
水体升温和生态情境对异育银鲫群体行为的影响
2022-12-16 04:50李武新马辉煌于长暄雷家鸣曾令清
生态学报 2022年22期
李武新,马辉煌,于长暄,刘 霜,雷家鸣,曾令清
重庆师范大学进化生理与行为学实验室,动物生物学重庆市重点实验室, 重庆 401331
水温是影响自然界鱼类等水生动物生理生化过程、行为表现甚至生活史对策的重要生态因子[1—2]。自然水体包括两种水温变化类型,即缓慢温度变化(如季节更替)和急性温度变化(如潮汐带、洄游和洪水期)[3—4]。目前日益严重的全球气候变暖和人类干扰自然活动等潜在改变自然水体的温度变化模式。研究发现,气候异常引起的温度骤然变化、水利设施导致的水温分层及热(核)电厂等的温排水常导致鱼类面临急性变温的逆境,对鱼类的繁殖、生长发育、摄食和避敌等活动造成一定的负面影响[5—7]。
群体行为是指因社会原因(如繁殖、迁徙)而使动物个体聚集一起的现象[8]。动物群体的行为特征主要包括凝聚力和协调性,前者的评价指标包括个体间距离[9]和最近邻距离等[9—10],后者的评价指标包括个体游泳速度同步性和群体极性等[9, 11—12]。研究发现,集群有助于增加个体摄食效率、繁殖成功率和游泳效率,及降低被捕食风险等生态收益[13—14],但也伴随寄生虫感染及食物、配偶等资源竞争增强的风险代价[13, 15—16]。集群的收益与代价是处于动态变化过程,与个体状态和环境条件(如食物资源、温度变化)等因素有关[17],进而影响群体的结构(凝聚力和协调性)和功能[18]。据此,本研究关注的第一个科学问题是急性水体升温是否导致鱼类的群体凝聚力和协调性下降。
群体行为是群体中的个体成员通过一定的社交规则而呈现的自组织结构特征[11, 17],然而群体特征并非一层不变,将随栖息环境的改变而发生相应调整[19—20]。例如,当动物群体在面对捕食者时群体表现更为紧密,而当动物群体在自由摄食时表现更为松散[21—22],即群体凝聚力的变化与外部环境有关。自然界动物的栖息生境不仅类型多样、复杂多变,而且生存资源(如食物,隐蔽所)及捕食者等具有明显的时空分布异质性。此外,前期的群体行为研究主要关注单一的生态情境[12,19],而不同生态情境对同一动物群体影响的研究仍然较为匮乏[20]。据此,本研究关注的第二个科学问题是鱼类的群体凝聚力和协调性是否在不同生态情境下维持稳定。
基于上述研究背景,本研究推测水体快速升温将导致鱼类个体的生理(如能量消耗增加)和行为(如活跃性增强)发生变化,这些个体水平的生理行为改变可能导致群体特征发生相应变化,并且群体特征的改变可能具有生态情境依赖性。为考察不同水体升温速率和生态情境类型对鱼类群体行为的影响,本研究以喜集群鲤科鱼类异育银鲫(Carassiusauratusgibelio)幼鱼为实验对象,考察在3个温度(恒温16℃、慢速升温及快速升温10℃)和3种生态情境(开放水域、食物及食物与隐蔽所)条件下实验鱼群体行为(如个体游泳速度及其同步性、个体间距离、最近邻距离、极性及鱼群吃饵比例等)的变化特征,为鱼类群体行为、群体决策及热(核)电厂温排水的科学排放等相关研究提供重要参考资料。
1 材料与方法
1.1 实验鱼及其驯化
异育银鲫(Carassiusauratusgibelio)购于重庆市北碚区歇马人工养殖基地,运回后在重庆师范大学进化生理与行为学实验室2个循环控温水槽(约250 L)驯化1个月以上。在每个水槽底部安置一定数量的人工水草,以模拟隐蔽所及提高驯化水体的环境丰富度。驯化期间,每天10:00用红线虫对鱼饱足投喂1次。为减少充氧对鱼群摄食的影响,在每次投喂前10 min关闭充气泵以使水面平静。投喂1 h后,用虹吸管清除水中残饵和粪便以维持水体清洁,但导致驯化水体的减少,随后以曝气4 d的自来水补充水体(换水量约10%)。溶氧水平维持在7 mg/L以上,水温控制在(16.2±0.1)℃,光周期设定为14 L∶10 D。
1.2 实验设计
驯化结束后,挑选大小相近、体格健康的异育银鲫(体重(3.96±0.03) g,体长(5.59±0.03) cm,n=144)作为实验对象。前期研究发现,一个由4尾鱼组成的鲤科鱼群已表现出明显的集群行为[23—24],并且由于安置单个鱼群的养殖单元格存在空间限制,因此本研究采用4尾鱼组成1个鱼群。水温的上升幅度设定为10℃。本研究包括3个温度处理组:(1)对照组(恒温在16℃左右);(2)慢速升温组(16→26℃,升温速率为2℃/d);(3)急速升温组(16→26℃,升温速率为10℃/d)。本研究采用3个多单元格循环控温水槽用于安置每个温度处理组的鱼群,每个水槽包括30个养殖单元格(长17 cm×宽10 cm×深15 cm)。每个单元格只放置一个鱼群,其上方置有网格盖子并以重物压紧,以防止鱼跳出。此外,本研究还设置3种生态情境:开放水域环境、食物环境、食物+隐蔽所环境(见后)。每个温度处理组包括12个鱼群,在升温前(测定Ⅰ)后(测定Ⅱ)采用一个自制的圆形水槽(直径100 cm,水深8 cm)拍摄鱼群的群体行为(详细方法见后)。本研究严格遵守国家动物福利伦理与保护等相关规章制度。
1.3 实验方法与参数计算
1.3.1群体行为
群体行为的观测装置为一个直径100 cm、壁高20 cm的圆形塑料水槽(水深8 cm),其内壁和底部均为白色,放置于直径2 m的大圆形水槽,通过大水槽的水体精确控制观察水槽的水温。在大水槽的四周安装1.5 m米高的纯绿色布帘,为鱼类群体行为的真实表现提供稳定的环境背景并消除外部潜在影响。在观测水槽的正上方置有一个连接电脑的广角高清摄像头(Webcam Pro9000,中国罗技公司,15帧/s),用于拍摄鱼群在3种生态情境(次序:开放水域→食物→食物+隐蔽所)中的群体行为。生态情境的测定次序设定参照前期的相关研究[9]。每个鱼群于第2 d重复1次上述群体行为的拍摄工作,以提高群体行为的可靠性。实验期间,所有环境条件(如光强均匀,水温恒定,安静等)维持稳定。
本研究采用的3种生态情境包括开放水域环境、食物环境、食物+隐蔽所环境[9,23]。其中,(1)开放水域环境是指圆形水槽无食物和隐蔽场所,但具有空的3个食物盘(玻璃培养皿,直径6 cm)。为减少空间异质性,食物盘以等边三角形固定在圆形水槽的底部边上,其四周用不透明胶带环绕,在每个食物盘中安置九宫格以增加鱼群发现食物的难度。(2)食物环境是指随机在食物盘的九宫格中放置4个红线虫,3个食物盘共放12个红线虫,此食物数量的设定是为减少鱼因摄食消化引起能量代谢上升对个体游泳能力产生的潜在影响。红线虫的添加办法:通过注射器及透明导管(内径5 mm)将红线虫轻轻注入食物盘中,此过程中鱼群无法看到外部的实验人员。(3)食物+隐蔽场所是指圆形水槽同时包括食物和隐蔽场所,隐蔽场所是由固定在同一底板的5棵塑料水草组成。在食物环境的群体行为拍摄之后,轻轻将水草放置于圆形水槽中间底部以便于摄食后鱼群进入该隐蔽场所。
行为拍摄前,对鱼群进行禁食48 h以排除食物消化吸收的影响并维持一定的食欲。集群行为的实验步骤:首先,将每个鱼群从养殖单元格轻轻地转移至圆形水槽中适应10 min,即在开放水域环境适应,然后用摄像设备拍摄12 min。其次,在每个食物盘快速添加4个红线虫(通常在1 min内完成,以减少对鱼群的干扰),之后拍摄12 min。最后,由于鱼群已摄取数量不等或全部的红线虫,需先补充红线虫的总数量至12个,每个食物盘仍维持4个红线虫,同时在圆形水槽中间放置水草,随后拍摄12 min。所有行为拍摄之后,鱼群被轻轻转移至其单元格,并适当投喂1次红线虫。在每个鱼群的3个生态情境全部拍摄完后,将圆形水槽的水体全部清除,再更换充分暴气后的等温自来水,用于下一个鱼群的群体行为拍摄。更换水体的目的是消除水体存留的化学信息素(如食物信息素、化学预警物质)对下一个鱼群的群体行为的潜在影响。当水温升至目标温度时,再次按上述实验步骤拍摄3个温度处理组的所有鱼群在升温后的群体行为。
1.3.2参数计算
本研究采用下列实验参数评价异育银鲫的个体和群体特征,前者包括个体游泳速度(Individual swimming velocity,V)及其同步性(Synchronization of velocity,Sv),后者包括个体间距离(Inter-individual distance, IID)、最近邻距离(Nearest neighbor distance, NND)、群体极性(Polarization,P)。先用格式工厂软件将视频WMV格式转化成AVI格式,随后将AVI视频导入图像跟踪软件idTracker进行视频分析[25],获取每尾鱼在视频中每帧的坐标数值。根据视频像素与观测水槽大小的换算比例(约15.3像素/cm)计算下列实验参数。为消除转运、添加食物或水草等操作可能对鱼群产生影响,舍弃前2 min的视频,统计分析只采用3—12 min的轨迹数据(10 min),实验参数的计算公式如下:
(1) 个体游泳速度
(1)
式中,V为个体游泳速度(cm/s),xt和xt-1分别为单尾鱼在t、t-1时刻的横坐标值;yt和yt-1分别为单尾鱼在t、t-1时刻的纵坐标值;dt为两个坐标点的时间间隔(1/3 s)。
(2)个体游泳速度同步性
Sv是评价个体游泳速度的同步性,介于0—1之间。如果该数值越接近1,那么个体游泳速度的同步性就越高[26]。
(2)
式中,Sv为个体游泳速度同步性(无单位),vi和vj为鱼群中第i、j鱼在同一时间的瞬时游泳速度。
(3)个体间距离
IID表示单个鱼群中所有个体之间的平均距离(cm),评价鱼群的群体凝聚力。
(3)
式中,IID为个体间距离(cm),xi和xj为鱼群中第i、j鱼在t时刻的横坐标值,yi和yj为鱼群中第i、j鱼在t时刻的纵坐标值。
(4)最近邻距离
NND表示鱼群中某一个体与其它所有个体之间距离中的最短距离(cm),衡量鱼群的群体凝聚力。
(4)
式中,NND为最近邻距离(cm),xi和xj为鱼群中第i、j鱼在t时刻的横坐标值,xi和xj为鱼群中第i、j鱼在t时刻的纵坐标值。
(5)群体极性
极性(P)表示鱼群个体排列的整齐程度(无单位),其数值介于0—1之间。当鱼群中所有个体以绝对一致方向排列时,P值为1;当鱼群中所有个体以方向相反且完全抵消的方式而排列时,P值为0[12, 27]。
(5)
式中,vi(t)为单尾鱼i的单位时间内的运动向量,运动方向是从t-1时刻的坐标点指向t时刻的位置坐标点。
(6)鱼群摄饵比例(Percentage diet fed by fish shoals,Pdiet)
Pdiet表示在视频拍摄时间内单个鱼群摄取红线虫的百分比(%),该数值介于0—100%。
(6)
式中,D表示单个鱼群在单位时间内实际摄取红线虫的数量。
1.4 统计分析
实验数据先采用Excel(2010)进行常规计算后,再用 SPSS(17.0)软件进行相关统计分析。采用三因素方差分析考察不同时间、温度及生态情境对个体游泳速度及其同步性、个体间距离、最近邻距离以及二者之比、群体极性和鱼群摄饵比例的影响,如存在显著差异再采用Duncan多重比较或配对T-检验分析各组间的显著性差异。采用组内相关分析(Intraclass correlation coefficient, ICC)考察所有实验参数在不同时间和环境的重复性,ICC介于0—1之间,该值越大,表型的重复性就越高。采用Pearson相关分析考察集群行为特征的表型关联。所有数据均以平均值±标准误(Mean±SE)表示,显著水平定为P<0.05。
2 结果
2.1 升温和环境对异育银鲫集群行为的影响
不同升温速率对异育银鲫个体游泳速度具有显著的影响。快速升温导致该种鱼的个体游泳速度明显增加,而对照组和慢性升温均未呈现此效应(图1,表1,P<0.05)。不论快速升温还是慢速升温均未影响个体游泳速度同步性(图1,表1,P>0.05)。此外,不同生态情境对个体游泳速度及其同步性产生影响。个体游泳速度及其同步性均随环境异质性的增加而呈下降趋势(图1,P<0.05)。
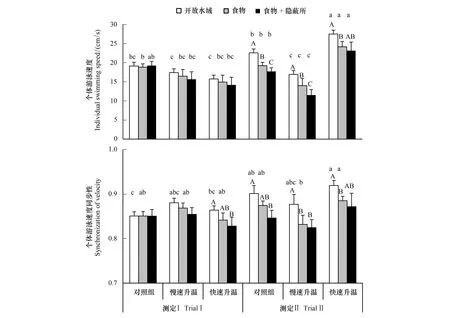
图1 升温速率和生态情境对异育银鲫个体游泳速度及其同步性的影响
三个温度处理组中异育银鲫的个体间距离和最近邻距离均未受升温和生态情境的影响,但不同处理组间最近邻距离与个体间距离的比例具有差异(图2,表1,P<0.05)。虽然该种鱼的极性不受升温影响,但具有生态情境依赖性,三个温度处理组的极性均呈下降趋势(图3,表1,P<0.05)。不论升温组还是对照组,鱼群摄饵比例在测定Ⅱ中明显提高(P<0.05),但不受生态情境的影响(图4,表1,P>0.05)。

表1 三因素方差分析的统计结果

表2 异育银鲫集群行为特征的重复性
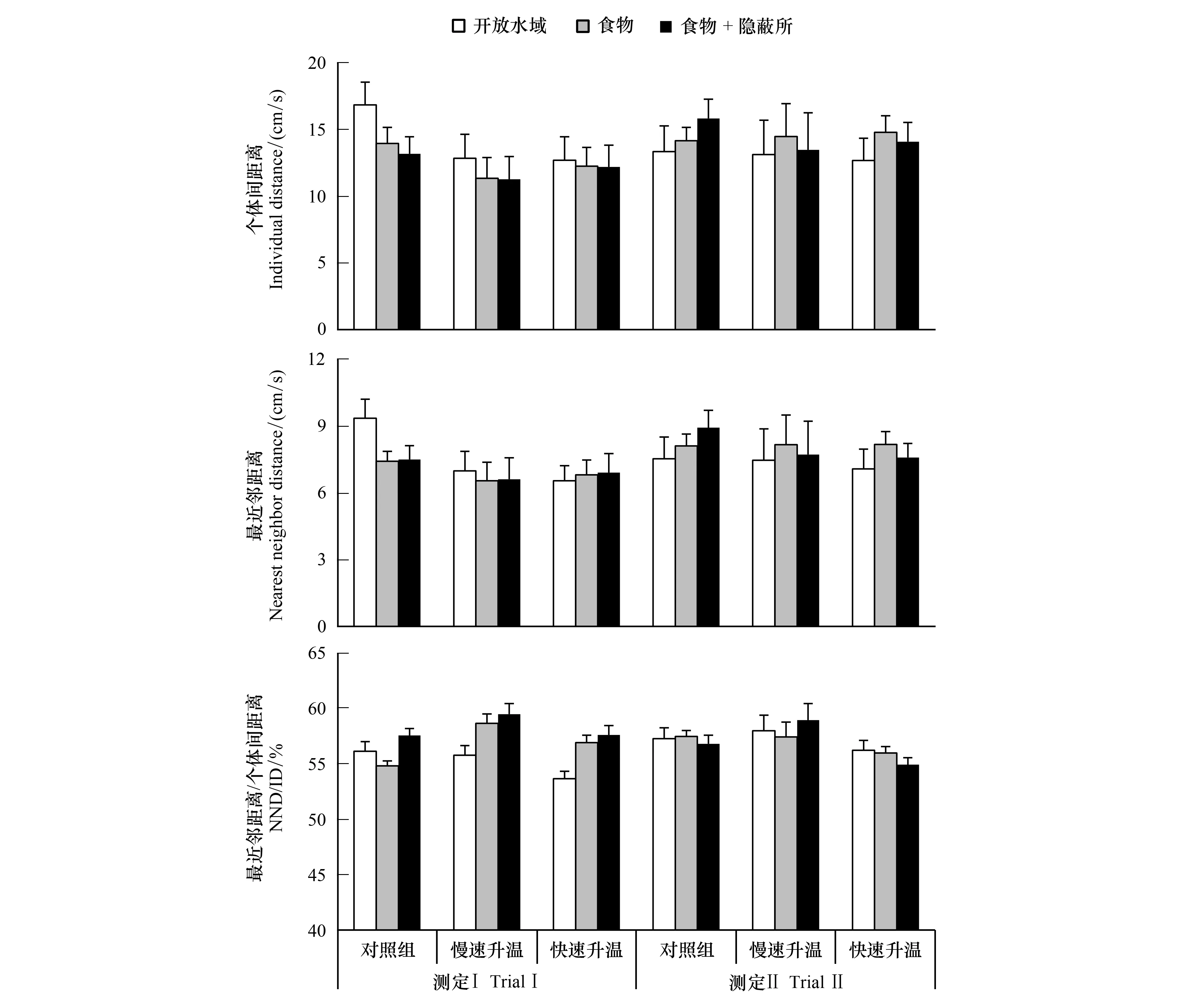
图2 升温速率和生态情境对异育银鲫个体间距离和最近邻距离及二者之比的影响
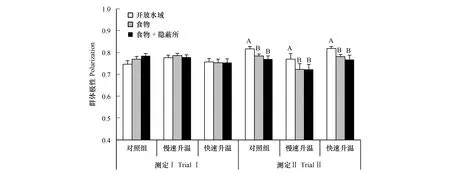
图3 升温速率和生态情境对异育银鲫群体极性的影响

图4 升温速率和生态情境对异育银鲫鱼群摄饵比例的影响
2.2 表型关联
异育银鲫集群行为中不同实验参数具有表型关联(表3,所有P<0.01)。其中,个体游泳速度与其同步性、个体间距离、最近邻距离以及极性均呈正相关;个体游泳速度同步性与个体间距离、最近邻距离呈负相关,与极性呈正相关;个体间距离与最近邻距离呈正相关,二者均与极性呈负相关。
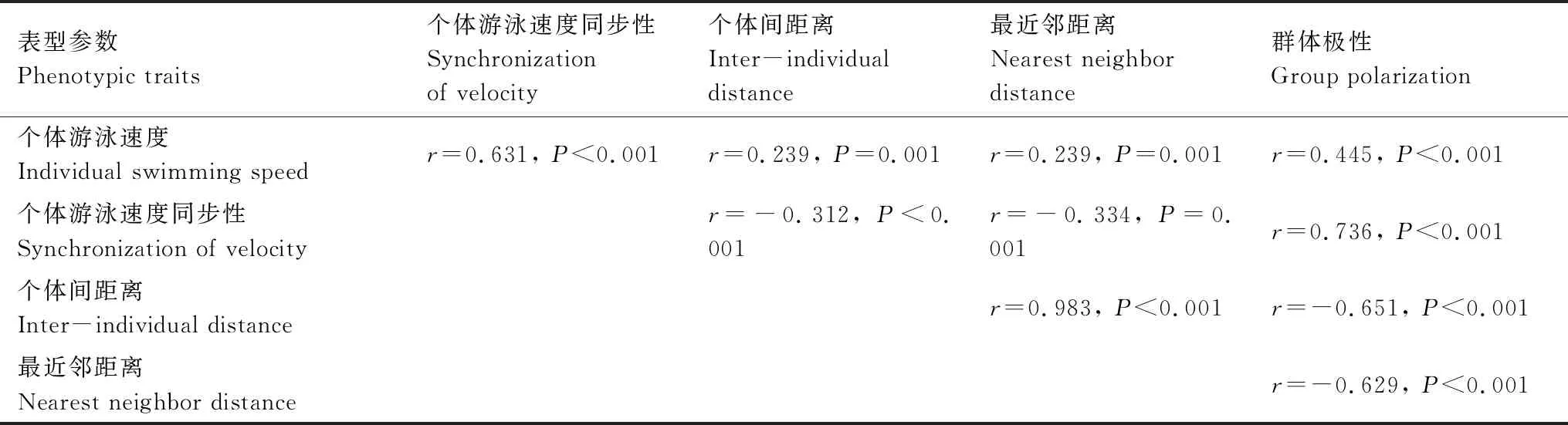
表3 异育银鲫集群行为的表型关联
3 讨论
本研究目标是考察水体升温及生态情境类型对异育银鲫幼鱼群体行为的影响。结果显示:在个体水平上,快速升温导致个体游泳速度显著增加,但恒温组(对照)和慢速升温的个体游泳速度均未变化;个体游泳速度同步性不受升温速率的影响,可能与较高的个体游泳速度有关。然而,个体游泳速度及其同步性随生态情境复杂性的增加而呈下降趋势。在群体水平上,个体间距离和最近邻距离均未受升温和情境类型的影响,表明异育银鲫群体凝聚力在升温前后和不同情境间维持稳定。该种鱼的群体极性不受升温影响,但随情境复杂性的增加呈现下降趋势,表明异育银鲫群体协调性在较大的温度变化后仍维持原来的群体排列,生态情境复杂性的增加却降低群体排列的整齐性。虽然鱼群摄饵比例的提高与升温及生态情境均无关,但实验鱼的个体和群体特征在升温前后及不同生态情境中仍具有重复性。
3.1 水体升温对个体和群体特征的影响
一般而言,鱼类个体游泳速度随水温的升高呈现先上升后下降的变化趋势[28—30],存在游泳速度的最适温度,但此温度-游泳速度关系与水体的升温模式(如长期驯化与急性变化)密切相关。本研究发现,慢速升温对异育银鲫群体中的个体游泳速度并无影响,但急速升温明显提高个体游泳速度,与三角洲胡瓜鱼(Hypomesustranspacificus)的研究结果相似[31]。鱼类游泳速度的增加与机体代谢空间[32]、肌肉线粒体代谢(如氧化磷酸化)[33]等能力的提高有关。此外,在体型大小、水温相近及相同尾数条件下,升温后异育银鲫的个体游泳速度(12—28 cm/s)明显高于青鱼(Myloppharyngodonpiceus)(0—10 cm/s)[18]、麦穗鱼(Pseudorasboraparva)(0—10 cm/s)[34]、中华倒刺鲃(Spinibarbussinensis)(约7 cm/s)和鳊(Parabramispekinensis)(约4 cm/s)[35]。这种差异的可能原因是因为实验室前期研究的水温驯化时间较长(如4 w及以上),而本研究的升温过程是短期(1或5 d)完成,并未使实验鱼在达到目标水温后经历较长的水温驯化时间,导致个体遭遇了较严重的升温胁迫而无法及时调整机体的生理状态以适应升温后的水体环境,而表现出较强的自发游泳运动。
水温不仅影响异育银育的个体游泳速度,而且还影响其群体结构特征。与预测相反,本研究发现不论慢速升温还是急速升温对异育银鲫群体的个体间距离、最近邻距离和群体极性均未产生影响,即该种鱼的群体凝聚力和协调性在急性升温之后仍然维持相对稳定。本研究还发现,不论恒温组还是升温组的大部分鱼群紧靠圆形水槽边缘,进行较长时间的绕环游泳运动,表现出较强的活跃性且较整齐的群体队列。这种群体运动模式可能是水体的急性升温并未导致异育银鲫凝聚力和协调性发生改变的重要原因。虽然本研究未计算群体运动的时间比例,但上述群体运动的行为特征也可能暗示,异育银鲫在水体的急性升温后还处于不良适应阶段。然而,其它鱼类如黄线狭鳕(Theragrachalcogramma)的最近邻距离,以及鳟(Salmotrutta)和三角洲胡瓜鱼的个体间距离及最近邻距离均随水温的上升而呈增加的变化趋势[31,36—37],表明高温环境导致这些鱼类群体结构变得更为松散,群体凝聚力明显下降。理论上分析,在较高水温环境中鱼类增强机体的能量代谢水平[38]、活跃性[39]和个体食欲[40],通过增加个体游泳速度(自发活动)的方式可在单位时间内扩大觅食空间[31]。然而,为减少个体间对食物和隐蔽所等环境资源的竞争,目标导向的个体运动可能更多关注环境中的可利用资源(如食物)而非个体间距离的紧密维持,导致群体结构的个体间距离及最近邻距离增加[21,41],可能降低群体运动的协调性。
3.2 生态情境对个体和群体特征的影响
有综述指出大多数群体行为的观察是在单一的较高均质生态情境完成的[19],而多种异质生态情境的研究正逐渐吸引研究人员的关注[20]。这是因为自然界动物短期内经常经历的复杂多变且高异质性的生态情境,个体和群体结构特征在不同生态情境中将产生适应性的改变或调整。本研究共设置三种生态情境类型(简单到复杂),发现个体游泳速度及其同步性和群体极性从开放水域到食物环境,再到食物+隐蔽所环境呈现下降的变化趋势,与三刺鱼(Gasterosteusaculeatus)及中华倒刺鲃的研究结果相似[20, 23]。然而,个体间距离和最近邻距离却未随情境复杂性的增加而发生变化,这些相异结果表明生态情境的复杂性降低异育银鲫的群体协调性,但不影响其群体凝聚力。相比较于开放水域情境,鱼群发现环境出现食物后可能放慢游泳速度而各自分散寻找食物,降低了个体间对食物的竞争,在此生态情境下群体显得较为松散[42],理论上觅食情境中个体游泳速度(如时快时慢)的变异程度可能较大。当生态情境同时存在食物及隐蔽所(水草)时,鱼群出现部分个体(平均1.5尾)进入隐蔽所并停于其中一段时间,或者整个鱼群进入隐蔽所。一段时间后部分个体率先游出隐蔽所,剩余个体随后再游泳出,这些情况常导致鱼群分裂为两个亚群,降低个体游泳速度同步性和群体极性。虽然本研究并未设置捕食者情境,但有关研究发现排列整齐的鱼群的被捕风险明显低于排列不齐的鱼群[43],环境中的捕食者信息素可引发鱼群运动变慢且更加紧凑[22]。因此,鱼类可能会在评估资源利用与捕食风险之间的权衡后及时调整群体行为。
3.3 群体个性与表型关联
个性是指动物个体间的行为差异在不同时间或环境中稳定维持,包括勇敢性,探索性,活跃性,好斗性及社会性等5个行为特征[8, 44]。近期研究开始关注动物(如蜜蜂、蜘蛛、鱼类等)不同群体之间稳定的集群行为差异(Collective personality,称为群体个性),这种差异同样在较长时期或不同环境仍维持稳定[20, 45—47]。虽然本研究发现三个温度处理组的群体特征在升温前后和不同环境条件下保持较高的重复性,但两个升温组群体特征参数如个体游泳速度、个体间距离及最近邻距离的变异程度整体上大于对照组。进一步分析,两升温组中不同群体间实验参数(如个体游泳速度)的平均值差异可达3—5倍并且在升温前后及不同生态情境中稳定存在,即有些鱼群比其它群体具有更快的游泳运动速度,更弱的凝聚力和更高的群体极性,反之亦然,表明异育银鲫在群体水平上具有稳定的群体行为差异(即群体个性)。产生该现象的可能原因:在鱼群随机分组之后,所有个体均驯养于环境条件一致的养殖单元格之中,有助于群体成员彼此相互熟悉,使群体成员之间产生较高的成员熟悉度和拥有相似的社会经历。因此,群体成员可能会根据个体间在生理和个性等的差异而形成特定的社交互动规则,可区别于其它群体。除此之外,个体间的社交互动规则所产生的社会反馈也可能有助于形成群体个性[20]。
研究发现,三刺鱼的群体运动速度与个体间距离及极性均呈正相关,个体间距离与极性也呈正相关,即群体运动速度越快,个体间距离越大,群体队列越整齐[9];斑马鱼(Daniorerio)的游泳速度越快,其群体越松散,即个体间距离和最近邻距离随着游泳速度的增加而变大[12]。与上述研究发现类似,异育银鲫也具有个体特征和群体特征的表型关联。本研究发现,异育银鲫的个体游泳速度越快,其同步性和极性同样越高,说明快速运动可形成同步性高和排列整齐的群体运动,反之则形成个体速度和运动方向参差不齐的聚群特征。然而,异育银鲫群体的个体间距离越大,其最近邻距离相应越大,群体极性反而越低,即群体凝聚力与协调性之间可能存在某种权衡,但该现象是否普通存在于其它鱼类之中值得关注。
综上所述,本研究发现水体的快速升温影响异育银鲫的个体特征,但不影响其群体凝聚力和协调性。然而,随生态情境复杂度的增加,异育银鲫的个体游泳速度及其同步性和群体极性均下降,而群体凝聚力维持稳定。异育银鲫的鱼群存在群体个性现象及在个体水平和群体水平的表型关联。鱼群既会经历较单一均质环境,也会遭遇生态收益与风险代价并存的复杂异质环境,而其及时准确调整的群体行为策略(如缓解凝聚力与协调性的利益冲突)是在不同生态情境下的适应性结果,可能有助于提高群体中个体适合度。自然界鱼类的栖息环境多种多样且复杂多变,受人类干扰的程度和影响日益严重,使得自然水域的生态作用更加复杂多变且难以预测。未来研究仍然需要持续关注栖息环境的各种生态因子(如低氧、海洋酸化、水体污染、食物资源、栖息地变化等)对鱼类群体行为的影响,并探讨其中的内在机制,为人类活动的生态风险评价及鱼类资源保护提供重要参考。
猜你喜欢
廊坊师范学院学报(自然科学版)(2021年2期)2021-09-10
水下无人系统学报(2021年2期)2021-05-15
电子制作(2019年23期)2019-02-23
中外文摘(2017年19期)2017-10-10
Coco薇(2017年8期)2017-08-03
金融与经济(2017年6期)2017-07-19
故事作文·高年级(2017年6期)2017-06-29
中国篆刻·书画教育(2017年5期)2017-06-08
红蜻蜓·低年级(2015年8期)2016-02-02
湖南大学学报·自然科学版(2014年3期)2014-12-30
