
圆齿野鸦椿嫁接愈合过程的显微观察及嫁接体酶活性的变化分析
2022-12-05 06:53万瑞琪姜雪茹涂淑萍
经济林研究 2022年3期
黄 航,刘 聪,万瑞琪,姜雪茹,涂淑萍
(江西农业大学 林学院,江西 南昌 330045)
圆齿野鸦椿Euscaphis konishiiHayata 为省沽油科Staphyleaceae野鸦椿属Euscaphis常绿小乔木,是我国特有的药用与观赏树种[1]。该树种的果皮于9月开始逐渐张开,露出鲜明艳丽的红色内果皮,内果皮上附着黑色的种子,观赏价值极高,观果期跨越秋、冬和春(翌年)3 个季节,历时7 个月之久。圆齿野鸦椿可丛植或列植,用于庭院、街道、风景名胜区的绿化与美化,以丰富秋、冬季园林景观及生物多样性,还可盆栽观赏[2]。据报道,野鸦椿属植物富含酚酸类、木脂素类和黄酮类等生理活性物质,具有抗肿瘤、抗菌、消炎等功效[3],其推广应用前景十分广阔。目前,主要采用播种繁殖技术繁育圆齿野鸦椿的苗木。但是,其种子发芽率低,且有深休眠特性[4],需经层积沙藏、热水浸泡处理方可发芽[5];而且播种繁殖后代的性状参差不齐,不能保持良种的优良性状。扦插繁殖需选择在3年以下的幼树上剪穗扦插,而在成年树上剪穗扦插,插穗成活率极低[6],且因受插穗的限制,繁殖系数也较低。组织培养污染率高,成本昂贵,技术要求严格[7]。而嫁接育苗可保持良种的优良性状,能使其提前开花结果,增强其抗性,提高其品质,加快良种繁育速度[8-9]。因此,关于圆齿野鸦椿嫁接及愈合机理的研究,是提高良种繁育速度,实现良种化栽培的重要研究课题。前人关于嫁接愈合过程解剖学观察[10]及酶活性的变化[11]等方面的研究报道已较多:黄蔓娜等[12]在对梨树嫁接体愈合过程的观察研究中将其分为隔离层产生期、愈伤组织产生期、愈伤组织分化与对接期、新输导组织形成期4 个阶段;骆建霞等[13]和Errea 等[14]的研究结果表明,砧、穗在嫁接愈合过程中的解剖结构在一定程度上能反映出嫁接愈合的亲和性;此外,接口的愈合与相关酶活性密切相关,如过氧化物酶(POD)和过氧化氢酶(CAT)都是植物体内的保护酶,在一定程度上均影响着嫁接体接口的愈合[15]。而多酚氧化酶(PPO)是末端氧化酶之一,当植物受到伤害时,其易催化多酚物质形成醌,从而对植物起到保护作用;苯丙氨酸解氨酶(PAL)参与细胞分化与木质素的合成。可见,POD、CAT、PPO 和PAL 均为嫁接体愈合过程的重要评价指标[16]。目前,关于圆齿野鸦椿嫁接愈合过程的生理生化、解剖学等嫁接机理方面的研究尚未见诸报道。为给圆齿野鸦椿良种嫁接快繁育苗技术的研究提供理论参考依据,本研究以适应性强、分布范围广的同属落叶树种野鸦椿E.japonica作砧木,以观赏价值高的常绿树种圆齿野鸦椿作接穗,于2020年7月进行嵌芽接,对其接口愈合过程进行显微观察,并对嫁接体相关酶活性的变化情况进行研究,以探讨圆齿野鸦椿嫁接成活的机理。
1 材料与方法
1.1 试验材料及处理
以江西农业大学林学院花卉盆景基地(28°46′N,115°55′E)内2年生的盆栽野鸦椿实生苗为砧木,以8年生圆齿野鸦椿的当年生枝上健壮腋芽为接穗,2020年7月下旬在砧木离地约20 cm 处进行嵌芽接,于嫁接后不同时间剪取嫁接口1.5 cm 左右的茎段为试验材料,对其进行解剖学观察和相关酶活性的测定。嵌芽接的具体操作步骤为:先在接穗的芽上方处稍带木质部向下斜切一刀,长约2.5 cm,再在芽下方角度为30°处斜切至第一切口底部,取下带木质部芽片,在砧木光滑处自上而下稍带木质部削一与接芽片大小相等的切面,随后将芽片插入砧木切口并将两者形成层对齐,用嫁接膜自下而上绑紧,仅将芽片上的叶柄露出即可。
1.2 试验方法
1.2.1 圆齿野鸦椿嫁接愈合过程的解剖学观察
于嫁接当天至嫁接后第8 天每天取样1 次,嫁接后第8 ~22 天每隔2 d 取样1 次,嫁接第22 天后每隔4 d 取样1 次,至嫁接体成活为止。每次采集2 个嫁接体接口茎段,用70% 的FAA 固定液固定,置于4 ℃冰箱中保存以备用。将固定好的样品取出,采用10%的乙二胺软化处理45 d后,以流水冲洗2 h,以乙醇逐级脱水,每级脱水30 min,然后将脱水后的材料依次置于25%的二甲苯、50%的二甲苯、75%的二甲苯、纯二甲苯、纯二甲苯中进行透明处理,每次透明时间20 min,包埋,随后将修整好的样品置于Leica RA2235 旋转切片机上切片,切片厚度为10 µm,展片,粘片,脱蜡,番红固绿染色,封片,制成永久石蜡切片。最后,将切片置于德国产的ZEISS Scope.A1 显微镜下观察并拍照。
1.2.2 圆齿野鸦椿嫁接愈合过程嫁接体相关酶活性的测定
于嫁接当天开始取样,每隔4 d 取样1 次,每次随机选取3 ~4 个茎段,重复3 次。将样品取下后立即置于液氮罐中速冻,带回实验室置于-80℃的超低温冰箱中保存以备用。
采用钼酸铵比色法测定过氧化氢酶(CAT)活性,采用分光光度法测定过氧化物酶(POD)、多酚氧化酶(PPO)和苯丙氨酸解氨酶(PAL)的活性;所用试剂盒均由苏州科铭生物技术有限公司生产。
1.3 数据处理
使用Excel 2010 软件对数据进行处理并绘图,利用Photoshop CS5 软件对照片进行处理,采用SPSS 17.0 软件对数据进行统计分析。
2 结果与分析
2.1 圆齿野鸦椿嫁接愈合过程的解剖学观察结果
以野鸦椿为砧木嫁接圆齿野鸦椿后,对其嫁接体的愈合过程进行显微观察,结果表明,其愈合过程可划分为3 个时期,即砧穗切口隔离层及愈伤组织形成期、砧穗结合部愈伤组织增殖与连接期、砧穗结合部愈伤组织分化形成维管组织期。
2.1.1 砧穗切口隔离层及愈伤组织形成期
砧穗切口隔离层及愈伤组织形成的过程如图1所示。从图1A 中可以看出,嫁接后第3 天在砧木的切面上可见一层染色较深的薄层,即为隔离层,而接穗的切面直到嫁接后第6 天才可观察到隔离层(图1C),说明砧木比接穗提早3 d 形成隔离层,早期形成的隔离层对伤口具有一定的保护作用。从图1B 中可以看出,嫁接后第5 天,砧木切口周围的部分形成层、木质部、木射线等部位的细胞开始脱分化形成愈伤组织。此时,横切面上砧木木质部薄壁细胞、木射线脱分化的细胞径向开始增厚并向外凸起,薄壁细胞明显增加,无规则地堆积在一起,部分薄壁细胞体积较大,清晰可见内含较大的细胞核,隔离层清晰可见。从图1D 中可以看出,嫁接后第7 天,砧木皮层、韧皮部薄壁细胞突破了隔离层,经脱分化形成愈伤组织,并于其四周大量分裂增殖,形成愈伤组织细胞团,多数细胞呈无序排列状。而接穗切面于嫁接后第10 天在皮层和韧皮部处开始形成愈伤组织(图1E)。可见,接穗较砧木晚5 d 形成愈伤组织。
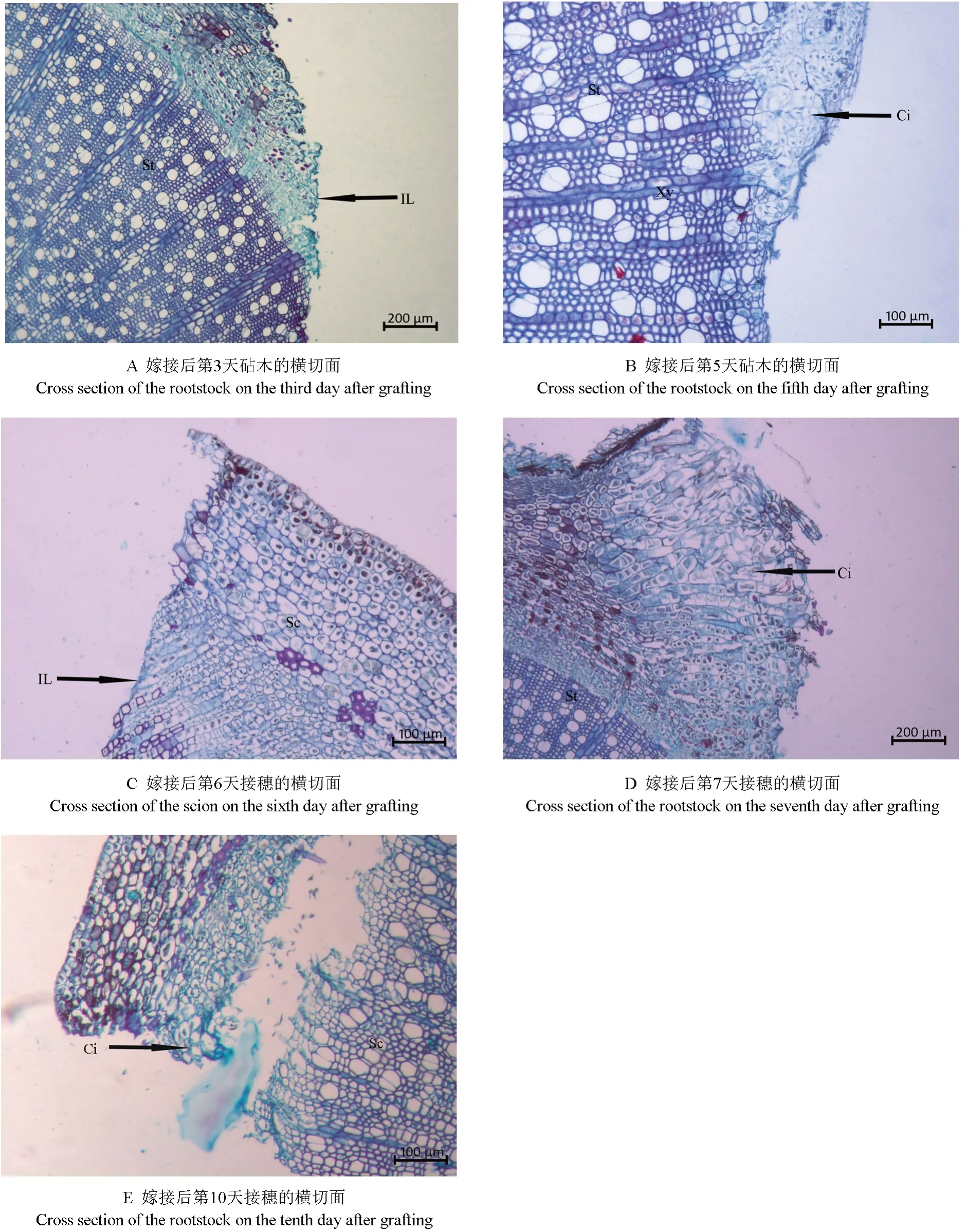
图1 隔离层及愈伤组织形成过程的显微观察结果Fig.1 Observation on the microstructure of isolation layer and callus period
2.1.2 砧穗结合部愈伤组织增殖与连接期
砧穗结合部愈伤组织增殖与连接的过程如图2所示。从图2A 中可以看出,嫁接后第14 天,砧木和接穗间的空隙因愈伤组织的抱合和连接而被填满,二者紧密结合形成愈伤组织桥,而隔离层已消失。此时,愈伤组织桥为砧穗间细胞流通的主要途径。嫁接后第20 天,砧穗结合部构成形成层环,表明嫁接体已经愈合(图2B)。

图2 愈伤组织增殖与连接过程的显微观察结果Fig.2 Observation on the microstructure of callus proliferation and connection period
2.1.3 砧穗结合部愈伤组织分化形成维管组织期
砧穗结合部愈伤组织分化形成维管组织的过程如图3所示。由图3A 可以看出,嫁接后第26 天,嫁接体形成层的细胞分化出较多结构规整的木质部和韧皮部,较大口径的导管已清晰可见,此时嫁接体已经形成了一个新的个体。嫁接后第40 天,新分化出的维管束扩大分化,维管束呈现出不规整的分布状,形成维管束混合群体(图3B)。
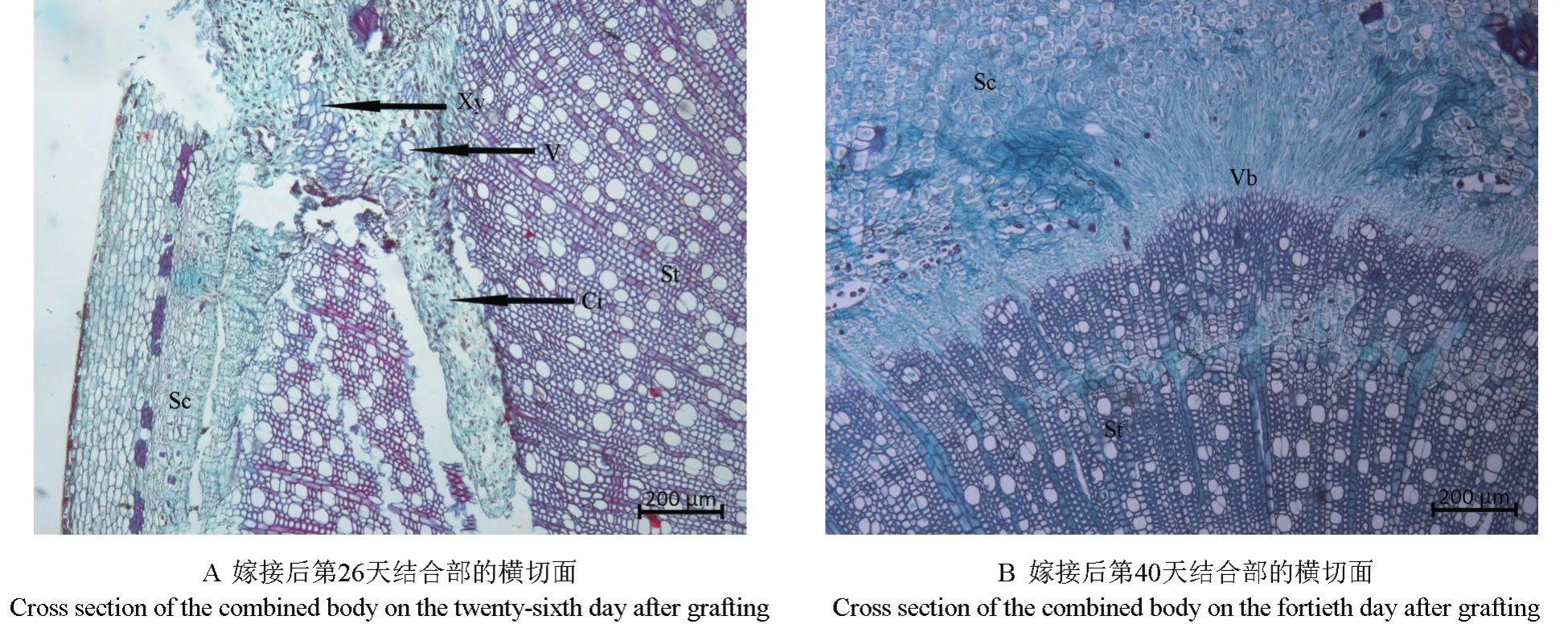
图3 愈伤组织分化形成维管组织过程的显微观察结果Fig.3 Observation on the microstructure of callus during differentiation into vascular tissue
2.2 圆齿野鸦椿嫁接愈合过程中嫁接体酶活性的变化趋势
2.2.1 过氧化氢酶和过氧化物酶活性的变化趋势
嫁接口愈合过程中嫁接体内过氧化氢酶(CAT)和过氧化物酶(POD)活性的变化趋势如图4所示。嫁接当天至嫁接后第20 天,嫁接体内CAT 和POD 活性的变化趋势基本一致。在嫁接后第10 天之前即砧穗切口隔离层和愈伤组织形成期,CAT 和POD 活性均显著上升,其中,CAT 活性在嫁接第10 天达到峰值。在嫁接后第10 ~15 天即嫁接口愈伤组织细胞分裂增殖期,CAT 和POD 活性均显著下降。在嫁接后第15 ~20 天即砧穗愈伤组织连接愈合期,CAT 和POD 活性均显著上升。在嫁接后第20 ~25 天即砧穗结合部愈伤组织进行新生维管组织分化期,CAT 和POD 活性开始出现不一致的变化情况,即CAT 活性下降,而POD活性上升。

图4 嫁接体内过氧化氢酶和过氧化物酶活性的变化趋势Fig.4 The change trend of CAT and POD activities in graft
2.2.2 多酚氧化酶和苯丙氨酸解氨酶活性的变化趋势
嫁接口愈合过程中嫁接体内多酚氧化酶(PPO)和苯丙氨酸解氨酶(PAL)活性的变化趋势如图5所示。在嫁接口的愈合过程中,PPO 和PAL 活性的变化趋势正好相反。在嫁接后第10 天之前即砧穗切口隔离层和愈伤组织形成期,PPO 活性显著升高,而PAL 活性显著降低;在嫁接后第10 ~15 天即砧穗结合部愈伤组织细胞分裂增殖期,PPO 活性显著降低,而PAL 活性升高;在嫁接后第15 ~20 天即砧穗结合部愈伤组织连接愈合期,PPO 活性较前稍有降低,而PAL 活性显著升高;在嫁接后第20 ~25 天即砧穗结合部愈伤组织分化形成维管组织期,PPO 活性升高而PAL活性降低。

图5 嫁接体内多酚氧化酶和苯丙氨酸解氨酶活性的变化趋势Fig.5 The change trend of PPO and PAL activities in graft
3 讨 论
3.1 嫁接体接口愈合过程的解剖学观察结果分析
嫁接成活过程实质上是砧、穗结合部位通过伤口产生愈伤组织、形成愈伤组织桥和形成层环,分化维管束组织,形成一个新个体的过程[17]。初庆刚等[10]将梨树嫁接愈合过程划分为愈伤组织的形成期和维管形成层的形成及活动期这2 个时期;兖攀等[18]将美国山核桃嫁接愈合过程划分为砧穗隔离期、愈伤组织生长期、形成层环构形成期和新生维管组织分化期这4 个时期。不同物种嫁接愈合的组织学特征存在差异,而其基本结构及愈合过程却大致相同。拟南芥嫁接后第3 天即分化出大量维管束结构[19];早实核桃芽接后第6 天砧穗维管束就已重新连接[20];黄瓜嫁接后第15 天维管束完全建成[21];梨树嫁接后第20 天新的输导组织就已形成[10];油茶芽苗砧嫁接后第35 天愈伤组织维管束才形成[22]。综上所述,物种不同,嫁接体木质化程度不同,嫁接成活的快慢则不一致。显微观察结果表明,以‘野鸦椿’为砧木嫁接‘圆齿野鸦椿’后,嫁接体在嫁接后第26 天已形成了新的导管和筛管,使砧穗间能够正常进行养分和水分的输送,并形成了一个新的个体。
嫁接时,砧、穗切口细胞受到机械损伤,细胞的内含物与残壁发生氧化反应,形成高分子物质,隔离层覆盖整个嫁接面[22]。植物遭受创伤后易形成隔离层,嫁接初期隔离层能有效防止水分的蒸发,保护伤口不被病原体入侵,但是,隔离层太厚也会影响砧穗间愈伤组织的形成[11]。显微观察结果还表明,切口处开始出现隔离层与愈伤组织的时间,砧木比接穗分别早3 和5 d,砧木早于接穗恢复生理活动,故有利于嫁接体的愈合成活。这一观察结果与杨志坚等[22]的研究结果一致。其中,砧木切口处隔离层与愈伤组织开始形成的时间分别为嫁接后第3 天和第5 天,与接穗切口处隔离层与愈伤组织开始形成的时间相比,分别早3 和5 d。随着嫁接体的发育,嫁接第5 ~10 天,砧穗切口处部分细胞突破隔离层,形成愈伤组织,愈伤组织细胞在分化过程中不断生长。
嫁接后第10 ~20 天,为砧、穗切口处愈伤组织细胞分裂增殖期;嫁接后第14 ~20 天,为砧、穗愈伤组织连接愈合期;嫁接后第14 天,砧、穗间的空隙被愈伤组织填满,嫁接体形成愈伤组织桥,隔离层消失;嫁接后第20 天,砧穗结合部构成形成层环,嫁接体完成愈合过程。兖攀等[18]观察发现,愈伤组织分化形成愈伤组织桥和形成层环,并随即产生新的输导组织(如韧皮部、木质部),砧、穗便接合成一个整体。兖攀等[18]的观察结果与本研究结果一致。
嫁接愈合过程中,嫁接体的显微结构在一定程度上可以反映砧、穗嫁接的亲和性[13]。嫁接后第21 ~26 天,为嫁接体愈伤组织分化形成新生维管组织期。嫁接后第26 天,砧穗结合部形成层的细胞已经分化出新的导管和筛管,标志着嫁接体已成活,并已形成一个完全独立的新个体。从外观上看,此时接芽萌发,砧穗紧密连接,这与砧穗结合部愈伤组织分化形成维管组织期的显微观察结果相吻合,表明野鸦椿与圆齿野鸦椿均有较强的嫁接亲和性。因此,嫁接后第26 天之前是圆齿野鸦椿嫁接成活的关键时期。
王淑英等[23]在观察葡萄嫁接愈合过程时发现,形成层的细胞是愈伤组织产生的唯一来源。但是,杨志坚等[22]在观察油茶芽苗砧嫁接口愈合过程时发现,木本植物愈伤组织的来源不仅限于形成层的细胞,还包括髓区、木射线、皮层的细胞。周乃富等[24]的研究结果表明,核桃嫁接愈合方式不仅存在由形成层细胞主导的砧穗愈合方式,还存在由木射线细胞主导的新方式。显微观察发现,嫁接初期砧穗伤口处皮层、韧皮部薄壁细胞和木射线及形成层的细胞均可产生愈伤组织,但是,形成层的细胞在嫁接愈合过程中自始至终占主导地位。因此,在圆齿野鸦椿嫁接过程中,创造有利于愈伤组织生长和新的形成层产生的条件,是提高嫁接成活率的重要途径。
3.2 嫁接体接口部位抗氧化酶活性的变化趋势
过氧化物酶(POD)和过氧化氢酶(CAT)是植物体内重要的抗氧化酶类,具有清除接口处过量活性氧(ROS)、促进木质素合成的作用,有利于嫁接体接口的愈合[15]。有关研究结果表明,与非亲和性的黄瓜、甜瓜嫁接苗相比,亲和性的黄瓜、甜瓜嫁接苗接口处均有较高的抗氧化酶(SOD、POD、APX)活性和较低的活性氧含量[25]。胡艳青等[11]的研究结果表明,嫁接初期,CAT 活性升高,有利于清除氧自由基,起到防止过氧化伤害和清除植物体内过剩的H202的作用;嫁接体成活后,CAT 活性随之下降。试验结果表明,嫁接后第0 ~10 天,以野鸦椿为砧木、以圆齿野鸦椿为接穗进行嵌芽接,其嫁接体的CAT 活性显著升高,而此时即为解剖学上砧穗切口隔离层与愈伤组织的形成时期,说明嫁接体在初期受到创伤后,嫁接苗的抗氧化能力得到提升。嫁接后第20 天,即砧穗结合部愈伤组织分化形成新生维管组织时期,CAT 活性开始降低,此时嫁接体已经愈合,形成层细胞分化出较多结构规整的木质部和韧皮部,新的输导组织已经形成,结合部 CAT 活性呈下降趋势,这与上述研究结果基本一致。但是,曲云峰[26]研究发现,CAT 不是杏嫁接成活的关键酶,具体原因有待进一步研究。
卢善发[27]的研究结果表明,POD 是氧化吲哚丁酸的关键酶,嫁接初期POD 活性升高,有利于维持激素的平衡;王瑞敏等[28]研究了金叶银杏的扦插生根机制,结果表明,高活性的POD 能够促进插穗愈伤组织的形成;张红梅等[29]研究发现,嫁接愈合过程中,POD 活性呈升—降—升的变化趋势,POD 活性高有利于维管组织的木质化。试验结果还表明,嫁接初期,嫁接体内的POD 活性显著升高,并于嫁接后第10 天出现第1 个高峰值,此时即为愈伤组织形成时期,解剖学观察结果显示,嫁接第5 天后,砧穗愈伤组织大量分化,由此可以推测,嫁接后第5 ~10 天是愈伤组织形成和生长的高峰期;随着嫁接愈伤组织的分裂增殖,POD 活性显著降低;嫁接后第15 ~25 天,嫁接体内的POD 活性又显著升高;嫁接后第26 天,嫁接体接合部位的POD 活性最高,此时正是愈伤组织分化形成维管组织时期,POD 的活性高,表明其参与了维管组织的分化。综上所述,嫁接体结合部的POD 和CAT 活性均高,则有利于嫁接口的愈合,这与上述试验结果类似。
3.3 嫁接体接口部位苯丙烷代谢相关酶活性的变化趋势
苯丙氨酸解氨酶(PAL)和多酚氧化酶(PPO)为苯丙烷代谢相关酶,在嫁接体愈伤组织细胞形成、增殖及分化过程中发挥了重要作用[30]。
Chen 等[31]研究发现,嫁接接口处PPO 活性升高,有利于嫁接体的愈合与生长;López-Gómez等[32]的研究结果表明,PPO 主要作用于嫁接初期,表现为嫁接初期PPO 活性上升。试验结果表明,嫁接后第0 ~10 天即隔离层与愈伤组织形成时期,PPO 活性快速升高,形成了足够厚的隔离层,可以保护嫁接体免受感染,这与上述研究结果一致。但是,亦有研究结果表明,PPO 活性升高加速了接口处木质化的进程,从而使嫁接难以成活[33]。嫁接后第10 天,嫁接体内PPO 活性迅速下降并趋于平稳,而此时正是愈伤组织增殖与连接期,隔离层的消失有利于嫁接口的愈合,这一结果与张淑红等[34]的研究结果类似。
刘婧冉等[16]的研究结果表明,PAL 参与嫁接愈合过程中细胞的分化,有利于嫁接体组织的发育和功能的重建。冯金玲等[35]对油茶嫁接的试验结果表明,在嫁接初期即愈伤组织形成时期,PAL活性呈下降趋势;而在嫁接愈合完成后,PAL 活性却上升。嫁接试验中发现,在嫁接初期即愈伤组织形成期,PAL 活性降低;而在愈伤组织细胞分裂增殖与连接期,PAL 活性升高。这一结果表明,此时砧、穗间细胞分化旺盛,有利于木质部的形成,这与上述研究结果类似,但与黄蔓娜等[12]在对梨树进行嫁接试验时发现的嫁接后第18 天砧、穗形成层连接后其PAL 活性快速增加的结果不同。
试验结果表明,PPO 和PAL 酶活性在嫁接体愈合过程中的变化趋势正好相反。嫁接初期,PPO活性升高,而PAL 活性降低,这有利于嫁接体接口愈伤组织的形成。
目前,关于圆齿野鸦椿嫁接愈合过程的生理生化机制、解剖学等嫁接机理方面的研究报道较为鲜见。本研究对圆齿野鸦椿嫁接愈合过程进行了解剖学观察,并分析了嫁接体的生理生化特征,但因受场地、苗量等因素的影响,未设置自根苗与嫁接苗的酶活性的比较试验,故试验结果存在一定的局限性;对嫁接愈合过程的细胞学观察,还需结合嫁接接口愈合分子机制、基因组学和蛋白组学等方面的研究结果来分析,以优化并完善圆齿野鸦椿嫁接繁殖技术体系。
4 结 论
本研究以野鸦椿实生苗为砧木、以圆齿野鸦椿当年生枝上健壮的腋芽为接穗进行嵌芽接试验,观察了砧穗愈合过程的解剖学形态特征,测定并分析了嫁接后不同时期其结合部的相关酶活性的变化情况,得出了如下结论。
1)嫁接体的愈合过程可划分为3 个时期,即砧穗切口隔离层及愈伤组织形成期(嫁接后第0 ~10 天)、砧穗结合部愈伤组织增殖与连接期(嫁接后第11 ~20 天)、砧穗结合部愈伤组织分化形成维管组织期(嫁接后第21 ~ 26 天)。
2)嫁接体在嫁接后第26 天形成了新的导管和筛管,砧穗间细胞能够正常接触和流通,输送养分和水分,形成了一个新的个体,说明野鸦椿与圆齿野鸦椿均有较强的嫁接亲和性。
3)嫁接愈合过程中,嫁接体内酶活性的变化趋势为:嫁接初期,POD、CAT 和PPO 活性均升高而PAL 活性降低,这有利于嫁接体接口的愈合;而嫁接体愈合后,PAL 活性降低而PPO 和POD 活性均升高,这可促进嫁接体维管组织的形成。
猜你喜欢
现代园艺(2022年11期)2022-06-09
河北果树(2021年4期)2021-12-02
中国生态农业学报(中英文)(2021年8期)2021-07-28
中国果业信息(2020年12期)2020-12-15
林业科学(2020年10期)2020-11-30
西藏人文地理(2020年4期)2020-11-19
农业工程技术·综合版(2020年1期)2020-04-23
铁道通信信号(2019年6期)2019-10-08
铁道通信信号(2018年1期)2018-06-06
中国卫生标准管理(2015年1期)2016-01-14
