
藏东南川滇高山栎天然林的种内与种间竞争指数的海拔差异*
2022-12-21 11:04姚慧芳曾加芹罗大庆张新生于德水
林业科学 2022年8期
姚慧芳 卢 杰 曾加芹 罗大庆 张新生 王 超 于德水
(1.西藏农牧学院高原生态研究所 林芝 860000; 2.西藏高原森林生态教育部重点实验室 林芝 860000; 3.西藏林芝高山森林生态系统国家野外科学观测研究站 林芝 860000; 4.西藏农牧学院资源与环境学院 林芝 860000)
林木生长受许多生物和非生物因素的制约,竞争可在很大程度上控制林分结构和生长发育(罗梅等, 2018),作为一种重要影响因子而备受研究者关注。林木竞争是指相邻树木之间为自身的生存生长而争夺环境资源和生存空间的现象,也是植物的种内与种间关系的主要形式之一(汪清等, 2021),对种群动态、群落结构组成和稳定性维持具有重要作用(Thorpeetal., 2010; Malekietal., 2015)。因此,探究同种与不同种之间的竞争关系及其对植物生长发育的影响,对阐明群落物种多样性和结构变化有重要意义(段光爽等, 2020)。
川滇高山栎(Quercusaquifolioides)属山毛榉科(Fagaceae)栎属(Castanea),为我国特有树种,广泛分布于云、贵、川、滇、藏等地,海拔2 400~3 900 m,最高可达4 000 m(杨益帆等, 2020)。该种是亚高山硬叶栎林的主要树种,分布在干旱山坡或山顶处常呈灌木状,环境适应力较强,可在较恶劣环境中生存(王奥等, 2017; Wangetal., 2018)。在西藏,川滇高山栎林大部分为纯林,分布在阳坡或半阳坡,也有与白桦(Betulaplatyphylla)、急尖长苞冷杉(Abiesgeorgeivar.smithii)、林芝云杉(Picealikiangensisvar.linzhiensis)、华山松(Pinusarmandii)等混交的复层林。目前,有关川滇高山栎的研究报道主要在生理生化特性(黄晓霞等, 2015)、群落植物特征(杨朗生等, 2017)、遗传多样性(程小毛等, 2017)等方面,还未见到关于西藏川滇高山栎群落竞争关系的研究。鉴于此,本研究以西藏林芝市巴宜区鲁朗镇的川滇高山栎天然林为对象,通过设置样地、采集数据和运用Hegyi竞争模型,定量研究川滇高山栎林的种内与种间竞争指数,揭示竞争力较强的树种,并阐明林分的种内与种间竞争的海拔差异,以期为川滇高山栎天然林的经营和保护提供理论参考。
1 研究区概况
研究区位于西藏林芝市巴宜区鲁朗镇,平均海拔3 385 m。由于印度洋水汽从雅鲁藏布江大峡谷北上,使这里的气候较湿润,年降水量约650mm,年均气温8.7℃,属高原温暖半湿润气候,素有“西藏江南”美称。该区域土壤类型为山地暗棕壤,植被类型丰富,乔木层有川滇高山栎、急尖长苞冷杉、林芝云杉、糙皮桦(Betulautilis)、白桦等; 灌木层有三花杜鹃(Rhododendrontriflorum)、白背杜鹃(R.leucaspis)、黑茶藨子(Ribesnigrum)、冰川茶藨子(R.glaciale)、柳叶忍冬(Loniceralanceolata)、陇塞忍冬(L.tangutica)、越桔叶忍冬(L.myrtillus)、粉叶小檗(Berberispruinose)、腺齿蔷薇(Rosaalbertii)、野丁香(Leptodermispotanini)和西南花楸(Sorbusrehderiana)等; 草本层有五裂蟹甲草(Parasenecioquenquelobus)、光蔓茎堇菜(Violadiffusoides)、腋花扭柄花(Streptopussimplex)、防己叶菝葜(Smilaxmenispermoidea)、宽叶兔儿风(Ainsliaealatifolia)、落芒草(Oryzopsismunroi)、凉山悬钩子(Rubusfockeanus)、假鹤虱(Eritrichiumthymifolium)、酢浆草(Oxaliscorniculate)、堇花唐松草(Thalictrumdiffusiflorum)、高山露珠草(Circaeaalpine)和近多鳞鳞毛蕨(Dryopteriskomarovii)等。
2 研究方法
2.1 样地调查
在野外实地调查基础上,根据川滇高山栎林具体分布情况,采用典型抽样调查法,按海拔梯度设置4个50 m×100 m的川滇高山栎林样地。记录样地基本信息(经纬度、海拔、郁闭度、林分类型等)。参照高浩杰等(2017)以及罗梅等(2018)的方法,对样地内胸径≥5cm的乔木每木检尺,记录其位置、胸径、树高、冠幅和枝下高(表1),统计各样地的树种组成及所占比例(表2)。在各样地的对角线上分别设置9个5 m×5 m样方和17个1 m×1 m小样方,分别调查样方和小样方内的灌木和草本种类。

表1 样地概况

表2 各样地的树种组成及比例
2.2 竞争范围及径级结构确定
依据不同样地的树冠接触面积和遮荫状况、林窗半径、上层树影响范围等指标的差异,确定川滇高山栎的林窗半径在5 m左右。为消除边缘效应的影响,在样地边缘设置5 m宽的缓冲带,缓冲带的任何乔木只作为竞争木,缓冲带以内为核心区。采用镜像、偏移和线性膨胀方法(Radtkeetal., 1998)在核心区内选取部分川滇高山栎为对象木,以对象木为中心,将在其半径5 m的样圆内所有胸径≥5cm的乔木都视为竞争木(曾思齐等, 2016)。图1以样地b为例说明川滇高山栎天然林内的林木分布和缓冲带设置。
按测得树木胸径大小范围,以5cm为径阶距,可划分出5~6个径级,则第Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ径级的胸径变化范围分别为5~10、10~15、15~20、20~25、25~30、≥30cm。统计各径级的树木数量。
2.3 竞争指数计算及函数回归模型
Hegyi竞争模型具有应用简便性,所以常被用于竞争关系分析。首先,根据对象木和竞争木的坐标,利用式(1)计算二者之间的距离(刘万生等, 2020):
(1)
式中:Lij为竞争木和对象木之间的距离(m);xi,yi为对象木i的坐标值(m);xj,yj为竞争木j的坐标值(m)。然后,采用Hegyi于1971年提出的单木竞争模型,公式(2)和(3),量化川滇高山栎的种内与种间竞争指数:
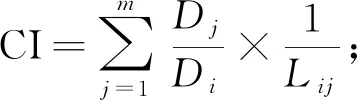
(2)
(3)
幂函数回归模型:
CI=aDBH-b。
(4)
式中: CI为竞争强度指数,DBH为个体胸径,a和b为模型参数。
三次多项式函数回归模型:
CI=cDBH3+dDBH2+eDBH+f。
(5)
式中: CI和DBH表示的含义与上式相同,c,d,e,f为模型参数。
指数函数回归模型:
CI=gehDBH。
(6)
式中:g,h为模型参数,e为常数,近似等于2.718281828。
2.4 数据分析
所有统计和计算都利用Excel 2019软件进行。采用Origin 2019b软件绘制样地内林木分布图,SPSS19.0软件拟合竞争指数与对象木胸径的关系并绘制相关图表。
3 结果与分析
3.1 川滇高山栎天然林林分基本特征因子
各样地核心区内的对象木和竞争木概况见表3。对象木和竞争木的株数均随海拔降低而增加,但胸径和树高随海拔的变化没有明显规律。对象木的最大胸径随海拔升高先减后增,在样地d(海拔3 440 m)最大; 竞争木的最大胸径则在样地b最大(海拔3 640 m)。

图1 样地b内的林木分布及缓冲带设置

表3 对象木与竞争木概况
3.2 川滇高山栎种内与种间竞争
在样地a、b、c、d中,川滇高山栎对象木的种内竞争指数分别为16.236、23.530、45.887和119.373(表4),分别占总竞争指数的17.11%、22.26%、30.79%和70.13%。样地d的川滇高山栎以种内竞争为主,可能是由于海拔相对低,川滇高山栎可长为大树,且是阔叶树种,对阳光、水分等资源的竞争力较强,而急尖长苞冷杉、林芝云杉等针叶树占据着不利地位,样地d的川滇高山栎树木数量明显增加,在群落中占绝大部分,因此川滇高山栎的竞争主要来自种内。
调查样地中对川滇高山栎形成竞争的其他树种主要有急尖长苞冷杉、林芝云杉、白桦和糙皮桦。样地a、b、c、d的种间竞争指数分别为78.682、82.187、103.123、50.855,占总竞争指数的比例分别为82.89%、77.74%、69.21%、29.87%,表现为随海拔降低而不断减小。在样地a、b、c中,川滇高山栎的种间竞争指数明显大于种内竞争指数,且样地a的种间竞争指数所占比例最大,这是由于在海拔相对高(3 540~3 740 m)的样地内川滇高山栎的生长形态不同于较低海拔地区,表现为从根部分叉成多枝,无主干且叶面积较小,树高较矮但有1~2枝垂直生长较高,整体呈簇状分布,对阳光等可利用资源的吸收效果较差; 相反,针叶树种在这些样地内占优势地位,调查中发现样地a~c内的急尖长苞冷杉数量最多,因此,样地a~c的川滇高山栎以种间竞争为主。
总竞争指数是川滇高山栎的种内和种间竞争指数之和。在所有样地中,小径级对象木受到的竞争压力最大,总竞争指数随着径级增加而逐渐下降。由此可见,随着树龄增加和树体增大,竞争力持续提高,受到外界竞争的压力减小。样地a、b、c、d的总竞争指数分别为94.918、105.717、149.010、170.228,每株对象木的平均竞争指数分别为3.273、3.109、2.070、1.685。对不同海拔样地的分析发现,川滇高山栎对象木的平均竞争指数与海拔呈正相关,即平均竞争指数随海拔升高而增加。

表4 川滇高山栎天然林对象木胸径分布和竞争指数
3.3 川滇高山栎的伴生树种及竞争指数
不仅海拔和林木胸径影响川滇高山栎的竞争指数大小,不同种类的竞争木也对其产生影响。在4个伴生树种中,样地a、b、c内竞争力最大的都是急尖长苞冷杉,其竞争指数分别为64.334、49.539、38.761(表5),表现为随着海拔降低而减小。各树种在各样地的分布情况有所不同,所以竞争指数也存在较大差异。川滇高山栎伴生种的竞争指数在样地a表现为急尖长苞冷杉>林芝云杉>糙皮桦>白桦,在样地b表现为急尖长苞冷杉>白桦>林芝云杉>糙皮桦,在样地c表现为急尖长苞冷杉>白桦>糙皮桦>林芝云杉,在样地d表现为白桦>糙皮桦>急尖长苞冷杉>林芝云杉。调查中发现,川滇高山栎天然林内的林芝云杉数量相对较少,其次是糙皮桦。在川滇高山栎的整个生长发育过程中,周围林木不断与其争夺资源,但由于每种林木的生态学和生物学特征不同,表现出的竞争指数也不同。通常,随着林木年龄和胸径逐渐增大,冠幅显著扩大,一些阔叶树种的竞争力会明显提高,而针叶树种的竞争力逐渐减弱。
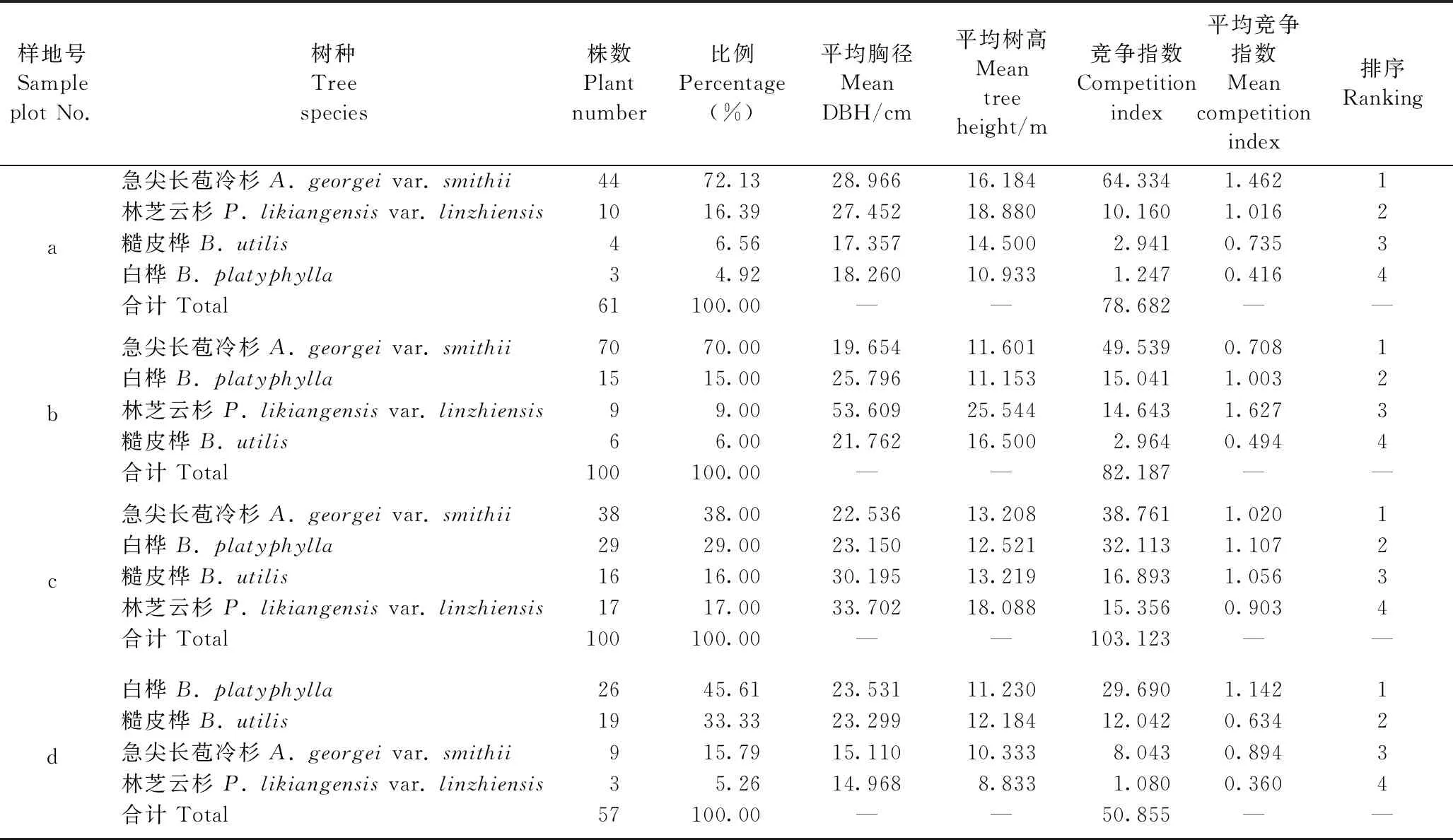
表5 各样地伴生树种种类组成及竞争指数
3.4 川滇高山栎竞争指数与胸径的关系及预测
林木竞争能力大小受多种因素影响,如冠幅、胸径、生态位宽度等,其中影响最大的是胸径(黄小波等, 2016)。以每株对象木的竞争指数为因变量,对象木个体胸径为自变量,采用对数函数、指数函数、幂函数、多项式、Logistic模型探究二者的关系,比较其决定系数(R2)大小,通过了显著性检验,结果表明各样地均达极显著水平(表6)。最优拟合函数在样地a为幂函数 CI=aDBH-b,在样地b和c为三次多项式函数CI=aDBH3+bDBH2+cDBH+d,在样地d为指数函数CI=aebDBH。将各样地调查数据和模型拟合情况展示在图2中,表明竞争指数随对象木胸径增加而逐渐下降,其中对象木胸径大于25cm时的下降幅度变缓。
利用表6中的回归模型预测了4个样地中川滇高山栎不同胸径大小对象木的个体竞争指数(表7),由表7可知,在4个样地中,竞争指数均在胸径为5cm时最大,并随胸径增加而逐渐减小,在胸径5~25cm内下降幅度较大,当胸径超过25cm后下降变缓并渐趋稳定。

表6 对象木胸径及其竞争指数拟合分析
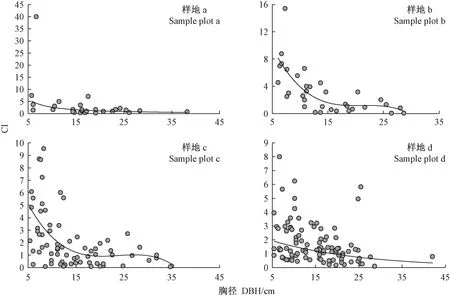
图2 川滇高山栎天然林对象木竞争强度与其胸径的回归曲线

表7 川滇高山栎天然林对象木个体竞争指数的模型预测结果
4 讨论
4.1 不同海拔的川滇高山栎种内种间竞争差异
竞争是植物种内种间的重要关系,它不仅体现植物生长发育状态,还可促进群落更新演替(惠刚盈等, 2013)。计算竞争指数通常用Hegyi单木竞争模型,但其局限性在于通过静态的胸径和距离来计算,未考虑气候变化、空间分布等动态因子,今后应进行动态分析。林木的竞争木范围大小与林窗大小、上层树木影响范围等有关,研究中以5 m(高浩杰等, 2017)、6 m(巢林等, 2017)、8 m(徐建等, 2014)、10 m(喻泓等, 2009)为竞争木范围,还有人利用Voronoi图确定竞争木范围(汤孟平等, 2007; 李际平等, 2015; 曹小玉等, 2016),相较而言很难找到普遍适应的竞争木范围,需根据具体情况来确定。
本研究通过比较藏东南川滇高山栎天然林内树木的种内与种间竞争的海拔差异,发现在样地a、b、c以种间竞争为主,说明他疏作用大于自疏作用,这可能由川滇高山栎在长期进化过程中的生态位特化造成的,反映了环境筛选和生物个体特点间的关系。在海拔较高(3 540~3 740 m)的样地内,川滇高山栎的林木数量少于其伴生树种的数量,并多呈簇状,对环境资源利用率较低,因此其竞争力较弱,以种间竞争占主导。这一现象与云南红豆杉(Taxusyunnanensis)(李帅锋等, 2013)、大别山五针松(Pinusdabeshanesis)(项小燕等, 2015)的种内种间竞争研究结果相似。在本研究中,林分内与川滇高山栎竞争较激烈的树种是属于暗针叶树种的急尖长苞冷杉(高郯等, 2020),它主要分布在海拔3 400~4 200 m的高山地带,是藏东南主要优势树种之一。样地d内的川滇高山栎林木以种内竞争为主,即自疏作用占主导,原因可能是相对低的海拔(3 440 m)更适合川滇高山栎生长,可以长成冠幅更大、树高更高的乔木,以此来占据更多环境资源和提高自身竞争力。实地调查中发现,样地d中的川滇高山栎林木数量骤然增加,成为优势树种,因此,以种内竞争为主,这与华南五针松(Pinuskwangtungensis)(王俊等, 2020)、兴安落叶松(张准霞等, 2019)的种内种间竞争研究结果类似。在影响林木竞争的因子中,物种的生态位越接近和生态习性越相似,它们的竞争就越激烈; 在生态系统中占据主要地位的树种,通常竞争力也较强(张靖梓等, 2016)。
4.2 川滇高山栎对象木在生长发育过程中所受压力的变化
本研究表明,随着对象木个体胸径增加,它对应的竞争指数也在不断下降,受到的竞争压力逐渐减弱,这与很多研究结果一致(罗梅等, 2018; 刘彤等, 2007; Zhang et al., 2010)。小径级的川滇高山栎对象木在争夺阳光、水分等资源时受到其他竞争木的激烈竞争,其抗逆性较差,所受竞争压力较大。对象木在生长过程中适者生存,随着一些林木被淘汰,树木间的距离会变大,最后生存下来的个体到达成熟阶段时,会成为主林层的大树,周围其他竞争木的影响会随之减小,因此受到的竞争压力就会减弱。本研究的模型预测发现,当川滇高山栎对象木胸径超过25cm时,竞争指数下降幅度趋缓,竞争木对其产生的影响逐渐减弱并最后趋于稳定。
4.3 对川滇高山栎种群的保护措施
藏东南地区属湿润半湿润气候,水分充足,川滇高山栎叶片的总厚度、海绵组织和栅栏组织的厚度等随海拔升高而逐渐增厚,这样不仅可增加植物抗寒性能,且具有良好保温效果(郭文文等, 2021),因此,川滇高山栎成为较高海拔森林的适生树种。但野外调查中发现: 该树种在海拔3 540~3 740 m的样地内幼苗较少,且分蘖数较多,影响树木生长高度,受到伴生种的剧烈争夺,种群更新困难。针对这些问题,可以有限地进行人工干扰,即在幼苗较少的天然林内进行一定程度的人工去杂,砍伐一些竞争力较强的非濒危树种,拓宽幼苗生长空间,并结合进行人工抚育,以更好保存较高海拔区域的川滇高山栎种质资源。
5 结论
基于Hegyi单木竞争模型,调查分析藏东南川滇高山栎天然林内对象木的种内与种间竞争指数。该指数存在海拔差异,在海拔3 540~3 740 m范围内以种间竞争为主,在海拔3 440 m以种内竞争为主,即种间竞争指数随海拔降低而不断下降,川滇高山栎逐渐成为优势种。川滇高山栎林木的竞争指数表现出随个体胸径增加而不断下降的变化规律,在胸径大于25cm时下降幅度变缓并渐趋稳定,停留在较低水平。
猜你喜欢
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
林业科学研究(2021年6期)2022-01-05
内蒙古林业调查设计(2021年5期)2022-01-05
成都信息工程大学学报(2019年6期)2019-08-13
音乐天地(音乐创作版)(2018年4期)2018-07-18
宝藏(2018年3期)2018-06-29
蔬菜(2018年5期)2018-05-17
林业勘察设计(2017年4期)2017-07-06
Zoological Research(2016年1期)2016-03-22
杂草学报(2015年2期)2016-01-04
