
基于混合线性模型的高州油茶杂交亲和性遗传评估Ⅰ
2022-12-05 06:53张应中张坤昌廖柏勇李永泉张盟雨李铭荃徐煲铧吕宇宙梁瑞友
经济林研究 2022年3期
张应中,张坤昌,廖柏勇,李永泉,张盟雨,李铭荃,王 溢,徐煲铧,郭 微,吕宇宙,梁瑞友
(1.广东省林业科学研究院 广东省森林培育与保护利用重点实验室,广东 广州 510520;2.仲恺农业工程学院,广东 广州 510225;3.韶关市曲江区国有小坑林场,广东 韶关 512600)
油茶Camellia oleifera是属于山茶科Theaceae山茶属Camellia油茶组Sect.C.oleifera的一类植物。油茶在中国栽培历史悠久,与油棕、油橄榄和椰子并称为“世界四大木本油料植物”[1-3]。以油茶种子为材料所提取的茶油含90%以上不饱和脂肪酸,营养比例合理,是我国传统的优质食用油。茶油色泽清,味道香,花生酸含量极低,易于消化,且能促进脂溶性维生素吸收,长期食用对人体健康非常有益。
高州油茶,又名越南油茶[4],是我国著名的油茶地理小种之一,主要分布在广东西南部、广西南部、海南北部以及中南半岛,尤以广东的高州和阳春及广西的陆川、容县、玉林和宁明等地资源丰富,是南缘油茶的重要组成部分[5-6]。高州油茶树形高大、枝叶茂密、果实中等偏大、单株产量高,是山茶亚属中优良的油茶树种[7-9],其早期品系资源有大石冲、福芳、大双、白石坡、罗平、平山、合水口等[10],但产量普遍偏低,因此提高高州油茶产量成为其资源利用的重要研究内容。油茶和其他林木品种的遗传改良一般采用农家优良品种选育、诱变育种、杂交育种、转基因或基因编辑等方法[11-19],杂交育种一直是高州油茶品种改良的重要方法之一。
油茶为专性异交授粉植物,在大面积种植单一无性系时,需要配置高亲和性的授粉树,以提高结实率[20]。染色体倍性变异、花期不遇、杂交不亲和、授粉昆虫特异性等因素常导致油茶杂交授粉失败[21]。在早期优树选择的基础上,大量开展高州油茶种内和种间杂交选育,检测优良品系的种间可配性和种内亲和性,估算杂交亲和性的一般配合力和特殊配合力,可提高优良品系的杂交选育成功率,选配高亲和性的杂交亲本。
本研究中选取高州油茶4 个优树无性系以及小果油茶C.meiocarpa、普通油茶和香花糙果茶C.suaveolens各1 个无性系,开展5×7 的双列杂交试验,探讨不同油茶树种种间杂交可配性和高州油茶无性系间的杂交亲和力等遗传变异特征,旨在为广泛开展高州油茶杂交选育提供参考,从而进一步提高南缘油茶的产量和品质。
1 材料与方法
1.1 试验地概况
试验地位于广东省韶关市曲江区国有小坑林场(113°35′E,24°15′N)。地处南岭山脉大庾岭南缘,属亚热带气候区,平均最高气温为34.0 ℃,平均最低气温10.0 ℃。林地为红壤,适宜油茶生长。
1.2 试验材料
具体参试油茶无性系见表1。高州油茶的4 个嫁接无性系优树中,3 个来自广东高州,1 个来自海南琼东;小果油茶、普通油茶和香花糙果茶各1 个无性系,分别来自广东兴宁、广西岑溪和广东高州。授粉母树共40 株,于2015年3月5日定植。
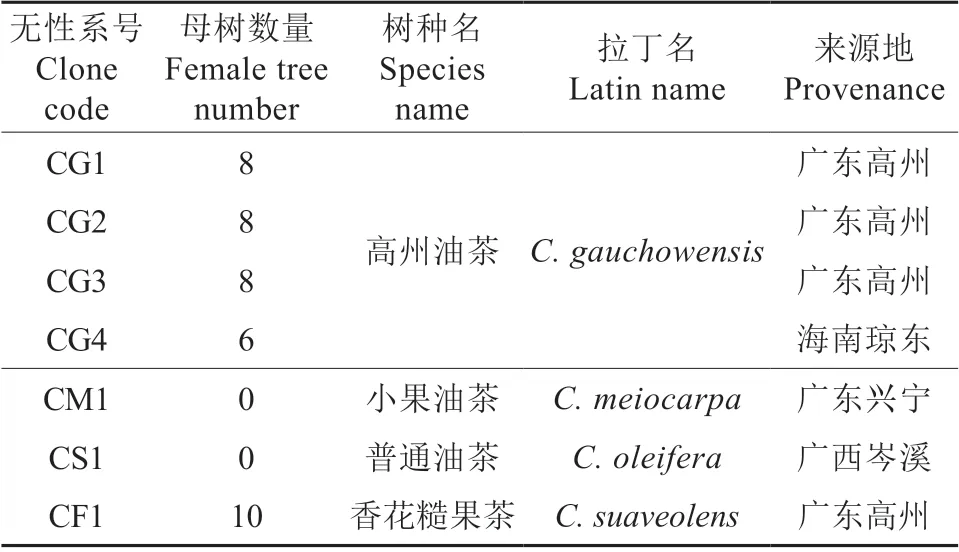
表1 参试油茶无性系†Table 1 The tested clones of C.oleifera
1.3 染色体倍性估测
以六倍体‘华鑫’、六倍体‘华硕’和八倍体‘岑软3 号’(CS1)为染色体倍性观测样品的对照[22-23]。2022年3月6日,采集对照和试验样品的嫩枝,选取新鲜幼嫩、无病虫害的叶片,使用Sysmex CyFlow®染色体倍性分析仪(德国)分析各亲本染色体倍性。按照CyStain® UV Precise P Automate试剂盒流程操作:取400 μL 提取缓冲液放入培养皿中,将样品放入提取缓冲液中裂解,用刀片垂直把样品切碎,持续30 ~60 s,用30 μm 的滤膜过滤样品,加入1 600 μL DAPI 染色溶液,上机检测。
X=(S/S0)×X0。
式中:X表示待测样品染色体倍性;S表示待测样品DNA 荧光面积均值;S0表示对照样品DNA 荧光面积均值;X0表示对照染色体倍性。
1.4 杂交试验设计
7 个参试无性系中,因CM1 和CS1 盛花期已过,仅作为父本,其他无性系间互为父母本。因为油茶自交成功率低[24],本试验中不统计自交。为方便统计和区别组合数量,每株母树仅接受1个父本的花粉,合计30 个杂交配对组合(表2)。

表2 油茶杂交试验设计†Table 2 Hybridization experiment design for the tested varieties of camellia-oil species
花粉处理方法:用硫酸纸袋采集花药,采用50 W 的白炽灯,在30 cm×45 cm×50 cm 的半密封纸盒中烘烤至花粉自由散出,分装3 ~4 瓶备用。授粉方法:分别在2019年12月19—21日和2019年12月25—27日(共6 d)进行授粉,气温为8 ~16 ℃,选择合适花苞,去雄、授粉、套袋,统计总授粉花朵数量。
1.5 数据统计和分析
拆除授粉套袋,检验并记录授粉情况。授粉花朵发育正常记录为1,败育或者无授粉花朵记录为0。从授粉后第6 天开始连续3 d(2020年1月3—5日),统计授粉花朵的保存数量和保存率。从授粉后第103天开始连续3 d(2020年3月10—12日),统计坐果数量和坐果率。保存率为授粉花朵保存数量占授粉花朵总数量的比例;坐果数量为授粉花朵保存数量减去发育失败的子房数量;坐果率为坐果数量占授粉花朵数量的比例。
使用Excel 2010 软件统计授粉花朵的保存率和坐果率;使用SPSS 22.0 软件开展方差分析、描述性统计量和多重比较分析。针对授粉花朵的保存数量和坐果数量,使用Asreml-R 软件[25]拟合混合线性模型[26](亲本模型I),参考林木非完全双列杂交的全同胞亲本模型[27-28]估算配合力,并采用R 软件绘制图表。
亲本模型Ⅰ:yijk=μ+Rijk+Mj+Dk+Sjk+e。
式中:yijk是第i次杂交第j母本和第k父本的亲和性观测值,μ是整体亲和性总体均值的固定效应,Rijk是第j母本和第k父本的第i次重复杂交的固定效应,Mj是第j母本的一般配合力(GCA)随机效应值,Dk是第k父本的一般配合力随机效应值,Sjk是第j母本和第k父本的特殊配合力(SCA)的随机效应值,e是随机误差。
2 结果与分析
2.1 参试样品染色体倍性
参试油茶样品流式细胞峰值分布结果如图1所示,参试油茶样品染色体倍性估测参数见表3。由表3可知,在3 个对照品种互为对照的情况下,‘华鑫’的染色体倍性平均预估值为6.38,‘华硕’为6.02,CS1 为7.51。参试样品中,CM1 为六倍体,其他均为十倍体。
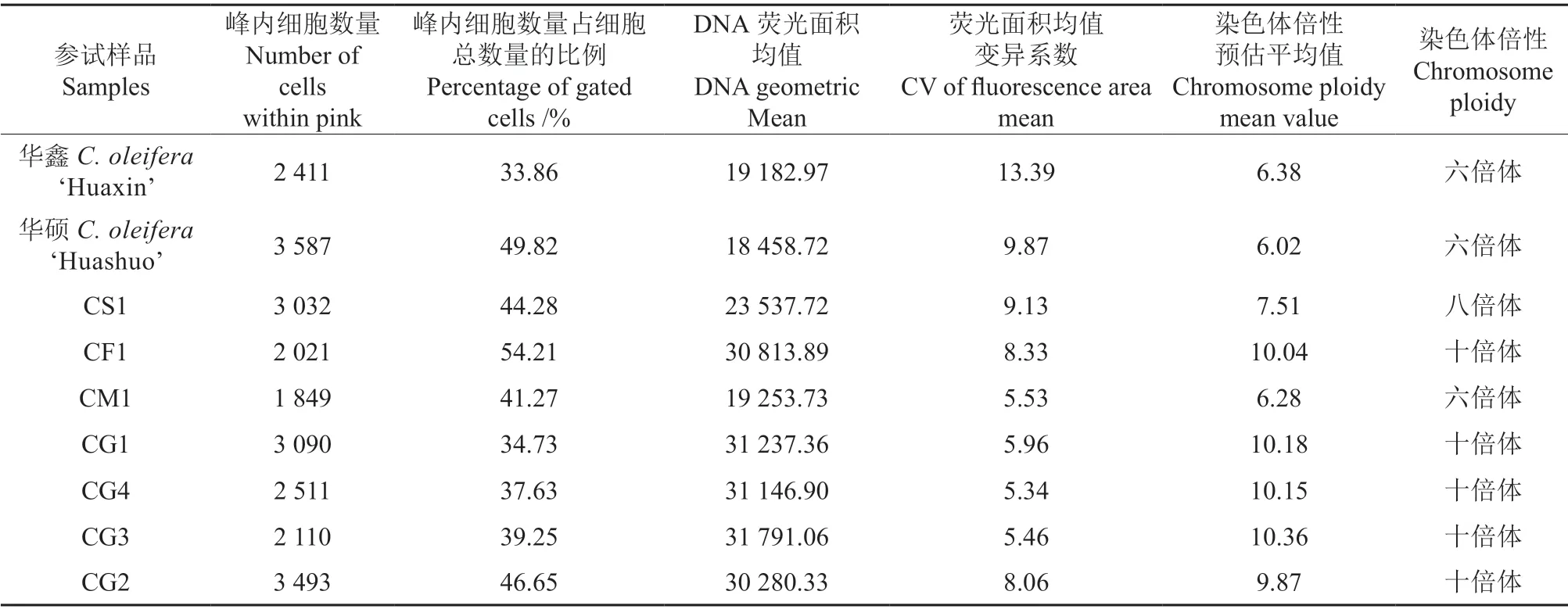
表3 参试油茶样品染色体倍性估测参数Table 3 Parameters of chromosome ploidy estimation for the tested varieties of Camellia-oil species
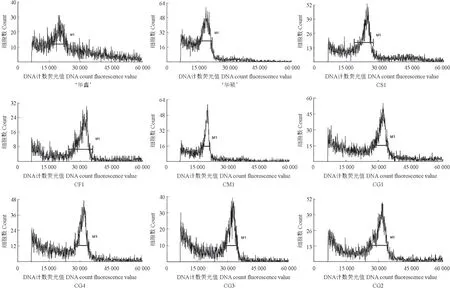
图1 参试油茶样品流式细胞峰值分布Fig.1 Peak distribution of flow cytometry for the tested varieties of Camellia-oil species
2.2 参试样品杂交的保存和坐果情况
参试杂交授粉母树共40 株(表4),含5 个母本无性系,7 个父本无性系,30 个杂交组合。总授粉花朵数量为1 866,授粉后第6 天的授粉花朵保存数量是1 465,授粉后第103 天的坐果数量为713,平均保存率约78.51%,平均杂交坐果率约38.21%。其中,母本CF1 的平均坐果率最低(0.81%±1.7%),但父本CF1 的平均坐果率最高(63.92%±4.76%)。
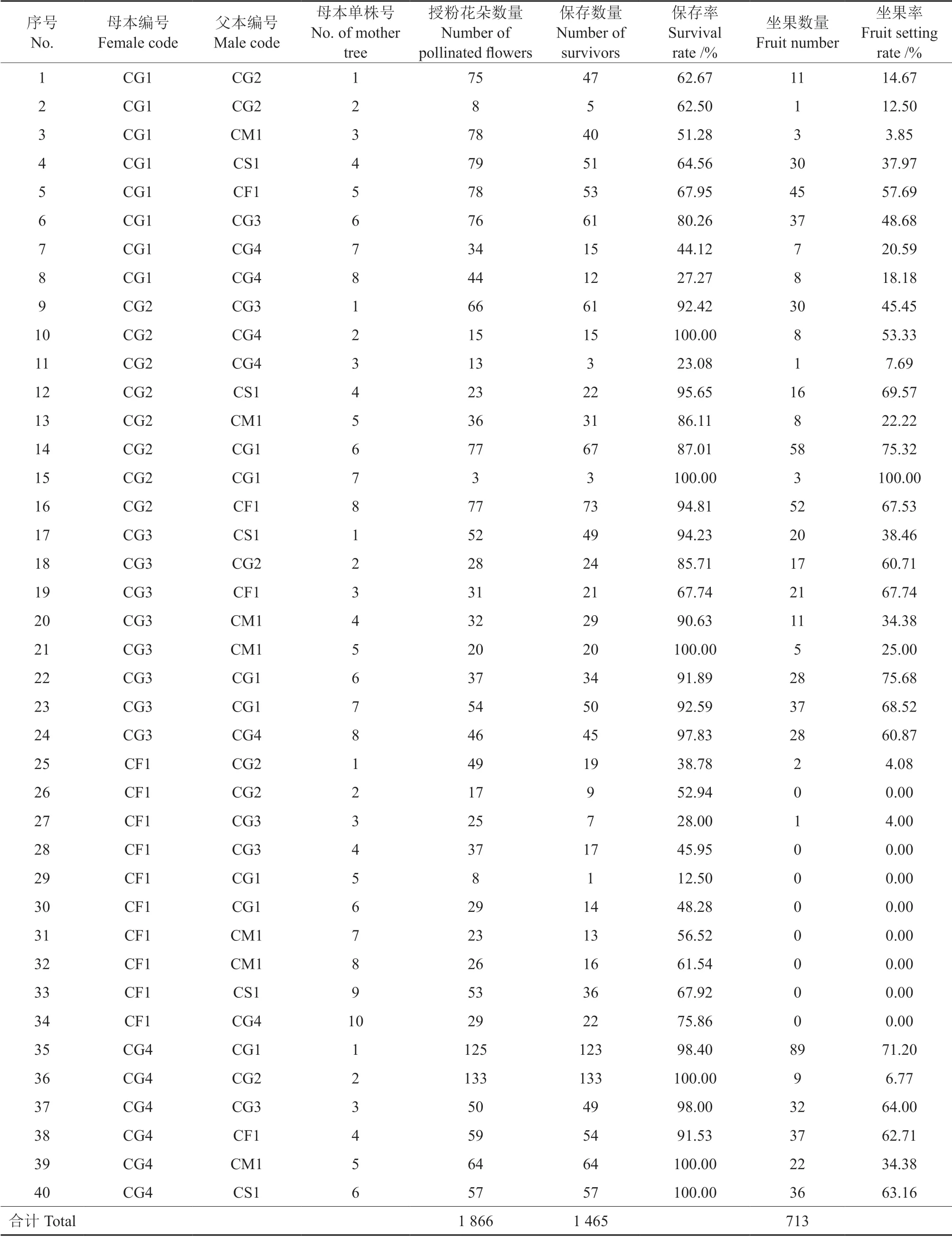
表4 参试油茶杂交试验结果Table 4 The results of hybridization experiment for the tested varieties of Camellia-oil species
2.3 参试样品杂交保存率和坐果率的方差分析
分别对参试杂交授粉母树的授粉花朵保存率和坐果率进行方差分析,结果见表5,采用组内平均值处理母本和父本内的自由度不同导致数据不平衡。由表5可知,保存率在母本间F值为9.020,远大于父本间F值(0.442),存在极显著差异;坐果率在母本间和父本间均存在极显著差异,且母本间F值是父本间的4 倍多。母本间和父本间杂交组合效果存在极大的选择空间,且母本间选择空间较父本间更大。

表5 参试油茶杂交组合的授粉花朵保存率和坐果率的双因素方差分析结果†Table 5 Two factors analysis of variance for survival rate and fruit setting rate for the tested cross combination of Camelliaoil species
组合间方差分析二维交互作用结果如图2所示。由图2可见,保存率交互效应表现最好的是♀CG4 和♂CS1,而坐果率交互效应表现最好的是♀CG2 和♂CG1。
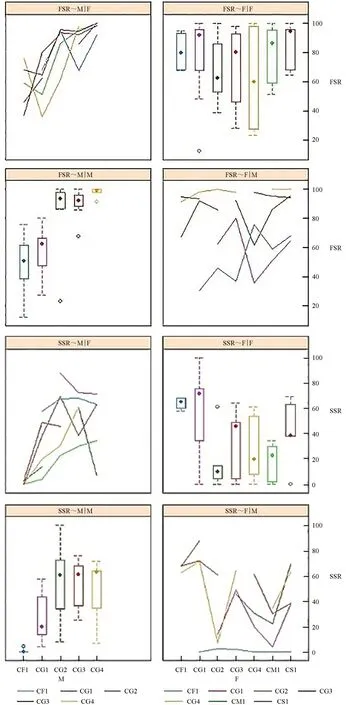
图2 参试油茶杂交组合的授粉花朵保存率和坐果率两两交互作用Fig.2 Pairwise interaction of preservation rate and fruit setting rate for the tested cross combination of Camellia-oil species
2.4 参试样品杂交保存率和坐果率的多重比较分析
对参试母本间和父本间的授粉花朵保存率和坐果率进行0.05 水平多重比较分析,结果见表6。由表6可知,母本间保存率差异度为ab,均值为75.87%,保存率最高为CG4(97.99%),保存率最低为CF1(48.83%)。而高州油茶CG4、CG3、CG2 作为母本的保存率显著高于CG1和CF1。母本间坐果率差异度为abc,均值为37.40%,其中高州油茶CG2、CG3 和CG4 的坐果率显著高于CG1,坐果率最高的为CG2(55.14%),坐果率最低的为CF1(0.81%)。
父本间保存率的差异不显著(表6),均值为73.74%。父本保存率最高的为CS1(84.47%),最低的为CG4(61.36%);父本间坐果率差异度为abcd,其中CF1、CG1、CS1 的坐果率显著高于其他父本,均值为36.33%,坐果率最高的为CF1(63.92%),最低的是CG2(16.46%)。
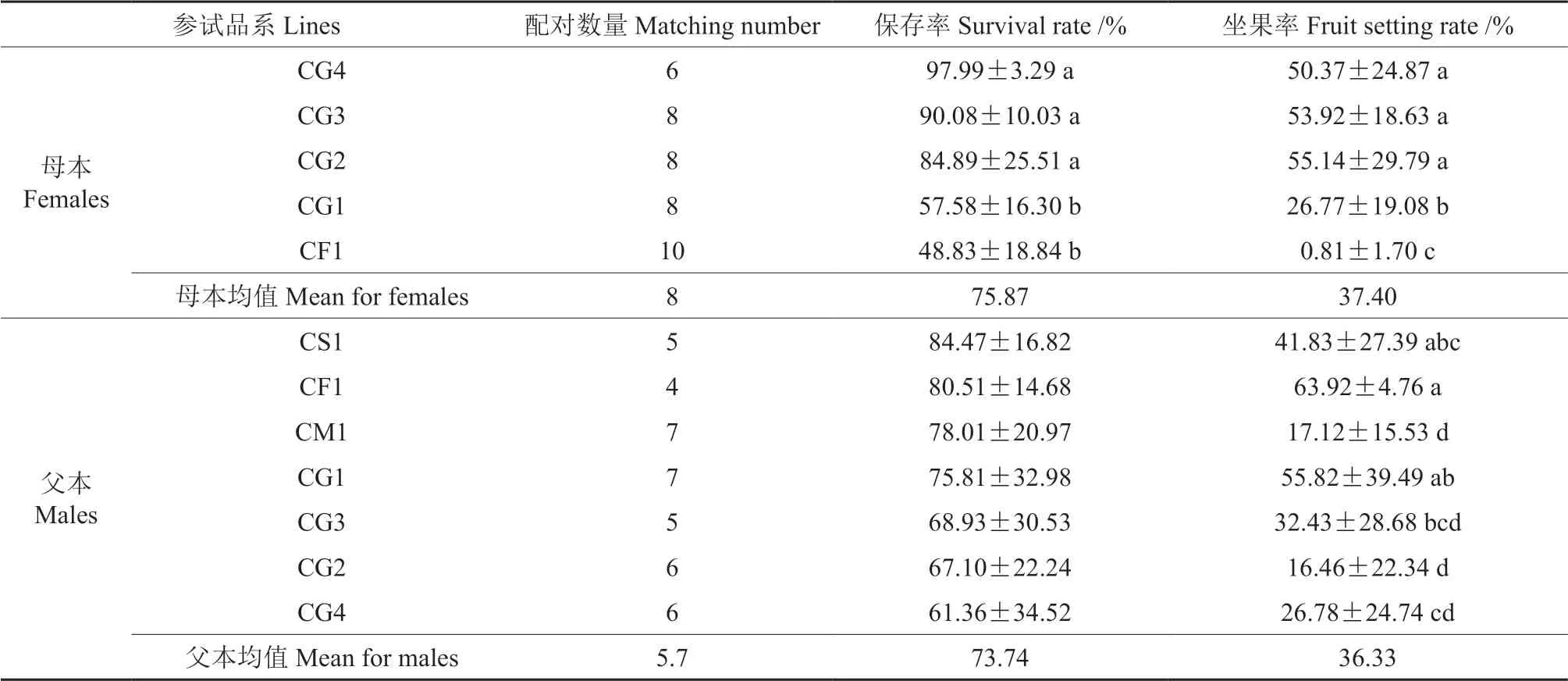
表6 参试油茶杂交组合的授粉花朵保存率和坐果率的多重比较结果†Table 6 Multiple comparisons of survival rate and fruit setting rate for the tested cross combination of Camellia-oil species
2.5 参试样品杂交坐果率影响因素分析
杂交试验中参试样品的平均坐果率偏低,表明作为母本的香花糙果茶(糙果茶组)与作为父本的高州油茶等(油茶组)种间杂交亲和性较差。平均坐果率偏低的另一个影响因素可能是气候,低温天气对杂交造成了影响。
此次人工杂交授粉时间段为2019年12月19—21日和2019年12月25—27日,共计6 d,中间出现1 次寒潮,夜间温度为2 ~3 ℃,白天为8 ~10 ℃,导致部分母树次日开花数量骤减。从母本花朵日开发数量(表7)可以看出,不同母本的花朵日开放数量变化较大。配置授粉组合时,需要根据花蕾数量和花朵开放程度进行规划,从而保证花粉活性和花朵开放条件基本一致,保障授粉顺利完成。此次授粉是在秋冬季节第1 次降温之后进行的,3 d 的低温对花朵的开放无明显影响,但需要进一步观测持续低温对高州油茶授粉后受精及果实发育的影响。
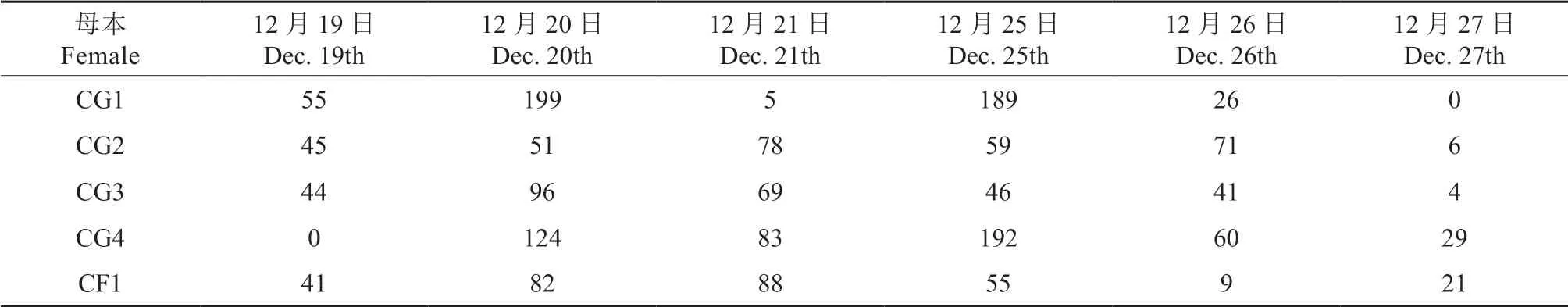
表7 参试油茶杂交过程中母本花开放数量的变化Table 7 Flower numbers change for female parents during cross-hybridizations
2.6 参试样品杂交配合力
采用亲本模型I 估算授粉花朵保存数量和坐果数量的配合力效应值。亲本模型I 方差分析结果(表8)显示,母本间、父本间和组合间的授粉花朵保存数量和坐果数量效应值均达到极显著水平,说明父本间、母本间在授粉花朵保存数量和坐果数量的一般配合力和组合间的特殊配合力均存在极显著的选择空间。
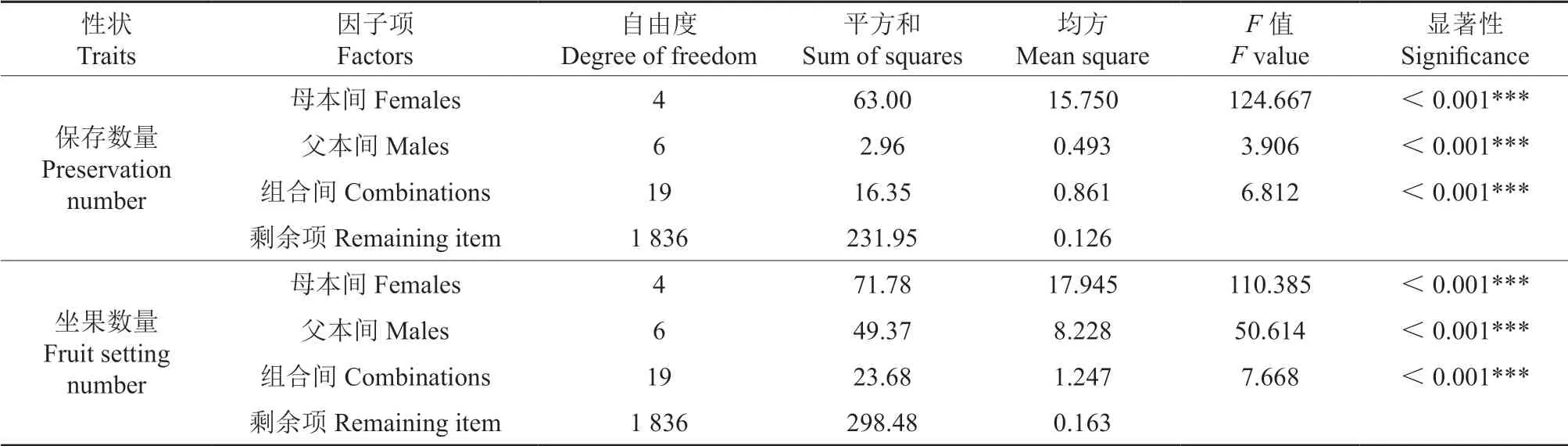
表8 参试油茶杂交组合授粉花朵保存数量和坐果数量的亲本模型Ⅰ方差分析结果†Table 8 Mixed linear model fitting for survival numbers and fruit setting numbers for the tested cross combination of Camellia-oil species
从亲本模型Ⅰ各因子方差分量估计参数(表9)可知,授粉花朵保存数量和坐果数量在母本间的方差分量大于父本间和组合间,授粉花朵保存数量在父本间方差分量为负值,在母本间、组合间及剩余项中均为正值。去掉组合间的效应时,混合线性模型父本间方差分量呈现为正值。为了观测特殊配合力,采用不去除组合间效应的模型进行配合力估算。
根据亲本模型Ⅰ估算授粉花朵保存数量的一般配合力和特殊配合力,结果见表10 ~11。由表10 可知,授粉花朵保存数量在母本间一般配合力最高的是CG4(0.191),其次是CG3(0.107),最低的是CF1(-0.221);授粉花朵保存数量在父本间一般配合力最高的是CS1(<0.001),最低的是CG4(>-0.001)。由表11 可知,授粉花朵保存数量的特殊配合力以♀CG1×♂CG3 为最高(0.167),其次是♀CF1×♂CG4(0.148)和♀CF1×♂CS1(0.102)。
根据亲本模型I 估算坐果数量的一般配合力和特殊配合力,结果见表10 ~11。由表10 可知,坐果数量在母本间一般配合力最高的是CG3(0.157),最低的是CF1(-0.328);坐果数量在父本间一般配合力最高的是CG1(0.137),最低是CM1(-0.165)。由表11 可知,坐果数量的特殊配合力以♀CG2×♂CG1 为最高(0.177),其次是♀CG3×♂CG2(0.139)和♀CG2×♂CS1(0.113)。
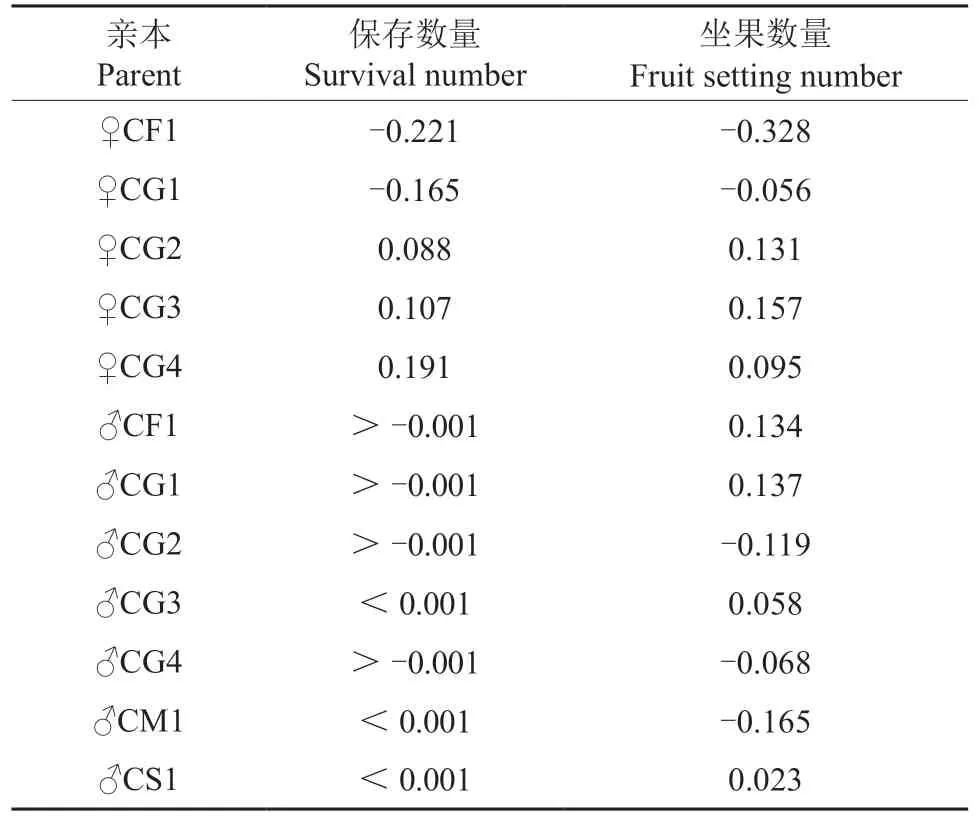
表10 参试油茶杂交组合授粉花朵保存数量和坐果数量的一般配合力Table 10 GCA and SCA for survival numbers and fruit setting numbers
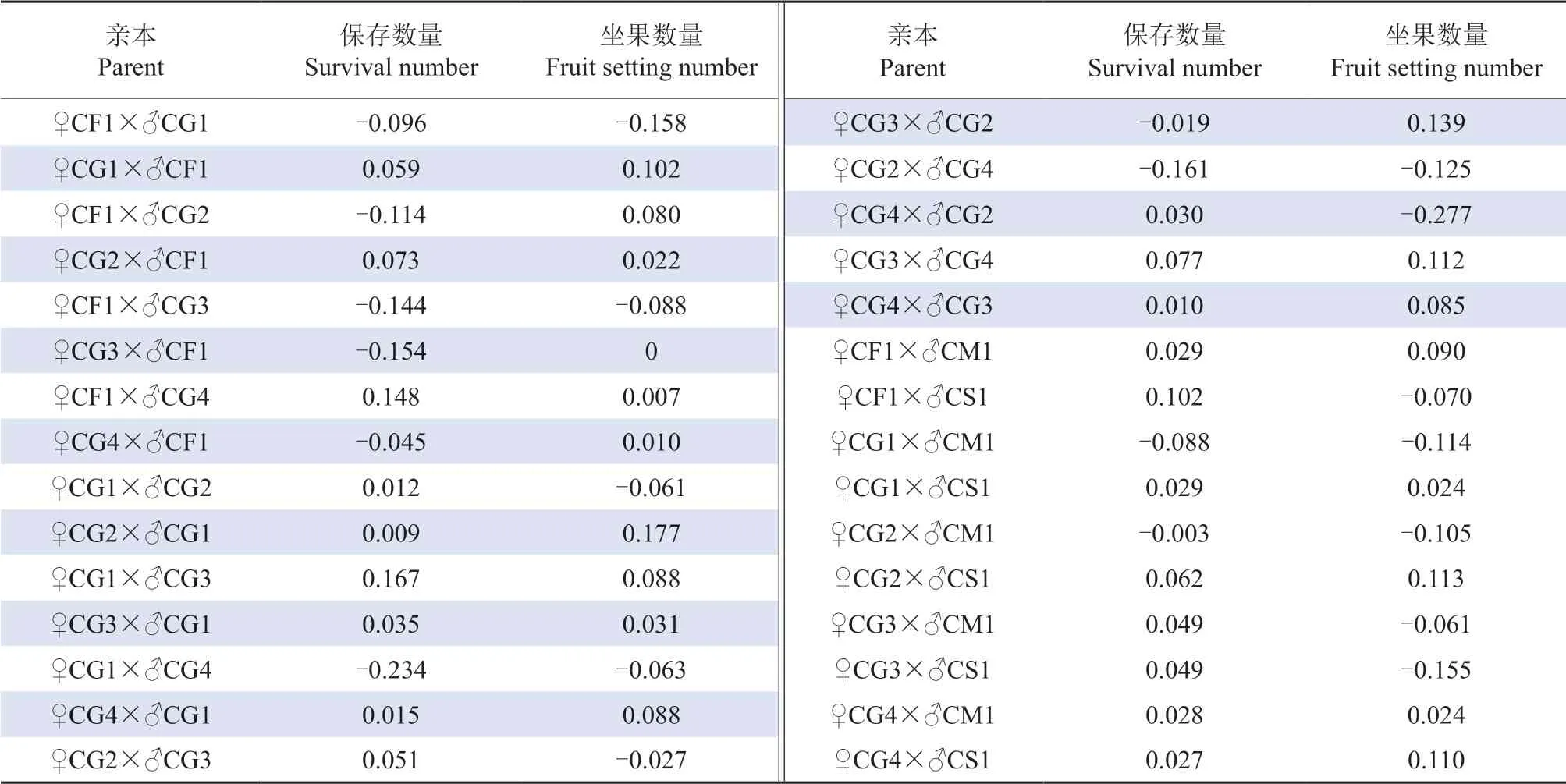
表11 参试油茶杂交组合授粉花朵保存数量和坐果数量的特殊配合力†Table 11 GCA and SCA for survival numbers and fruit setting numbers
3 结论与讨论
高州油茶种内杂交亲和性普遍高于种间杂交。香花糙果茶不适宜作为高州油茶、小果油茶和普通油茶的杂交母本,更适合作为父本或配置授粉树;高州油茶、小果油茶和普通油茶杂交可互为亲本,坐果率差异化和染色体倍性以及亲缘关系密切相关;高州油茶CG3 作母本的杂交亲和性各参数表现最好,CG1 作父本杂交亲和性最好。明晰种间杂交亲本可配性,通过高亲和性的种内杂交亲本正反交,获得大量改良的杂交子代将是高州油茶遗传改良的重要方法。
3.1 高州油茶种间杂交不亲和因素
本试验中,杂交亲本来自4 个油茶树种,分别是高州油茶、普通油茶、小果油茶和香花糙果茶,但在文献[4]中其分类归属有部分变化。本文中按照油茶育种传统的物种名称(异名)进行表述,不同树种按种间杂交处理。
植物正交和反交对亲和性或可育性的响应称为植物的向性,即杂交亲本的搭配方式或方向[29]。全球山茶亚属约6 组112 种,我国约6 组105 种[4]。油茶树种在组内和组间的种间杂交存在一定的向性规律,主要分为双向可育、单向不育和双向不育等。油茶中典型的双向可育组间杂交是普通油茶与小果油茶杂交,其次是小果油茶与高州油茶杂交,以及普通油茶与高州油茶杂交[30]。本试验中,以高州油茶作母本(CG1、CG2、CG3 和CG4),以小果油茶(CM1)作父本进行杂交的坐果率为3.85%~34.38%,平均为23.97%;以高州油茶作母本(CG1、CG2、CG3 和CG4),以岑软3 号(CS1)作父本的组合中,坐果率为37.97%~69.57%,平均为52.29%。高州油茶、小果油茶和普通油茶的双向可育现象与其他试验结果一致[31],该现象在杜鹃花属中也有发现[32]。
种间杂交在组间和属间存在较多的单向不育现象。表现为雌性不育的原因有其他树种花粉与柱头的绒毡层不亲和,或者是胚珠、胚囊、卵细胞等雌性器官发育异常等[33];雄性不育的主要原因有花粉质量降低,或花粉管表达异常等[34]。本试验中香花糙果茶(CF1)呈雌性不育表现,将其作母本与小果油茶和高州油茶杂交时,各组合坐果率为0 ~4.08%,平均坐果率为0.81%;香花糙果茶为十倍体,小果油茶为六倍体,高州油茶为十倍体,既有同倍性又有异倍性,但均表现为不育,染色体倍性差异导致其雌性不育的因素偏小[35-36]。但以香花糙果茶作父本时,各组合的坐果率为57.69%~67.74%,平均坐果率为63.92%。因此,香花糙果茶单向雌性不育或与柱头绒毡层表达异常等相关性较大。
此外,试验中观察到以高州油茶CG2 作父本时,各组合的坐果率为0 ~60.71%,平均坐果率为16.46%,其母本按照坐果率从大到小排列依次是CG3、CG1、CG4、CF1。除 了♀CG3×♂CG2组合的坐果率为60.71%,其他种内杂交组合(♀CG1×♂CG2 与♀CG4×♂CG2)的坐果率均低于15%,种间杂交♀CF1×♂CG2 组合授粉后受精全部失败,这种强烈的杂交成活率差异化可能是亲缘关系所导致[37]。
以小果油茶作父本的坐果率为0 ~34.38%,平均坐果率为17.12%,其母本按照坐果率从大到小排列依次是CG4、CG3、CG2、CG1、CF1。试验中观测到广东兴宁的小果油茶为六倍体[35],与其他十倍体高州油茶杂交时,作父本时坐果率更高,作母本时坐果率则普遍更低,如小果油茶×博白大果油茶、小果油茶×广宁红花油茶[37],结果表现为不孕。从亲和性角度来看,高倍性更适宜作母本,低倍性更适宜作父本。
3.2 高州油茶种内杂交
以高州油茶CG4、CG3 和CG2 作母本的平均坐果率普遍高于50.37%,以CG1、CG3 作父本的平均坐果率大于32.43%,高州油茶种内杂交亲和性较好。如:♀CG1×♂CG3 组合的坐果率为48.68%,授粉76 袋,可以收37 个果实;♀CG2×♂CG1 组合的坐果率为75.32%,授粉77 袋,可收58 个果实。♀CG3×♂CG2(60.71%)、♀CG3×♂CG1(68.52%~75.68%)、♀CG3×♂CG4(60.78%)、♀CG4×♂CG3(64.00%)组合的坐果率均较高,有利于获得数量众多的子代苗,也是油茶高产的重要性状之一。特别是♀CG4×♂CG1 组合,授粉125 袋,收获89 个果实,坐果率达71.20%。这些组合的坐果率均远高于普通油茶×越南油茶、南荣油茶×越南油茶的正交坐果率和小果油茶×越南油茶的正反交坐果率(33.71%~56.40%)[37],有望成为高州油茶中的优势组合。高州油茶种内杂交坐果率远高于种间杂交的结果表明,同倍性的油茶树种之间的杂交亲和性高于种间的组合[30,38]。
作为中间指标,母本中以CG4 的授粉花朵保存率最高(97.99%),其次是CG3(90.08%),在父本中以CS1 的授粉花朵保存率最高(84.47%),其次是CM1(80.51%)。在2 次观测中,授粉花朵保存率不断下降,以CF1 为母本的平均授粉花朵保存率降幅为48.02%,以CG4 为母本的降幅为47.62%,以CG3 为母本的降幅为36.16%。保存率降低的原因可能有胚珠和子房发育异常、营养元素缺失等。从品种培育的角度考虑,减少“千花一果”将是促进油茶增产的重要研究方向。
尽管种间杂交坐果率偏低,但染色体高倍性的亲本在吸收了低倍性物种的基因后,再通过同倍性杂交的高亲和性,最终导致其种内变异非常丰富。在2021年观测的高州油茶育种亚群体自由授粉子代林13年生林分中,单果鲜质量的变异为12.31 ~260.60 g,单果鲜质量的最高值是其最低值的21.17 倍。单果鲜质量最高达到了广宁红花油茶的单果鲜质量,最小单果鲜质量小于普通油茶果实;果形丰富多样,有圆形、橘形、扁圆形、橄榄形,有棱、底部凹陷等;果皮颜色有红色、黄色、绿色等。因此,种内杂交结合种间杂交可作为综合改良油茶的重要方法。
3.3 高州油茶杂交亲和性的配合力
一般配合力通常为性状的加性效应,特殊配合力一般为杂交组合的显性和上位效应等。试验中母本间的一般配合力方差分量普遍大于父本间和组合间的一般配合力方差分量,说明母本间加性效应普遍高于父本间加性效应和组合间显性和上位效应。
各母本按照坐果数量的一般配合力由大到小排列依次是CG3、CG2、CG4、CG1、CF1,按照坐果率平均值由大到小排列依次是CG2、CG3、CG4、CG1、CF1;各父本按照坐果数量的一般配合力由大到小排列依次是CG1、CF1、CG3、CS1、CG4、CG2、CM1,按照坐果率平均值由大到小排列依次是CF1、CG1、CS1、CG3、CG4、CM1、CG2。尽管4 个高州油茶品系中以CG2 作母本的坐果率最高(55.14%),但以CG3 作母本的坐果数量的一般配合力最高(0.157)。以CG1作父本的一般配合力最高(0.137),但坐果率低于雌性不育的香花糙果茶。在进行亲本选择时,与坐果率平均值相比,以一般配合力为参考指标更为合理。综合考虑上述亲和性参数,可选择CG3 作为最优母本,选择CG1 作为最优父本。
特殊配合力以♀CG2×♂CG1 组合最高,其次是♀CG3×♂CG2 组合。CG3×CG1 和CG4×CG3正反交的特殊配合力均是正数,CG2×CG4 正反交的特殊配合力均是负数,这2 个组合出现超亲现象的可能性较大。其他正反交组合间的特殊配合力多为一正一负,或者CF1 作为母本存在不孕现象。因此,CG3 坐果数量的特殊配合力存在明显优势,且母本一般配合力效应值最高,说明CG3 是试验中4 个高州油茶亲和性表现最稳定的亲本,可作为高亲和性优势亲本,也有可能表现出超亲现象。
3.4 研究局限及研究方向
本文中采用混合线性模型拟合保存数量和坐果数量这一类阈性状的遗传参数[26]。该类模型基于最优无偏线性预测(best linear unbiased prediction,BLUP)方法,构建A 矩阵描述不同个体之间的成对关系,估算各组合的遗传参数。该类模型较多应用于二倍体物种,在四倍体参数估算中也有较好的案例[39-41],且四倍体和二倍体之间遗传参数相关性为0.95 ~1.00。本文中基于二倍体混合线性模型初步对各高倍性亲本进行遗传参数估算,但四倍体以上物种的遗传参数估算方法和配套软件等有待进一步优化。油茶因亲、子代培育周期过长,不利于作为上述模型优化的模式树种。此外,研究油茶杂交子代的表型观测分析和气候因素对杂交的影响,是完善油茶杂交育种技术的方向。
致谢:广东省林业科学研究院的唐旭晓、王海玲、林韦希、陈韵、欧阳剑杰及仲恺农业工程学院温敏、王晓阳、黄润生、栗永娟、谈静、王龙远、薛彬娥参与了亲本自然结实、花朵数量统计和杂交授粉调查。
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
世界热带农业信息(2022年11期)2022-10-18
吉林林业科技(2020年6期)2020-12-16
岷峨诗稿(2019年4期)2019-04-20
猪业科学(2018年5期)2018-07-17
岭南音乐(2016年5期)2017-01-17
岭南音乐(2016年5期)2017-01-17
河北农业科学(2016年2期)2016-09-09
华南农业大学学报(2015年5期)2015-12-04
食品与机械(2012年2期)2012-12-28
