
细叶百合DREB转录因子生物信息学及胁迫应答表达分析
2023-01-12 02:37谭萌萌孙绍营王静文王依萍张彦妮
西北林学院学报 2023年1期
谭萌萌,孙绍营,王静文,王依萍,燕 浩,张彦妮
(东北林业大学 园林学院,黑龙江 哈尔滨,150040)
植物的生长发育及进化过程常常会受到干旱、低温、高盐等非生物胁迫的影响。DREB转录因子是植物中同非生物胁迫相关的重要转录因子之一,含有1个由60~70氨基酸残基组成的保守结构域,位于第14位的V(缬氨酸)和第19位的E(谷氨酸)对转录因子与顺势元件的结合至关重要[1-2]。DREB家族成员可以分为A1~A6亚族,A1成员主要参与冷胁迫反应,A2成员主要参与干旱和高盐应激调节,A1~A2成员功能已在水稻(Oryzasativa)[3]、高粱(Sorghumbicolor)[4]、虎杖(Polygonumcuspidatum)[5]等研究中得到证实。A3~A6成员的功能与植物的生长发育和非生物胁迫都有一定关系,拟南芥(Arabidopsisthaliana)中属于A5的DEAR4基因与叶片的衰老相关,可由ABA和干旱等诱导表达[6];拟南芥中A6成员RAP2.4可以在干旱胁迫下激活叶片中表皮蜡的生物合成[7],过表达菊花(Dendranthemamorifolium)A6亚族基因CmDREB6可提高菊花的耐热性[8]。目前,已在草莓(Fragariaananassa)[9]、小麦(Triticumaestivum)[10]、油菜(Brassicanapus)[11]、水稻[12]、马铃薯(Solanumtuberosum)[13]等多种植物中鉴定出DREB家族成员,但对细叶百合DREB转录因子研究甚少。
细叶百合(Liliumpumilum)为百合科(Liliaceae)百合属(Lilium)植物,花下垂,花被鲜红,观赏价值极高,具有较强的抗旱和抗病性,为百合抗性育种的重要亲本[14]。目前,细叶百合的研究主要集中在杂交育种、组织培养等方面,关于细叶百合分子机制的研究初有进展。近年来虽从细叶百合中鉴定出一些与非生物胁迫相关的转录因子WRKY[15]和MADS-box[16]等,但有关细叶百合DREB转录因子的研究鲜有报道。因此,筛选鉴定细叶百合DREB转录因子并分析细叶百合DREB基因在逆境下的表达模式,有助于对其进一步开发利用。本研究从细叶百合转录组(根茎叶)中筛选出12个DREB转录因子,对其进行生物信息学分析,并用qRT-PCR技术探究了DREB转录因子在干旱、低温、ABA和高盐胁迫下的组织表达模式,为筛选优质抗逆基因奠定理论基础。
1 材料与方法
1.1 材料
以细叶百合组培苗为研究材料,苗龄45 d,高度10 cm,具8~10个叶片,鳞茎长2 cm,根部健壮,整体生长状态良好。DREB转录因子源自课题组已构建的细叶百合(根茎叶)转录组数据库(未发表)。
1.2 方法
1.2.1 DREB转录因子的鉴定分析 以“DREB”为关键词在细叶百合转录组“gene-expression.xlsx”文件中搜索获得DREB基因 ID。通过基因ID在“Trinity-gene.fasta”文件中查找DREB基因序列。利用ORF finder确定DREB基因的ORF,并用CDD预测DREB基因的保守结构域,选择有完整的ORF和保守结构域的DREB作为候选基因。
1.2.2 DREB蛋白的生物信息学分析 使用在线分析软件Protparam、ProtScal、Cell-PLoc 2.0、SOPMA[17]、MEME对细叶百合DREB蛋白进行理化性质、亲/疏水性、亚细胞定位、二级结构及保守基序分析。运用MEGA 7.0软件对细叶百合和拟南芥的DREB蛋白进行系统进化分析。
1.2.3 非生物胁迫下DREB基因的表达模式分析 将细叶百合幼苗置于150 μmol·L-1ABA溶液、20% PEG6000溶液、200 mmol·L-1NaCl溶液和4 ℃光照培养箱中进行胁迫处理。各处理于0、2、6、12 h时分别取细叶百合的根尖、鳞茎、叶片,液氮速冻并保存于 -80 ℃冰箱中。每个处理进行3次生物学重复,每次重复3棵植株。
用OMEGA公司的提取试剂盒提取RNA,TOYOBO公司反转录试剂盒将RNA反转录成cDNA。以百合的LilyActin作为内参基因,引物序列见表1。qRT-PCR反应利用Roche Light Cycer96系统和UltraSYBR Mixture,反应体系为:2×UltraSYBR Mixture 10 μL,引物0.4 μL,cDNA模板为0.8 μL,用双蒸馏水调至20 μL。反应程序为:95 ℃ 6 min,95 ℃ 10 s,60 ℃ 30 s,72 ℃ 35 s,45个循环。采用2-ΔΔCt方法进行基因相对表达量的计算,Heatmapper在线软件绘制热图。
2 结果与分析
2.1 DREB转录因子的鉴定及分析
经初步筛选,从细叶百合转录组数据库中得到51个细叶百合DREB转录因子,通过剔除序列重复、无完整ORF和保守结构域残缺的转录因子,最终筛选出12个DREB转录因子,命名为LpDREB1-LpDREB12(表1)。通过对细叶百合DREB蛋白序列的理化性质分析发现(表2):细叶百合DREB蛋白的序列长度在363~648 bp,理论等电点在4.55~10.96,其中LpDREB2和LpDREB3为碱性蛋白质;不稳定系数在35.85~90.51,其中LpDREB5为稳定性蛋白,其余为不稳定性蛋白;脂肪系数在55.12~75.7,蛋白质的亲水指数在-0.116~-0.803,亲水性最强的为LpDREB3蛋白。亚细胞定位预测分析表明(表2),7个基因定位在细胞核内,5个基因定位于细胞核和细胞质内,说明这12个DREB转录因子在细胞核内行使一定的功能。
SOPMA对细叶百合DREB蛋白的二级结构预测表明(表3):12个细叶百合DREB蛋白的结构主要由α螺旋和无规则卷曲构成,延伸链和β转角占比较少。所有蛋白中,LpDREB2蛋白的无规则卷曲占比最大,为64.16%;LpDREB8的α螺旋占比最大,为44.87%;除LpDREB2外,其他蛋白的α螺旋和无规则卷曲占比都大于25%,表明α螺旋和无规则卷曲对蛋白质三级结构的形成有一定影响。
2.2 DREB转录因子系统进化分析
为揭示细叶百合DREB转录因子家族的进化关系,以61个拟南芥DREB转录因子家族成员作为参考,与12个细叶百合LpDREB转录因子进行系统的进化分析。由图1可见,细叶百合DREB转录因子和拟南芥DREB转录因子聚类到一起,说明细叶百合和拟南芥的DREB转录因子具有较高的保守性。拟南芥AT1G75490转录因子属于DREBA-2亚组的成员,A-2亚组的成员都参与干旱响应,说明细叶百合LpDREB5、LpDREB9、LpDREB8和LpDREB7基因可能都与干旱胁迫相关。其余细叶百合DREB转录因子的功能也可与拟南芥等模式植物中功能明确的DREB转录因子进行预测。
2.3 DREB蛋白保守基序分析
通过对细叶百合LpDREB蛋白保守基序和保守结构域分析得出(图2),12个细叶百合DREB转录因子均有一个完整的AP2保守结构域,所含保守基序有一定差异。其中所有基因均识别到motif 1和motif 3,表明这2个基序高度保守。LpDREB4和LpDREB11蛋白含有相同的motif,LpDREB1和LpDREB12含有相同的motif,说明两者氨基酸序列结构最为相似。细叶百合DREB转录因子的进化关系和各自的基序表明,同一进化分支的LpDREB蛋白保守基序种类和位置大致相同,而不同进化分支的蛋白保守基序差异较大,由此推断同一进化分支的LpDREB转录因子的功能相似。

表1 qRT-PCR 引物序列
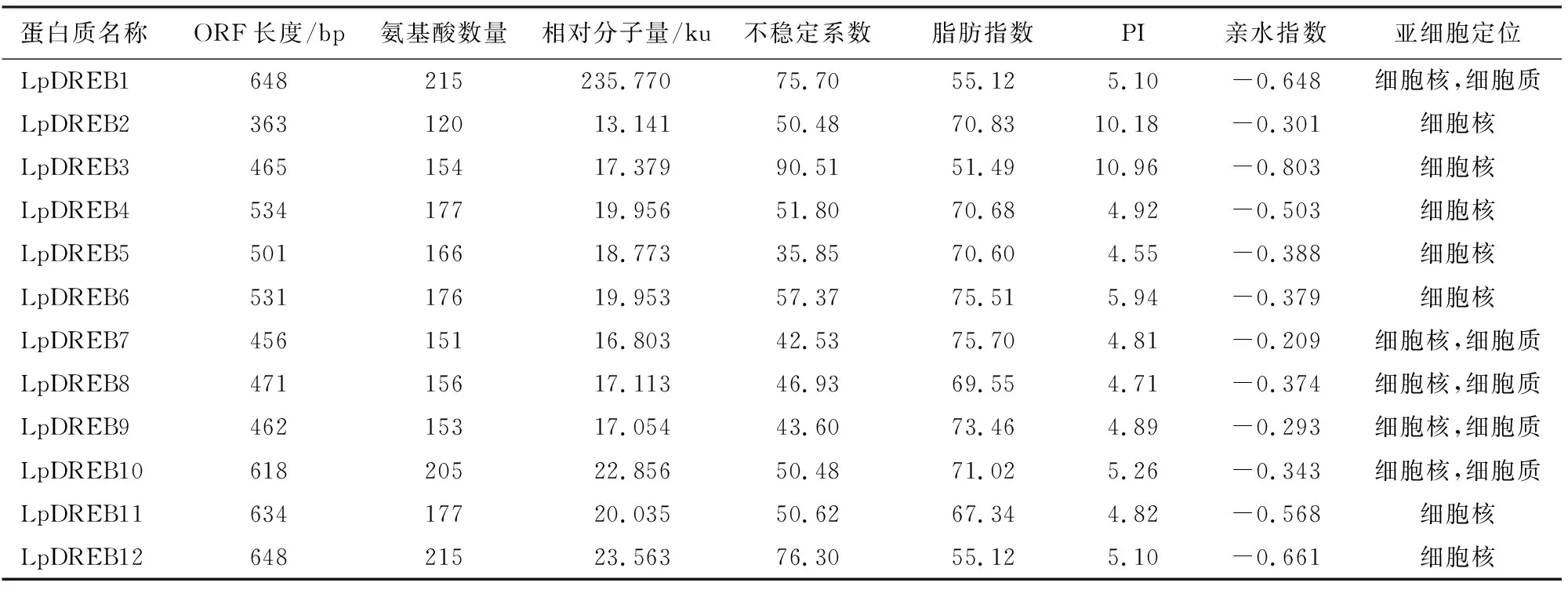
表2 细叶百合DREB转录因子的理化性质

图1 LpDREB转录因子和AtDREB转录因子进化分析

图2 LpDREB基因家族保守基序和保守结构域分析
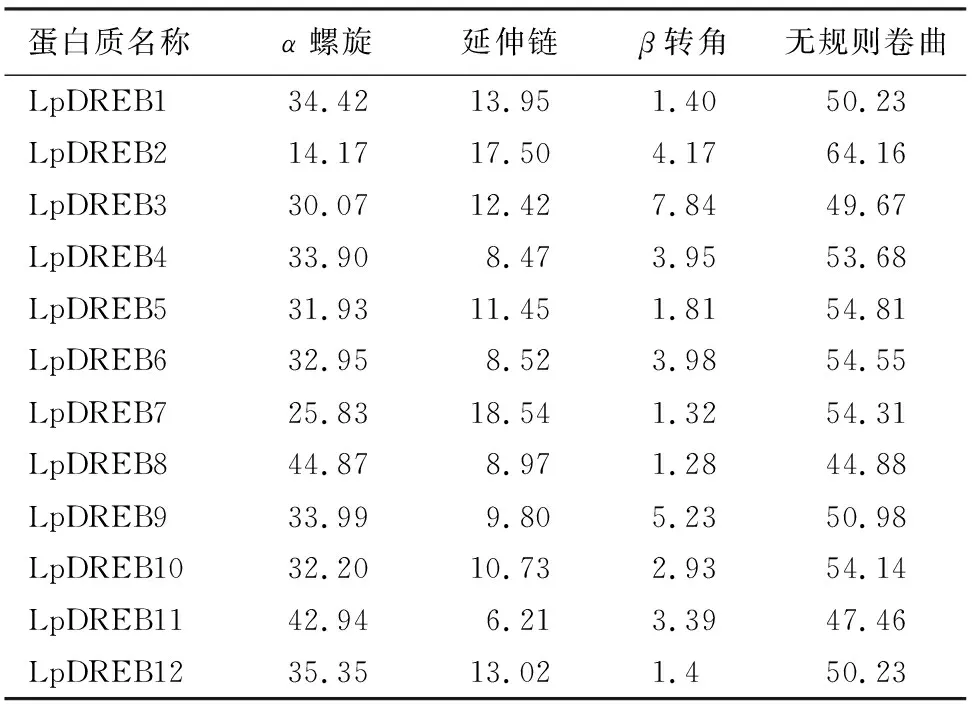
表3 细叶百合DREB蛋白二级结构
2.4 非生物胁迫下DREB基因的表达分析
2.4.1 干旱胁迫下DREB基因的表达模式 细叶百合在20% PEG6000模拟干旱胁迫下(图3A),根中6个LpDREB基因,胁迫 12 h后表达量上调;LpDREB1基因在所有胁迫时间下表达量均低于正常水平;LpDREB12和LpDREB6基因表达量为先上升后下降。鳞茎中,3个LpDREB基因在12 h胁迫后表达量上调,2个基因在所有时间点表达量均低于CK;LpDREB12、LpDREB2和LpDREB9基因在胁迫2 h表达量达到最高水平后随时间延长表达量降低至正常水平之下。叶中,5个LpDREB基因在胁迫12 h后表达量上调;LpDREB8和LpDREB9基因表达量随时间的延长表现为波动变化,且波动趋势相似。LpDREB1基因在各部位各时间处理下表达量均下调,推测其与干旱胁迫呈负相关;LpDREB10基因在根茎中表达量无明显变化,仅在叶中表达量上调,表明LpDREB10基因在不同组织中表达的特异性。
2.4.2 低温胁迫下DREB基因的模式分析 细叶百合在4 ℃低温处理下(图3B),根中除LpDREB3、LpDREB2和LpDREB1基因在2 h或12 h胁迫下基因表达量略高于正常水平,其余基因均无明显变化。鳞茎中, 4个LpDREB基因表达量无明显变化,7个LpDREB基因均在12 h胁迫后表达量高于正常水平,LpDREB12基因表达量先下降后逐渐升至上调水平。叶中,LpDREB6基因随胁迫时间增长,表达量逐渐升高,且都高于CK;LpDREB9和LpDREB12基因表达量先升高后降低;除LpDREB9和LpDREB12基因外,共10个基因在2 h或12 h 处理下上调表达。低温处理下,LpDREB2和LpDREB3基因在所有部位都上调表达,推测LpDREB2和LpDREB3与低温胁迫正相关。
2.4.3 盐胁迫下DREB基因的模式分析 细叶百合在200 mmol·L-1NaCl胁迫下(图3C),根中LpDREB10、LpDREB4、LpDREB12、LpDREB6和LpDREB2基因表达量无明显变化,其他基因在2 h或12 h时上调表达。茎中,LpDREB6和LpDREB12基因的表达水平呈现先升高后降低再升高的趋势;除LpDREB10、LpDREB4、LpDREB7和LpDREB5基因外,所有基因在12 h处理下表达量均高于CK。叶中,LpDREB1基因表达量始终低于CK;LpDREB3、LpDREB12和LpDREB2基因表达量无明显变化;LpDREB6基因在6 h时表达量上调,其余7个基因12 h胁迫后上调表达。盐胁迫下,LpDREB9和LpDREB11基因在根茎叶中均上调表达,推断LpDREB9和LpDREB11基因与植物耐盐性正相关。

图3 细叶百合LpDREB基因表达水平热图
2.4.4 ABA胁迫下DREB基因的模式分析 细叶百合在150 μmol·L-1ABA胁迫下(图3D),根中LpDREB3基因在各时间点表达量都低于CK;LpDREB11基因表达量随着胁迫时间延长,表达量逐渐增加,6 h时增至上调水平;LpDREB4和LpDREB5基因胁迫 12 h时表达量升至上调水平。茎中,除LpDREB5基因表达量无明显变化,其余基因在6 h胁迫后表达量均有不同程度提高,且4个基因在12 h胁迫后表达量持续高于正常水平。叶中,4个基因表达量无明显变化,8个基因分别在2、6 h或12 h时,表达量有不同程度上调。脱落酸胁迫下,LpDREB4和LpDREB11基因在各部位不同胁迫时间点下均上调表达,表明这2个基因与脱落酸胁迫调节机制正相关。
3 结论与讨论
从细叶百合转录组数据中最终得到12个细叶百合LpDREB转录因子,亚细胞定位预测显示,7个LpDREB转录因子定位在细胞核中,5个LpDREB转录因子定位在细胞核和细胞质中。转录因子主要在转录水平发挥作用,所以一般多定位在细胞核内。有研究表明,一些转录因子可分布在细胞质中作为结构蛋白参与细胞质的重要反应[18],针对这一现象可以开展关于细叶百合DREB转录因子的相关研究。蛋白的二级结构预测表明,α螺旋和无规则卷曲是细叶百合DREB蛋白二级机构的主要组成部分。系统进化分析表明 ,12个细叶百合DREB转录因子与拟南芥DREB家族聚类在一起,表明2个家族成员具有较高的同源性。通过对细叶百合DREB转录因子的研究,丰富了细叶百合基因家族的信息,为进一步研究细叶百合DREB转录因子奠定了理论基础。
为探求细叶百合DREB转录因子在非生物胁迫下的表达模式,对细叶百合进行了干旱、低温、高盐和ABA胁迫处理。干旱胁迫下,LpDREB1基因在根茎叶中表达量都低于正常水平,推测其与干旱胁迫负相关;11个LpDREB基因在根茎叶中被不同程度的诱导表达,且具有一定时序性。前人研究发现,干旱频发环境下,DREBs赋予作物多种非生物胁迫耐受性,并且DREBs可能具有提高作物水分利用效率的潜力[19]。因此,可以推测细叶百合DREB转录因子具有调控干旱胁迫的潜力。低温处理下,LpDREB2基因在细叶百合根茎叶中均有不同程度上调,其中叶中表达量最高,根中表达量最低。研究结果表明,细叶百合相同基因在不同部位对于胁迫的应答模式存在不同。盐胁迫下,LpDREB12和LpDREB10基因在茎或叶中上调表达,在其他组织中无明显变化。DREB转录因子的组织表达特异性已在大豆(Glycinemax)[20]、甘蔗(Saccharumspontaneum)[21]和菠萝(Ananascomosus)[22]、侧金盏花(Adonisamurensis)[23]等多种植物中报道,细叶百合LpDREB转录因子在根、茎、叶组织中的差异表达,暗示细叶百合在生长发育中功能的多样性以及应对逆境的差异性。细叶百合在干旱、盐和低温胁迫下,部分LpDREB基因随胁迫时间的变化表达量呈波动状态,这与苦荞麦(Fagopyrumtataricum)中FtDREB1和FtDREB2[24]的变化趋势相似。可能是非生物胁迫下,细叶百合LpDREB基因需要经过一个启动期才能被诱导表达,而一个功能基因又受到多个转录因子的调控,不同基因之间相互制约,表达量交替变化的结果。细叶百合LpDREB在应对非生物胁迫时基因表达量均有显著变化,说明LpDREB基因在应对逆境胁迫时发挥重要作用。
近年来的研究表明,植物对逆境胁迫的响应包括ABA依赖途径和非ABA依赖途径[25]。在拟南芥中过表达OsDREB1F,干旱胁迫能激活ABA依赖的胁迫响应基因RD29B和RAB18的表达,同时ABA也可以直接诱导OsDREB1F的表达,说明OsDREB1F通过2种途径参与对干旱胁迫的调控[26]。麝香百合(L.longiflorum)LlDREB1G在低温下被诱导表达,通过对转LlDREB1G拟南芥植株的功能研究 ,推测其可能以ABA依赖途径和非依赖途径参与非生物胁迫表达[27]。荧光定量结果显示,LpDREB4和LpDREB5基因能够被ABA和干旱胁迫诱导表达且变化趋势相同,推测LpDREB4和LpDREB5可能通过2种途径参与对干旱胁迫的应答。干旱胁迫会引起植物体内ABA的积累,从而使气孔开度和光合速率降低,进而阻碍植物的生长发育。因此,后续可继续开展试验阐明ABA与细叶百合干旱适应性的关系,为合理应用外源激素提供理论依据。植物应对各种逆境胁迫诱导时,DREB转录因子参与植物应答胁迫反应的重要过程[28]。因此,研究细叶百合DREB转录因子,对获得抗性新品种具有重要意义。
本研究对筛选鉴定出的12个细叶百合LpDREB转录因子进行了生物信息学和组织表达特异性分析,推测出部分基因的功能性。然而,仅凭表达分析模式判定个别基因的功能并不完全可信,影响其功能表达的因素较多。因此,今后可通过克隆细叶百合LpDREB基因并在细叶百合中过表达或抑制表达验证其功能性。
猜你喜欢
南方医科大学学报(2022年3期)2022-04-13
浙江大学学报(农业与生命科学版)(2021年3期)2021-07-10
中国预防兽医学报(2020年8期)2020-11-05
三农资讯半月报(2020年15期)2020-08-25
国际放射医学核医学杂志(2020年4期)2020-07-27
东坡赤壁诗词(2020年3期)2020-07-04
学生导报·东方少年(2018年4期)2018-05-14
上海农业学报(2017年3期)2017-04-10
广西林业科学(2016年3期)2016-03-16
天然产物研究与开发(2014年6期)2014-04-27
