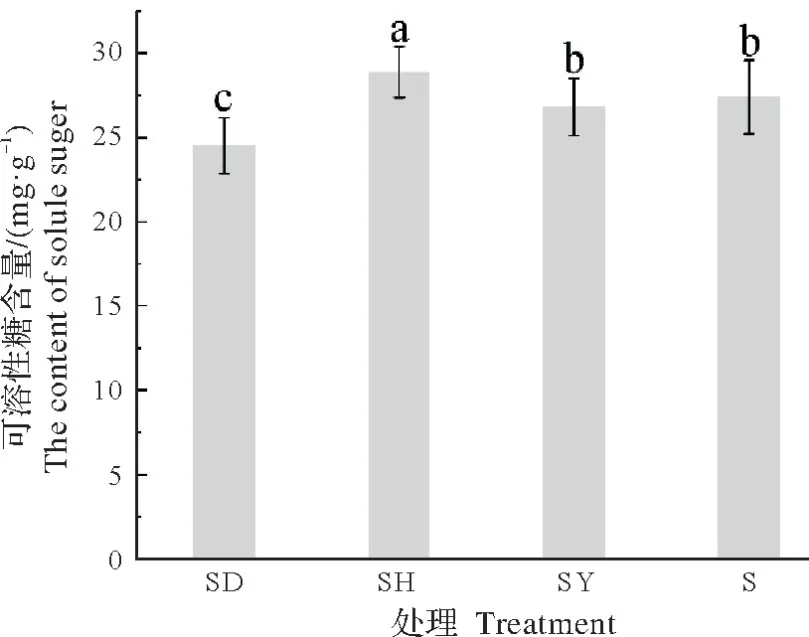
图6 不同共栽培植物对蒜头果可溶性糖含量的影响Fig.6 Effects of different host plants on soluble sugar content of M.oleifera
2.3.3 对可溶性蛋白含量的影响由图7 可知,不同处理蒜头果可溶性蛋白含量从高到低分别为SD、S、SY、SH,其中SD 处理的蒜头果的可溶性蛋白含量最高,达到了46.070 mg/g,显著高于其余处理。SY 和SH处理的蒜头果可溶性蛋白含量较S 处理显著降低,表明与单独种植的蒜头果相比,与吊兰共栽培会促进蒜头果的可溶性蛋白含量增高,而与油茶和艾蒿共栽培会导致其可溶性蛋白含量降低。
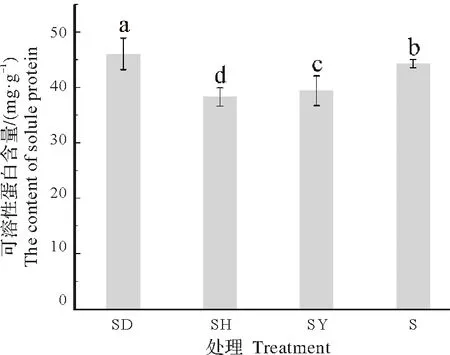
图7 不同共栽培植物对蒜头果可溶性蛋白含量的影响Fig.7 Effects of different host plants on soluble protein content of M.oleifera
3 结论与讨论
蒜头果是根部半寄生植物,叶片中含有叶绿素,自身可以进行光合作用合成碳水化合物自供,并不完全依赖于寄主植物,然而随着培育时间的延长,其根部对土壤中水分和养分的吸收能力有限,导致在没有寄主植物的情况下,蒜头果生长逐渐表现出营养缺乏、萎蔫死亡等症状[15]。同时,不同的植物体内养分存在差异,作为共栽培植物时对寄生植物的养分供应也存在差异[16]。因此筛选出适宜蒜头果共同栽培的植物,对深入探讨蒜头果的半寄生特性以及提升蒜头果高效栽培技术十分必要。
3.1 不同共栽培植物对蒜头果的生长的影响
相关研究表明半寄生植物与优良寄主共同栽培时,半寄生植物的生长发育将得到促进,常表现为株高快速增长[17]。本研究中,蒜头果与油茶共栽培后,株高、叶片、分支净增量、生物量以及寄主利用效率显著高于其余处理,这与Radomiljac 等[18]研究的印度檀香与优良寄主共同栽培的结果相似,优良寄主能更有效的促进印度檀香幼苗的生长。然而,当蒜头果与艾蒿和吊兰共栽培后,其株高、叶片、分支净增量均显著低于单株种植的蒜头果。与艾蒿共栽培的处理,生物量与单株种植的蒜头果无显著差异,而与吊兰共栽培的处理,生物量显著低于单株种植的蒜头果。综上所述,油茶共栽培对蒜头果的生长发育以及生物量积累的促进效果最为显著,而蒜头果与艾蒿、吊兰共栽培对其生长发育无明显的促进作用。
3.2 不同共栽培植物对蒜头果吸器的影响
相关研究表明,寄生植物与优良寄主植物共同栽培下,吸器直径较大,并且与寄主植物根部结合紧密[17]。本研究显示蒜头果与油茶共栽培时,蒜头果的寄生吸器数量最多,结合率最高,形态上寄生吸器饱满地包裹住油茶的根,且在相同栽培时间和条件下,吸器可发育分化形成吸管,表明了蒜头果可从油茶获得生长需要的营养物质,其利用效率最高。因此,蒜头果与油茶共栽培后,寄生吸器的发生及数量,在一定程度上反映出对油茶作为寄主的适应性和接受度。而不良寄主植物与半寄生植物所形成的吸器往往吸附不牢固且数量少,吸器发育不良或干瘪[19]。本研究中,蒜头果与艾蒿的根部所形成的吸器结合较不紧密,可轻易地将蒜头果吸器从艾蒿的根部分离,艾蒿与蒜头果所产生寄生吸器数量较少,结合率最低,吸器发育完全后仍未与艾蒿根部相连;蒜头果与吊兰所形成的吸器发育正常,数量较少,结合程度一般,虽有吸管产生,但吸器发育后期干瘪且内部结构被破坏。与此同时,蒜头果与不同植物共栽培后,吸器数量均显著高于单株种植的蒜头果吸器数量,表明寄主的刺激对寄生植物根部吸器的产生有促进作用;此外,相关研究表明,寄生植物可通过识别寄主植物分泌的一类化学信号物质诱导吸器的产生[20-21]。综上所述,油茶与蒜头果所形成的寄生吸器数量最多,结合率最高且最为紧密,蒜头果对油茶的利用效率最高,说明蒜头果对油茶具有明显寄主偏好。
3.3 不同共栽培植物对蒜头果生理的影响
叶绿素是植物进行光合作用必要的物质基础,叶绿素的含量在一定程度上可以反映植物的光合作用能力[22],对于植物的生长发育可产生直接的影响[23]。本研究中,与油茶共栽培的蒜头果叶片叶绿素含量显著高于其余处理,推测当蒜头果与油茶建立寄生关系后,蒜头果能从寄主体内获取养分供自身生长,同时通过提高叶绿素含量,增强自身的光合效率,这与李应兰[17]的研究结果一致,半寄生植物与优良寄主共同栽培时,植株叶片颜色较深,呈现具有光泽的浓绿色。此外,李勇鹏等[8]从华山松、洋芋、吊兰和白蒿的共栽培试验中发现,吊兰作为寄主植物蒜头果的吸器数量及各项生长指标较好,与本研究结果不一致的原因可能是吊兰的品种原因,试验中选用的金边吊兰与普通吊兰的叶绿素含量不同,能提供的养分含量可能存在差异;此外,试验中选择的基质以及容器的不同,可能也会造成结果产生较大的差异[24-25]。
可溶性糖是植物光合作用的直接产物,其含量变化既反映了碳水化合物的合成、运输情况,一定程度也反映出植物的物质积累情况[26-27]。而可溶性蛋白作为植物体内重要的渗透调节物质对植物的生长发育具有重要意义[26,28]。当植物受到轻度胁迫时,可溶性糖及可溶性蛋白的含量还会增加以应对不良情况[29]。本研究中,与艾蒿共栽培的蒜头果叶片可溶性糖含量显著高于单株种植的蒜头果,与吊兰共栽培的蒜头果叶片可溶性蛋白含量显著高于其余处理,推测是共栽培植物可利用的资源有限,导致植物间存在空间争夺从而产生了植物间轻度胁迫,促使其可溶性糖和蛋白含量升高。此外,有研究表明,半寄生植物偏爱寄生于含氮量较高的豆科及禾本科植物,原因是固氮植物能为其提供易于吸收的氮源[30],而蒜头果对固氮植物是否也具有偏好性还有待进一步研究证实。由此表明,油茶作为蒜头果的共栽培植物,对蒜头果的光合作用具有明显的促进作用,并且不会与蒜头果形成明显的竞争关系使蒜头果处于胁迫条件,再次表明油茶适宜与蒜头果共同栽培。
基于上述研究结果,表明油茶是蒜头果适宜的寄主植物种类之一,该物种与蒜头果位于同一生境,与蒜头果同为油料作物,共栽培时也能获得一定的经济效益。下一步将深入探究油茶与蒜头果寄生过程中吸器发生发育过程的解剖形态及生理生化响应,吸器与寄主植物根结合过程中释放的化学信号物质,并将继续扩大蒜头果寄主植物的筛选范围,为筛选蒜头果优良寄主植物,提升蒜头果人工培育技术提供数据支持。
致谢:云南省云岭产业技术领军人才项目(2018-212)和西南林业大学校级科研项目(112103,111930)同时对本研究给予了资助,谨致谢意!
猜你喜欢
基层中医药(2020年9期)2020-11-27
东方少年·阅读与作文(2019年2期)2019-06-11
现代园艺(2018年2期)2018-03-15
江苏农业科学(2017年7期)2017-05-23
作文评点报·中考版(2017年4期)2017-03-06
山东农业科学(2014年9期)2015-01-07
小学生导刊(中年级)(2009年2期)2009-03-23
