
猪肉肌原纤维蛋白氧化对亚硝化的影响
2023-01-12 05:54顾如霞兰艳丽葛凤芹李锋王永丽
中国食品学报 2022年12期
顾如霞,兰艳丽,葛凤芹,李锋,王永丽
(山东农业大学食品科学与工程学院 山东省高校食品加工技术与质量控制重点实验室 山东泰安 271018)
亚硝胺由食品中的亚硝酸盐、硝酸盐与蛋白质降解产物氨基酸、胺类物质发生亚硝化反应形成。它具有致癌、致畸及致突变作用,过量摄入会威胁人体健康。食品中亚硝胺类化合物的主要形成途径为:蛋白质分解产生氨基酸,氨基酸经脱羧作用生成肿胺和芳香族叔胺等含氮物质;在酸性条件下亚硝酸盐可生成亚硝酸,并进一步分解产生活性亚硝化试剂。两者在弱酸性环境下,发生亚硝化反应形成各种亚硝胺类化合物。亚硝酸盐和胺类化合物是反应必不可少的前体物质,而酸性pH 值是必须具备的反应条件[1]。此外,亚硝胺的形成还受其它诸多因素的影响,如肉基质环境的氧化-还原电位,前体物质的浓度、温度,亚硝化反应的促进剂和抑制剂,蛋白质的含量,蛋白质氧化程度及氧化形成的产物等[2]。
蛋白质暴露在活性氧环境下会发生氧化,导致肽键断裂、侧链氨基酸修饰、蛋白质分子间共价交联等变化,并最终影响蛋白质分子性质和生物学功能。最新研究表明,蛋白质氧化与亚硝胺形成关系密切,不仅可直接为亚硝胺形成提供前体物,还可间接影响食品中亚硝化反应过程[3-4]。蛋白质氧化产生多种含氮衍生物,过氧化氢诱导蛋白氧化产生二级氧化产物,如丙二醛或4-羟基壬烯醛,可作为亚硝胺的前体物质[5]。蛋白质在加工或储藏过程中发生的氧化、亚硝化和分解都会导致肉制品营养价值的降低。蛋白质氧化可能影响亚硝化进程。目前关于肉与肉制品中蛋白质氧化对N-亚硝胺形成的影响鲜有报道。
本文以猪肉肌原纤维蛋白为研究对象,构建羟自由基氧化体系,研究不同氧化程度对肌原纤维蛋白侧链修饰、构象变化、蛋白交联聚集及亚硝化的影响。通过测定羰基、总巯基、二硫键、自由氨基、二聚酪氨酸和表面疏水性等指标,揭示蛋白质氧化程度对NaNO2、3-硝基酪氨酸(3-nitrotyrosine,3-NT)和亚硝胺形成的影响。通过相关性分析阐明蛋白质氧化与亚硝化之间的关系,研究结果将为了解蛋白氧化体系中亚硝胺形成机制,控制蛋白氧化,提高肉制品安全品质提供理论依据。
1 材料与方法
1.1 材料与试剂
新鲜猪背最长肌,购于泰安大润发超市双汇肉食专柜。
亚硝胺混标、1,1-二苯基-2-苦基苯肼(2,2-diphenyl-1-picrylhydrazyl,DPPH)、β-巯基乙醇、2,4-二硝基苯肼(2,4-dinitrophenylhydrazine,DNPH)、5,5’-二硫代双-2-硝基苯甲酸(5,5’-dithio-bis-(2-nitrobenzoic acid),DTNB)、4-氟-7-硝基-2,1,3-苯并氧杂恶二唑(7-fluoro-4-nitrobenzo-2-oxa-1,3-diazole,NBD-F)、牛血清蛋白,美国Sigma 公司;2-硝基-5-硫代磺基苯甲酸(2-nitro-5-thiosulfobenzoate,NTSB)、食品级亚硝酸钠(≥99%)、十二烷基磺酸钠、二氯甲烷、四甲基乙二胺、盐酸纳乙二胺、邻苯二甲醛、对氨基苯磺酸和水溶性维生素E,上海Solarbio 公司;柠檬酸钠、丙烯酰胺、过氧化氢、酒石酸钾钠等为分析纯试剂,上海国药集团。
1.2 仪器与设备
ME204/02 分析天平,美国Mettler-Toledo 公司;SpectraMax M2 多功能酶标仪,美国MD 公司;7890A/5975C 三重四级杆气质联用仪,美国Agilent 公司;Bio-Rad 电泳仪,美国Bio-Rad 公司;F-7000 荧光分光光度计,日本日立公司;CTHI-250B型恒温恒湿箱,STIK 仪器设备(上海)有限公司;XM-P22H 型无级调功超声波清洗机,小美超声仪器(昆山)有限公司;Allegra 64R 型高速冷冻离心机,美国Beckman Coulter 公司;IKAT18basic 型高速分散机,德国IKA 公司;DHG-9030A 型电热恒温鼓风干燥箱,上海一恒科技有限公司;Turbo Vap LV 全自动浓缩工作站,美国Caliper 公司。
1.3 试验方法
1.3.1 肌原纤维蛋白的提取 肌原纤维蛋白(MP)的提取参考Park 等[6]的方法并略作修改。新鲜猪肉剔除可见脂肪、筋膜等组织,放入绞肉机绞碎,称取20 g 肉样,加入4 倍体积的pH 7.0 磷酸盐缓冲液(10 mmol/L NaH2PO4,100 mmol/L NaCl,1 mmol/L EGTA,2 mmol/L MgCl2),高速匀浆(10 000 r/min,30 s),4 ℃离 心(4 000 r/min,10 min)后弃上清液,此过程重复3 次。用4 倍体积的氯化钠洗液(0.1 mol/L)匀浆(10 000 r/min,30 s),置4 ℃离心(4 000 r/min,10 min),弃上清,重复两次。第3 次洗液匀浆后,用4 层纱布将其过滤,最后用0.1 mol/L 盐酸溶液调pH 值为6.25,离心,去上清液,所得白色膏状物即肌原纤维蛋白。将制备好的MP 置离心管中4 ℃冷藏,48 h 内使用。采用双缩脲法测定蛋白的含量,以牛血清蛋白为标准蛋白制作标准曲线,y=0.0033x+0.129,R2=0.997。
1.3.2 肌原纤维蛋白氧化处理 用25 mmol/L 磷酸盐缓冲液(含0.6 mol/L NaCl,pH 6.25)将MP蛋白膏稀释到40 mg/mL。将稀释好的MP 溶液加入离心管中,再加入羟自由基氧化体系(0.01 mmol/L FeCl3,0.1 mmol/L 抗坏血酸,H2O2),最终体系H2O2浓度分别是0,10,30,50,70 mmol/L,蛋白的最终质量浓度为25 mg/mL,将氧化体系置于37 ℃氧化1 h。通过添加水溶性维生素E(Trolox,1 mmol/L,终浓度)来终止氧化反应。以未加氧化剂,含Trolox 的蛋白溶液为空白对照。
1.3.3 肌原纤维蛋白理化特性分析
1)羰基含量测定 参照Oliver 等[7]的方法,采用2,4-二硝基苯肼(DNPH)法测定,摩尔消光系数为22 000 mol/(L·cm),羰基含量表示为nmol/mg MP。溶剂对照组,开始时加入2 mL 缓冲液代替蛋白溶液,其余操作相同。
2)总巯基含量测定 参照Cao[8]等的方法,采用5,5’-二硫代双-2-硝基苯甲酸(DTNB)法于412 nm 处测定吸光值,采用摩尔消光系数13 600 mol/(L·cm)计算,总巯基含量表示为nmol/mg MP。
3)二硫键含量测定 参考Liu 等[9]的方法,向各蛋白氧化体系溶液中加入2-硝基-5-硫代磺基苯甲酸(NTSB)试剂,避光反应,震荡15 min 后,于412 nm 处测定其吸光值。
4)自由氨基含量测定 参照Adler-Nissen等[10]的方法,采用邻苯二甲醛法测定,蛋白氧化处理后的自由氨基含量通过L-赖氨酸作标准曲线确定,单位为nmol/mg MP。
5)二聚酪氨酸含量测定 参照Davies 等[11]的方法,用20 mmol/L 磷酸盐溶液(含0.6 mol/L KCl,pH 6.0)将氧化后的蛋白溶液稀释到1 mg/mL。蛋白溶液经离心(10 000 r/min,10 min)除去不溶性物质。用荧光分光光度计测定荧光强度,激发波长为325 nm,发射波长为420 nm,狭缝宽度为10 nm。二聚酪氨酸含量用所测荧光强度除以蛋白浓度获得,为相对荧光值,单位为(arbitrary units,简写A.U)。
1.3.4 肌原纤维蛋白构象变化分析 表面疏水性:参照曹云刚等[12]的方法,采用溴酚蓝(bromphenol blue,BPB)结合法。将肌原纤维蛋白用20 mmol/L 磷酸盐溶液(pH 6.25)溶解,稀释至5 mg/mL。吸取各处理组蛋白溶液1 mL 置于离心管中,加200 μL 溴酚蓝溶液(1 mg/mL)混合均匀,在6 000 r/min 条件下离心15 min,吸取上清液于595 nm 处测量吸光值。表面疏水性用蛋白暴露的疏水性氨基酸残基与溴酚蓝的结合量表示。
SDS-PAGE 凝胶电泳:参照Laemmli 等[13]研究方法,分离胶12%,浓缩胶4%,pH 8.3 电极缓冲液(0.05 mol/L Tris-HCl 溶液,0.384 mol/L 甘氨酸,0.1% SDS)。将电泳样品用样品溶解液(含10% SDS、50%甘油、加或不加10% β-巯基乙醇、0.1%溴酚蓝、1 mol/L Tris-HCl 缓冲液,pH6.8)配制蛋白终质量浓度为1 mg/mL,漩涡混合1 min,沸水浴5 min。样品上样量10 μL。电泳结束后取出胶片,用蛋白染色液染色1 h 后脱色,结合Tanon软件进行数据分析和处理。
1.3.5 肌原纤维蛋白亚硝化处理 肌原纤维蛋白亚硝化处理参考Yang 等[14]方法并稍作改动。将处理好的氧化蛋白各体系用25 mmol/L 磷酸盐缓冲液(含0.6 mmol/L NaCl、430 μmol/L 亚硝酸钠,pH 6.0)稀释,蛋白终质量浓度为20 mg/mL(所有浓度均为最终浓度)。亚硝化反应在37 ℃下孵育4 h,用NaOH 调整pH 值至13,终止反应。各反应体系置于4 ℃下,待测。
1.3.5.1 亚硝酸盐残留量测定 按照GB 5009.33-2016《食品中亚硝酸盐与硝酸盐的测定》方法测定。
1.3.5.2 3-硝基酪氨酸(3-NT)含量测定 参照Feng 等[15]的方法,取处理好的肌原纤维蛋白溶液100 μL,加入0.36 mL乙腈、0.04 mL 1 mol/L NBD-F、0.5 mL 0.1 mol/L 四硼酸钠溶液,混匀,60℃水浴2 min。冷却后过0.45 μm 滤膜,进液相色谱分析。色谱柱为250 mm×4.6 mm,5 μm;流动相为磷酸氢钾-磷酸缓冲液/乙腈(20 mmol/L,pH 3),流速1.0 mL/min,上样量10 μL。
1.3.5.3 N-亚硝胺含量的测定 取各处理组的肌原纤维蛋白溶液2 mL,用5 mL 二氯甲烷涡旋超声提取,10 000×g,4 ℃离心5 min,重复1 次。二氯甲烷层氮吹后用2 mL 二氯甲烷复溶,过0.25 μm微孔滤膜,进行GC-MS 分析。
气相色谱条件:色谱柱为DB-WAX(30 m×250 μm,0.25 μm)毛细管色谱柱,进样口温度250℃,不分流进样;采用程序升温:初始温度50 ℃,保持2 min,以8 ℃/min 升至150 ℃,保持5 min,然后以20 ℃/min 升至250 ℃,保持2 min;载气流速1 mL/min。串联质谱条件:扫描模式:选择离子扫描(SIM);离子源:EI 源;离子源和四极杆温度分别为230 ℃和250 ℃;电子倍增器电压1 301 eV;电离电压70 eV;灯丝电流:50 μA;扫描质量范围:m/z 30~150,溶剂延迟时间:2.5 min;碰撞气:99.99%高纯氩气,进样量1 μL。采用外标法定量计算亚硝胺,每次测定重复3 次。
1.4 统计分析
指标测定均重复3 次,试验数据以平均值±标准差表示。采用SAS 8.2 统计软件进行不同处理间单因素方差分析(one-way ANOVA analysis)和各指标间的Pearson 相关性分析。当P<0.05 时,表示均值间差异显著;P<0.01 时,表示均值间差异极显著。
2 结果与分析
2.1 氧化强度对肌原纤维蛋白理化特性的影响
蛋白羰基含量是表征蛋白质氧化的重要指标之一。由表1 所示,在羟自由基氧化体系中,羰基含量随H2O2浓度的上升显著增加(P<0.05)。氧化1 h 后,空白对照组羰基含量为7.27 nmol/mg,与0 mmol/L H2O2浓度条件下无显著差异(P>0.05),与曹云刚等[12]研究结果一致。当H2O2浓度为70 mmol/L 时,羰基含量为22.04 nmol/mg,比空白对照增加203.16%。羰基含量的升高可能是由于肌原纤维蛋白氨基酸侧链基团经自由基攻击直接氧化产生,或肽链断裂加速羰基化进程,赖氨酸、苏氨酸和脯氨酸易受到自由基攻击而产生羰基衍生物;还可通过和还原糖反应,以及结合非蛋白羰基化合物形成[16]。Nyaisaba 等[17]研究阿拉斯加鳕鱼MP 羰基含量随H2O2浓度增加而显著上升,羰基的形成对H2O2有浓度依赖性。Xia 等[18]研究猪肉肌原纤维蛋白氧化,发现H2O2浓度从0 到20 mmol/L,羰基含量显著增加79.75%,与本研究结果一致。

表1 氧化程度对MP 羰基、总巯基、二硫键和自由氨基含量的影响Table 1 Effect of oxidation degrees on carbonyl,total sulfhydryl,disulfide bond and free amino content of MP
蛋白富含巯基,在氧化条件下巯基极易被氧化成二硫键,因此总巯基和二硫键含量常被用来表征蛋白的氧化程度[19]。总巯基与二硫键含量见表1。随着H2O2浓度的升高,总巯基含量显著下降(P<0.05),二硫键含量显著增加(P<0.05)。空白对照组总巯基含量为102.32 nmol/mg,当H2O2浓度为0 mmol/L H2O2时,总巯基含量与空白对照组无显著差异(P>0.05)。氧化组(10,30,50,70 mmol/L H2O2)与空白对照样品相比,总巯基含量分别降低10.46%,17.95%,22.24%和25.21%,二硫键含量分别升高了240%,277%,426%和461%。Wang 等[20]研究铁催化氧化系统处理牦牛MP 时发现,巯基含量下降,二硫键含量升高,巯基含量的降低依赖于羟自由基的浓度,与本研究结果一致。氧化导致巯基含量下降,可能是由于MP 中半胱氨酸残基极易受到羟自由基攻击,巯基被氧化成二硫键。另外,MP 在氧化过程中变性展开,使巯基暴露,进而被氧化成二硫键。Soladoye 等[21]指出随着H2O2浓度增加,蛋白片段交联聚集,形成多肽分子间或分子内二硫键,进一步氧化成亚磺酸、磺酸或其它氧化产物,从而导致蛋白表面巯基含量减少,二硫键含量升高。
蛋白自由氨基含量如表1 所示。自由氨基含量随H2O2浓度的升高呈下降的趋势,与H2O2浓度呈显著负相关(P<0.05),这与总巯基含量的变化趋势一致。空白对照组和0 mmol/L H2O2处理组的自由氨基含量分别为104.72 nmol/mg 和105.51 nmol/mg,两者无显著性差异(P>0.05)。当H2O2浓度为70 mmol/L 时自由氨基含量为85.10 nmol/mg,与空白对照组相比降低了18.74%。这与曹云刚等[12]报道的猪肉MP 自由氨基下降趋势一致。自由氨基含量下降可能是由于蛋白质氨基酸侧链含有的-NH2基团在自由基的攻击下,通过脱氨基作用转化为羰基,羰基再与氨基反应形成席夫碱,从而使自由氨基含量降低[22]。
酪氨酸残基易被活性氧自由攻击发生氧化聚合反应,生成二聚酪氨酸,因此,可以通过测定二聚酪氨酸的含量来反映蛋白质氧化程度。自由基氧化体系对MP 二聚酪氨酸含量的影响如图1 所示。氧化1 h 后,不同的氧化浓度(10,30,0、70 mmol/L H2O2)条件下,相对于空白对照组,二聚酪氨酸含量分别增长到390.73,566.94,651.00,716.21 AU,增长率分别为37.00%,98.79%,128.27%和151.13%,0 mmol/L H2O2处理组与空白对照组无显著差异(P<0.05)。Wang 等[20]研究自由基对牦牛MP 氧化时发现,二聚酪氨酸含量随羟自由基氧化体系浓度的升高而增加,与本研究结果一致。高浓度羟自由基使蛋白氧化暴露出更多的氨基酸残基,蛋白氨基酸残基被氧化后,通过共价和非共价相互作用形成蛋白聚合物[23-24]。二聚酪氨酸可以由两条不同氨基酸多肽链或由同一氨基酸多肽链上的两个不同位置的酪氨酸自由基反应产生,导致蛋白质分子内或分子间的交联,蛋白质氨基酸残基的共价或非共价修饰可导致蛋白构象和功能性质发生显著改变[25]。

图1 氧化强度对MP 二聚酪氨酸含量的影响Fig.1 Effect of protein oxidation degree on dityrosine content of MP
2.2 氧化强度对肌原纤维蛋白构象稳定性的影响
表面疏水作用力是维持蛋白质构象和功能特性的一种重要作用力,溴酚蓝分子可与蛋白质分子表面的疏水性结合位点结合,如色氨酸和苯丙氨酸。在羟自由基氧化体系内,大量的自由基会不断攻击蛋白质表面的疏水性氨基酸残基,从而通过结合的溴酚蓝含量反映蛋白质表面疏水性的变化。如图2 所示,蛋白表面疏水性随H2O2浓度的增加先降低后增大。0 mmol/L H2O2浓度的MP 溴酚蓝的结合量为96.17 μg,与空白对照组无显著差异(P>0.05);30 mmol/L H2O2浓度的MP 溴酚蓝的结合量最低为86.77 μg,与空白对照组相比降低了11.68%。曹云刚等[12]研究不同氧化强度对猪肉MP 结构的影响时发现,当H2O2浓度为10 mmol/L 时,表面疏水性与空白对照组相比降低了11%,与本研究结果一致。这可能是由于氧化导致MP 结构展开后又通过疏水相互作用聚集,使表面疏水性下降。当H2O2浓度增到70 mmol/L 时,溴酚蓝的结合量升到111.08 μg,与空白对照组相比增长了13.07%(P<0.05)。说明强氧化导致蛋白结构展开,使蛋白内部的非极性基团暴露在分子表面,蛋白分子之间交联减少,从而导致蛋白质表面疏水性增强。Zhang 等[26]对翘嘴鲌MP 在羟基自由基氧化体系中的结构、功能及凝胶特性进行研究,发现随着H2O2浓度的增加,MP 表面疏水性增加。蛋白质氧化后导致的表面疏水性的差异可能与氧化剂浓度以及氧化剂与肌原纤维蛋白的比重有关[27]。
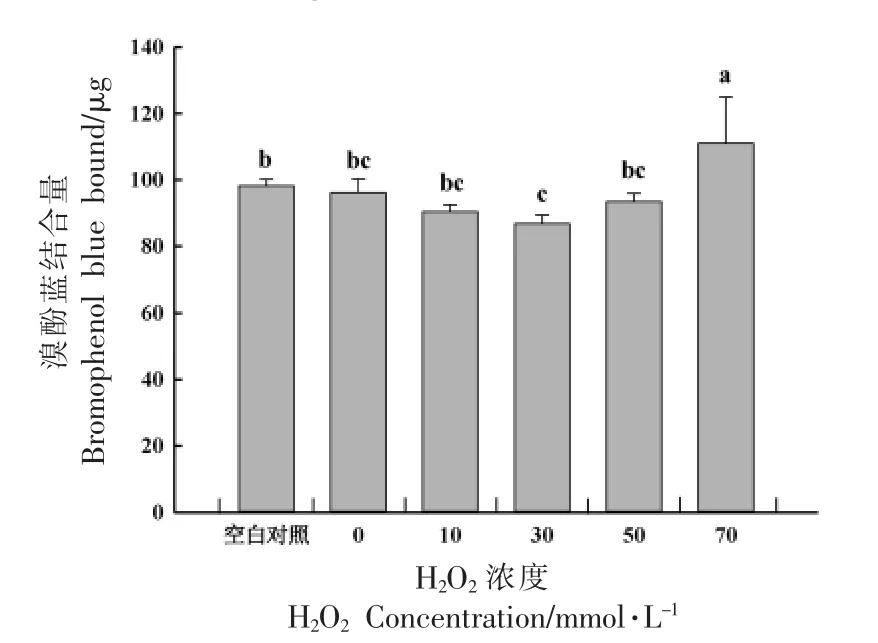
图2 氧化强度对MP 表面疏水性的影响Fig.2 Effect of protein oxidation degree on surface hydrophobicity of MP
氧化诱导蛋白质分子间或分子内共价交联和聚集,导致蛋白结构变化。通过非还原型(-β-ME)和还原型(+β-ME)SDS-PAGE 凝胶电泳反映氧化体系中肌原纤维蛋白结构的变化。如图3 所示,在非还原状态下,随着H2O2氧化浓度的增加,肌球蛋白重链(MHC)、肌动蛋白(Actin)和肌钙蛋白(Troponin)条带的强度明显降低,同时,在浓缩胶顶端大分子高聚物明显增多,且分子质量超过了200 ku,说明这些聚合物主要是来自肌球蛋白重链、肌动蛋白和肌钙蛋白的聚合。高分子聚合物的形成可能是由于氧化导致蛋白质分子间或分子内二硫键共价交联,以及蛋白质的交联和聚集引起的[6],与巯基含量降低和二硫键含量升高结果一致。在还原状态下,随着H2O2氧化浓度的增加,肌球蛋白重链和肌动蛋白条带强度明显加深,说明在氧化体系中蛋白交联和聚集的主要形式是二硫键,这与阿拉斯加鳕鱼在羟基自由基氧化条件下研究结果一致[17]。然而,仍有一部分聚合物在浓缩胶顶部,部分肌球蛋白重链没有恢复,说明聚合物中还存在除二硫键以外的其它共价交联,如:氧化导致二聚酪氨酸交联,羰基-氨基之间共价交联等诱导的聚集[12],这与二聚酪氨酸随H2O2浓度的增加,含量升高的结果一致。

图3 氧化强度对MP 影响的SDS-PAGE 图谱Fig.3 Effect of protein oxidation degree on SDS-PAGE patterns of MP
2.3 氧化强度对肌原纤维蛋白亚硝化的影响
氧化强度对MP 亚硝化的影响见表2。空白对照组NaNO2残留量为60.57 mg/kg,NaNO2残留量随着H2O2浓度的增加而显著降低(P<0.05),说明NaNO2在羟自由基氧化体系中,可作为前体物生成3-NT、NDEA 和NDMA,这与Vossen 等[3]研究NaNO2对MP 氧化的影响结果一致。NaNO2和羟基自由基反应能够产生活性氮,引发蛋白质的亚硝化和氧化反应[4,15],酪氨酸残基被活性氮修饰生成3-NT,因此其被认为是蛋白质亚硝化的标记物。MP 中3-NT 含量随H2O2氧化强度的增加而显著增加(P<0.05),当H2O2浓度增到70 mmol/L时,3-NT 含量升到7.04 nmol/mg,与空白对照组相比增长了43.38%(P<0.05)。结果表明,氧化强度增加促进了蛋白质亚硝化作用,这与Vossen 等[3]研究结果一致。空白对照组的NDMA 含量为0.93 μg/mL,70 mmol/L H2O2处理后NDMA 含量升到1.05 μg/mL,比空白对照组增加了12.9%。70 mmol/L H2O2处理组中NDEA 含量为1.04 μg/mL,比空白对照组显著增加了11.54%(P<0.05)。杨华等[28]研究反复冻融的猪肉蛋白对亚硝胺形成的影响,结果发现反复冻融造成蛋白质氧化,促进了二乙基亚硝胺(NDEA)的形成。

表2 氧化程度对MP 亚硝化的影响Table 2 Effect of oxidation degrees on protein nitrosation of MP
肌肉中易发生亚硝化的游离氨基酸包括脯氨酸、甘氨酸、丙氨酸和缬氨酸,以及具有生物活性的腐胺和尸胺,被认为是亚硝胺的前体物质[5]。在氧化体系中,抗坏血酸和亚硝酸钠反应可以产生一氧化氮,一氧化氮进一步与超氧化物自由基反应形成过氧硝酸盐,过氧硝酸盐可以启动蛋白质的亚硝化作用,产生3-NT[15]。在氧化体系中加入硝酸盐或亚硝酸盐,羰基的形成和巯基的氧化以及可能的脱氨作用都能导致新鲜猪肉中产生仲胺化合物,仲胺化合物与亚硝酸盐反应生成亚硝胺[29-30]。蛋白质氧化过程复杂,能够产生多种含氮衍生物,蛋白质降解产生的仲胺能促进亚硝胺的生成。本研究也证实蛋白质氧化产物作为亚硝化反应的前体物质,促进了N-亚硝胺的形成。
2.4 肌原纤维蛋白质氧化指标与亚硝化的相关性分析
肌原纤维蛋白各氧化指标与亚硝化之间的相关性见表3。H2O2浓度与MP 中羰基(r=0.905)、二硫键(r=0.908)和二聚酪氨酸(r=0.956)呈极显著正相关(P<0.01),H2O2浓度与总巯基(r=-0.934)和自由氨基(r=-0.936)呈极显著负相关(P<0.01)。说明随着氧化强度的增加,提高了MP 表面疏水性和二聚酪氨酸含量,降低了自由氨基和总巯基含量,蛋白结构受到破坏。NaNO2与MP 中羰基(r=-0.844)、二硫键(r=-0.879)和二聚酪氨酸(r=-0.888)呈极显著负相关(P<0.01),与总巯基(r=0.849)和自由氨基(r=0.924)呈极显著正相关(P<0.01),说明NaNO2能抑制蛋白氧化,避免巯基和氨基受羟自由基攻击。Feng 等[15]研究不同亚硝酸盐添加量对火腿肌原纤维蛋白氧化的影响,结果高剂量的NaNO2能显著抑制蛋白羰基和二聚酪氨酸的形成。

表3 肌原纤维蛋白质氧化指标与亚硝化之间的相关性Table 3 Correlation coefficients between protein oxidation variables and protein nitrosation of MP
3-NT 与蛋白质氧化各指标呈显著相关性。3-NT 与MP 中羰基(r=0.860)、二硫键(r=0.835)和二聚酪氨酸(r=0.792)呈极显著正相关(P<0.01),与总巯基(r=-0.824)、自由氨基(r=-0.865)和NaNO2(r=-0.778)呈显著负相关(P<0.05)。3-NT 与NDMA(r=0.788)和NDEA(r=0.812)呈显著正相关(P<0.05),这与Feng 等[1]研究亚硝酸盐对香肠中蛋白质氧化的影响结果一致。说明3-NT 与蛋白氧化程度及亚硝胺含量呈正相关,可能是蛋白质氧化和亚硝化的一个标志性产物[3]。蛋白质氧化和添加NaNO2促进了MP 亚硝化进程。
NDMA 含量与MP 氧化指标中羰基(r=0.779)、二硫键(r=0.757)和二聚酪氨酸(r=0.762)呈显著正相关(P<0.05),与总巯基(r=-0.817),自由氨基(r=-0.769)和NaNO2(r=-0.654)呈显著负相关(P<0.05)。NDEA 含量与MP 氧化指标中羰基(r=0.797)、二硫键(r=0.808)和二聚酪氨酸(r=0.829)呈显著正相关(P<0.05),与总巯基(r=-0.864),自由氨基(r=-0.857)和NaNO2(r=-0.760)呈显著负相关(P<0.05)。说明肌原纤维蛋白氧化促进NDMA 和NDEA 的形成。Sun 等[30]研究猪肉蛋白氧化与二乙基亚硝胺(NDEA)生成量的相关性时发现,MP 氧化导致羰基化合物生成,自由氨基降低,MP 蛋白氧化促进NDEA 的形成。Yang 等[14]研究发现NDEA 随蛋白质氧化程度的增加而增加,并与蛋白质羰基的产生和巯基的减少显著相关(P<0.05)。
3 结论
在羟自由基氧化体系中,随着氧化强度的增加,肌原纤维蛋白羰基、二硫键、疏水性和二聚酪氨酸含量上升,总巯基和自由氨基含量下降。同时研究了氧化强度对蛋白亚硝化的影响,发现随着H2O2浓度的增加,NaNO2残留量降低,3-NT、NDEA 和NDMA 含量升高,3-NT 与亚硝胺含量呈显著正相关,说明3-NT 是蛋白亚硝化的标志物,氧化促进蛋白的亚硝化进程,导致NDMA 和NDEA 的形成。通过蛋白氧化指标与亚硝化之间的相关性分析,揭示蛋白质氧化产物可作为亚硝化反应的前体物质促进N-亚硝胺的形成。研究结果将为阐明肉制品在氧化条件下亚硝胺的形成机制及建立抑制蛋白氧化和亚硝化进程相应的控制技术提供理论依据。
猜你喜欢
石油化工腐蚀与防护(2022年1期)2023-01-06
食品科学技术学报(2022年5期)2022-10-11
食品工业科技(2022年17期)2022-08-27
化工设计通讯(2020年11期)2020-11-17
食品与发酵工业(2020年16期)2020-09-03
食品科学(2020年11期)2020-07-13
分析化学(2019年4期)2019-05-13
中成药(2017年10期)2017-11-16
家庭百事通·健康一点通(2016年12期)2016-12-29
食品工业科技(2014年23期)2014-03-11
