
光质对蔬菜作物类胡萝卜素合成与调控研究进展
2023-01-31 03:13潘师峰徐志刚许亚良李清明郑胤建
中国蔬菜 2023年1期
潘师峰 徐志刚 许亚良 李清明* 郑胤建*
(1 中国农业科学院都市农业研究所,国家成都农业科技中心,四川成都 610000;2 南京农业大学农学院,江苏南京 210095)
类胡萝卜素(carotenoids)是具有广泛共轭多烯链的一大类萜类化合物,该共轭体系使得类胡萝卜素在可见光谱的蓝绿光(450~550 nm)波段范围内具有优异的光吸收性能(Moise et al.,2014)。在植物绿叶组织中,类胡萝卜素的组成和相对丰度对提升光合性能具有重要的作用,其中主要的类胡萝卜素有叶黄素(lutein)、β-胡萝卜素(β-carotene)、紫黄素(violaxanthin)和新黄素(neoflavin)等,并且它们在植物中的含量依次降低(DellaPenna &Pogson,2006)。同时,类胡萝卜素可以保护植物免受强光带来的损伤(Leonelli et al.,2017)。因此,植物体内的类胡萝卜素含量与植株生长发育、产量品质密切相关。通过不同的作用机制,类胡萝卜素或富含这些色素的食物对人体健康有益,大约有50 种不同的类胡萝卜素可转化为VA,并具有保护细胞和组织免受自由基和单线态氧破坏的抗氧化活性,可增强人体免疫功能,防止晒伤及延缓某些癌症的发生(Xavier &Pérez-Gálvez,2016)。
随着人造光源与设施栽培技术的发展,植物工厂成为现代农业发展的主流。LED(light-emitting diode)具有低能耗、简单轻便及可控性强等优点,被大规模投入到植物工厂中使用。近些年来,基于LED 的新型光源已经在植物栽培生产中得到了广泛的应用,研究发现使用不同波长组合的光源不仅提高了植物的光合作用效率,也影响了植物光形态建成(Bantis et al.,2018)。越来越多的研究表明,利用光调控植株生长发育及类胡萝卜素等次生代谢物积累以维持采后产品器官营养物质的稳定以及提升植株产品器官产量、品质是一种可行的方法。据报道,补充430 nm 和465 nm 波段蓝光可以显著提升普通白菜和芥蓝鲜质量以及类胡萝卜素、VC 和总酚的含量(Li et al.,2020)。红光和蓝光处理均显著提高了采后绿熟期樱桃番茄中番茄红素、β-胡萝卜素和可溶性糖含量,并且可以加速颜色的转变(Ngcobo et al.,2020)。蓝色LED 光会显著提高绿熟期辣椒总酚含量及抗氧化活性,红色LED光则显著提高了绿熟期辣椒果实中类胡萝卜素含量(Pola et al.,2020)。在标准红光/远红光和蓝光下补充绿光、黄光及橙光会引起芥菜苗类胡萝卜素含量显著上升,补充黄光显著提高了乌塌菜类胡萝卜素含量(Brazaitytė et al.,2015)。白光下补充红光、蓝光会显著提升叶用莴苣(生菜)鲜质量、叶绿素和类胡萝卜素含量,并且使植株株型更加紧凑强壮(Chen et al.,2016)。利用组合光谱UV-A/蓝光或红光/蓝光对叶用莴苣进行采前夜间补光,显著提升了类胡萝卜素、花色苷和酚类化合物等营养成分含量,并且可以使植株形态更加紧密(Hooks et al.,2021)。因此,不同波段的光对蔬菜作物有着不同的影响,目前主要的研究集中在红蓝光及UV 辐射。
光调控植物品质的课题因其具有重要的应用价值成为目前研究的热点之一,研究者对蔬菜作物中类胡萝卜素的研究更是较为热衷。本文概述了不同光质对蔬菜作物类胡萝卜素含量的影响,并重点从生物合成关键基因调控、翻译后调控及库源调控方面总结了光质对蔬菜作物类胡萝卜素的调控机制,旨在为设施蔬菜作物高效种植、产品器官品质光调控、植物照明灯具开发提供理论参考。
1 类胡萝卜素生物合成通路
1.1 前体的合成
自然界中存在两种途径合成类异戊二烯前体:二甲基烯丙基二磷酸(DMAPP)和异戊烯焦磷酸(IPP),即甲基赤藓糖醇途径(MEP)和甲羟戊酸途径(MVA)。MEP 途径主要发生在大多数真细菌、顶复门原生生物中(Jomaa et al.,1999;Nair et al.,2011),MVA 途径主要发生在古细菌、真菌和动物中(Braithwaite &Goodwin,1960;Horbach et al.,1993)。植物可以通过MEP 和MVA 途径产生类异戊二烯前体,并且MEP 途径主要发生在细胞器质体中,MVA 途径主要发生在细胞液中(Rodríguez-Concepción &Boronat,2002)。因此,在蔬菜作物中,这两种重要前体IPP 和DMAPP是通过质体中MEP 途径产生的,接着二者发生缩合形成类胡萝卜素及其他的一些萜类化合物(Rodríguez-Concepción,2010)。通过总结现有研究 报道(Domonkos et al.,2013),DMAPP 和IPP合成过程如图1 所示,主要经过以下几个步骤:首先D-葡萄糖经过一系列代谢过程形成丙酮酸,丙酮酸和3-磷酸甘油醛(G3P)在1-脱氧-D 木酮糖-5-磷酸合酶(DXS)的催化下形成1-脱氧-D-木酮糖-5-磷酸(DXP)。在该过程中,DXS 催化的反应是MEP 途径中的限速反应(Estevez et al.,2001),接下来DXP 通过一系列反应形成类异戊二烯重要前体DMAPP 和IPP。
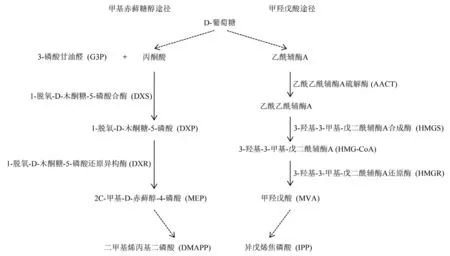
图1 植物中DMAPP 和IPP 合成通路
1.2 类胡萝卜素的合成
目前植物中类胡萝卜素合成一般通路已较为明晰,参阅相关研究报道(Domonkos et al.,2013),将其归纳为图2。DMAPP 和IPP 在牻牛儿基牻牛儿基二磷酸合成酶(GGPPS)催化下形成牻牛儿基牻牛儿基二磷酸(GGPP)。GGPP 是植物类异戊二烯代谢的关键中间体,也是类胡萝卜素、生育酚、叶绿素侧链、质体醌和植物激素等物质的重要前体(Moise et al.,2014)。合成的两分子GGPP 前体在八氢番茄红素合酶(PSY)作用下发生头对头缩合,形成15-顺式-八氢番茄红素(15-cis-phytoene)。该过程是类胡萝卜素生物合成的第1 步,并且是关键的限速步骤(Yuan et al.,2015a),PSY 是该过程中的限速酶。无色的15-顺式-八氢番茄红素在一系列关键酶的作用下经过去饱和、脱氢和异构化最终产生红色的全反式番茄红素(all-translycopene)。
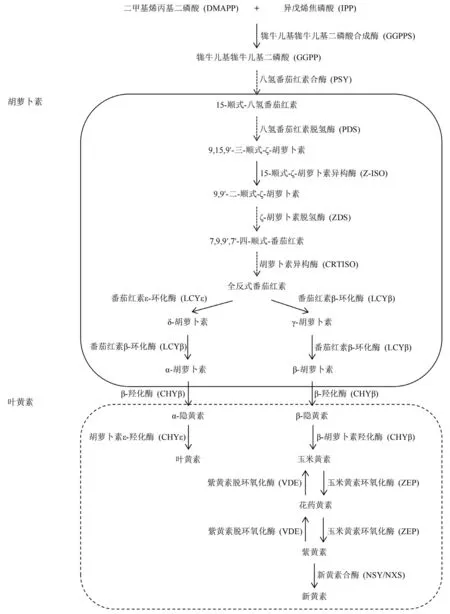
图2 高等植物中主要类胡萝卜素生物合成通路
番茄红素在植物中以全反式构型存在,并在番茄红素ε-环化酶(LCYE)或番茄红素β-环化酶(LCYB)作用下分别形成δ-胡萝卜素(δ-carotene)或γ-胡萝卜素(γ-carotene)。番茄红素的环化是类胡萝卜素代谢中的重要步骤,δ-胡萝卜素和γ-胡萝卜素的形成更是类胡萝卜素合成途径中1 个重要的分支点,β-环和ε-环的引入保证了植物中类胡萝卜素的多样性(Nisar et al.,2015)。随后δ-胡萝卜素和γ-胡萝卜素在LCYB 作用下进行环化,分别形成α-胡萝卜素(α-carotene)和β-胡萝卜素。
α-胡萝卜素和β-胡萝卜素在各种酶的催化下进一步羟基化形成叶黄素(xanthophyll)。叶黄素是植物光系统中主要的类胡萝卜素,它不仅赋予了植物特殊的颜色和气味,并且具有很强的光保护和抗氧化特性,保证了生命体维持正常的生长代谢(Ruban &Johnson,2010)。此外,在一系列关键酶的作用下,玉米黄素(zeaxanthin)、花药黄素(antheraxanthin)和紫黄素可以相互转换。叶黄素循环(xanthophyll cycle)系统中三者之间的相互转换,保证了植物在强光下可以有效增强自身光保护作用,以适应不同的光环境(Goss &Jakob,2010)。
2 光质对蔬菜作物类胡萝卜素含量的影响
2.1 蓝光的影响
蓝光在植物的生长发育进程中起着重要作用,它作为一种环境信号,能够增加植物对碳的同化,从而提高类胡萝卜素等次生代谢物合成的水平。因此,适量的蓝光对植物类胡萝卜素的积累是有益的。Gangadhar 等(2012)研究发现在蓝光下辣椒果实中类胡萝卜素含量显著增加,并且对叶绿素的生物合成十分有益。Johkan 等(2010)采用蓝光对红叶叶用莴苣幼苗进行处理,也发现植株叶片中类胡萝卜素含量显著升高;而处理后的幼苗置于自然白光下生长时,其叶片中类胡萝卜素含量略微降低。Lin 等(2021)采用不同比例的红、绿、蓝光对罗勒进行处理,发现适当增加蓝光比例,红∶绿∶蓝=1∶1∶1 时罗勒叶片中类胡萝卜素含量显著提升。
微型菜苗在色泽、质地、风味及营养成分上具有独特的优势,其富含氨基酸、维生素、多酚、色素和抗氧化物等营养物质,因此越来越受到广大人群的喜爱(Choe et al.,2018;Yadav et al.,2019)。Kopsell 和Sams(2013)研究发现花椰菜苗采前使用微量蓝光处理,可显著提高芽中类胡萝卜素、硫苷和矿物元素的含量,尤其是β-胡萝卜素和钾、镁、铁等元素。Samuolienė 等(2017)也指出在组合光谱中,随着蓝光强度增加,甜菜苗中α-胡萝卜素、β-胡萝卜素、叶黄素等类胡萝卜素的含量也逐渐上升。
通过改变蓝光的比例,来达到增加植株产量器官中类胡萝卜素含量的目的是一种可行的方式。但不可否认的是,提升植物营养品质不仅需要适当的光质条件,还需要适宜的光照处理时间。Metallo等(2018)研究发现,前期对羽衣甘蓝进行5%蓝光和95%红光处理22 d,而后适当增加蓝光的比例(20%)处理15 d,可以增加植株类胡萝卜素的含量。
植物的生长受到诸多因素的影响,光质作为主要影响因素之一,并不能单方面起到决定性的作用。考虑到品种效应及各项研究试验条件(如光强、光照时间、温湿度、CO2浓度等)存在的差异,不同光质下植物类胡萝卜素水平的变化也存在一定的差异。但总而言之,适量的蓝光处理对于大多数蔬菜作物类胡萝卜素的积累是有益的。
2.2 红光/远红光的影响
Lin 等(2021)研究发现,在组合光谱中,随着红光比例的上升,罗勒叶片中类胡萝卜素的含量逐渐下降。Liang 等(2021)也发现相对于纯蓝光,纯红光处理会显著降低黄瓜叶片中类胡萝卜素的含量。但Meas 等(2020)发现660 nm 红光可以显著增加苋菜中类胡萝卜素含量。Rahman 等(2021)采用70%红光和30%远红光对马铃薯植株进行处理,发现植株叶片中类胡萝卜素含量显著提高,更有助于植物光合作用的进行。这些结果表明红光/远红光对不同植物类胡萝卜素积累存在不同的效应,可能是不同蔬菜作物对红光/远红光的敏感性具有一定差异性。
光质处理与光周期密切相关,光周期的改变会直接影响植物某些次生代谢物的合成。Zou 等(2019)在对叶用莴苣的研究中发现,在红/蓝光谱中添加50 μmol·m-2·s-1的远红光处理16 h,会显著降低植株类胡萝卜素含量;而在同样的处理下,延长光周期1 h,则会轻微增加叶片中类胡萝卜素含量。
2.3 UV 辐射的影响
根据波段的不同,可将UV 分为3 个波段:UV-A(315~400 nm)、UV-B(280~315 nm)、UV-C(100~280 nm)。不同波段UV 对蔬菜作物类胡萝卜素的积累存在不同影响。Moreira-Rodríguez 等(2017)使用UV-A 对采前花椰菜苗进行短时间照射,发现可显著提高叶片叶黄素和新黄素的含量。Assumpção 等(2019)每天将叶用莴苣置于强度为0.5 kJ·m-2·d-1的UV-B 下照射1 h,两周后发现植株叶片类胡萝卜素含量显著上升。Zhang 等(2018)在自然光下补充UV-A 对番茄植株进行处理,发现可以显著提高植株叶片中类胡萝卜素含量。而Mariz-Ponte 等(2018)用UV-B 对番茄幼苗进行短时间处理,植株中类胡萝卜素含量却显著降低。这表明UV 波段及蔬菜作物的差异会引起植株类胡萝卜素积累的差异。
在蔬菜作物不同生长阶段使用不同UV 辐射进行处理,对植株类胡萝卜素含量的影响也不尽相同。Heinze 等(2018)报道,在芽期对普通白菜每天进行UV(UV-A+UV-B)辐射处理12 h,改变UV 强度并不会影响植株类胡萝卜素含量;随着UV 辐射时间的延长,发现低强度的UV(UV-A 1 W·m-2,UV-B 0.000 6 W·m-2)可以显著提升成熟期普通白菜叶片中类胡萝卜素的含量。一般来说,类胡萝卜素在植株中年期叶片中的含量高于早期发育阶段的叶片(Ibanez et al.,2008)。以上结果可能是由光质及植物发育阶段共同影响导致的。
目前有关UV-C 的研究主要集中在植株产量器官采后保鲜及贮存上。Khubone 和Mditshwa(2018)研究指出,采用强度为969.8 μW·cm-2的UV-C 对采后番茄处理5 min,可以显著提高果实中类胡萝卜素的含量,并且随着贮存时间的延长,果实中类胡萝卜素依然保持较高的水平。Gogo 等(2018)对采后苋菜进行UV-C 处理,发现相比于未经过UV-C 处理的植株,植株叶片中β-胡萝卜素和叶黄素的含量分别升高了34%和32%,并且随着辐射时间延长至4 d,β-胡萝卜素的含量提高了68%。然而,叶用莴苣采前进行UV-C 辐射处理却产生了不同的结果。Vàsquez 等(2017)发现采用高强度的UV-C 照射叶用莴苣,会显著降低叶片中类胡萝卜素的含量,而在低强度UV-C 处理下其类胡萝卜素含量并没有发生明显改变。因此,UV 辐射对植株类胡萝卜素含量的影响存在一定的剂量效应,与蔬菜作物的品类及发育阶段也有很大的关系。
2.4 其他光的影响
Kaiser 等(2019)研究指出,采用组合光谱对番茄进行处理,当保持红蓝光比例相似时,随着绿光强度的升高,其中部叶总类胡萝卜素的含量也随之显著升高。此外,Zhang 等(2018)在自然光下补充黄光对番茄植株进行处理,同样发现其可以显著提高植株叶片中类胡萝卜素含量。这可能是不同光质处理下,植物自身合成更多的类胡萝卜素,以便更有效地吸收其他光(如绿光和黄光)。Samuolienė 等(2019)发现,在红蓝光谱中添加其他光,如绿光或黄光或橙光,可以显著提高芜菁芽中β-胡萝卜素的含量;在对花椰菜幼苗及结球甘蓝进行相同的光照处理时,却发现其幼苗中β-胡萝卜素的含量显著降低。与上述研究结果类似,不同光质处理对植物类胡萝卜素积累产生的影响不尽相同。此外,如何利用不同的光质来维持蔬菜作物采后产品器官的营养成分也是迫切需要解决的问题。据相关报道,Kokalj 等(2016)对采后甜椒进行黄光处理,可以显著提升甜椒β-胡萝卜素和叶黄素含量,更有利于果实的贮存,减缓营养成分的流失。
2.5 小结
根据前人相关研究,总结了部分蔬菜作物在不同光质处理下类胡萝卜素积累的情况(表1),总的来说,不同波长的光对植株化学成分变化和果实颜色转变的影响取决于植物的种类、栽培方式及不同组织中色素的相对含量。

表1 不同光质对蔬菜作物类胡萝卜素含量的影响
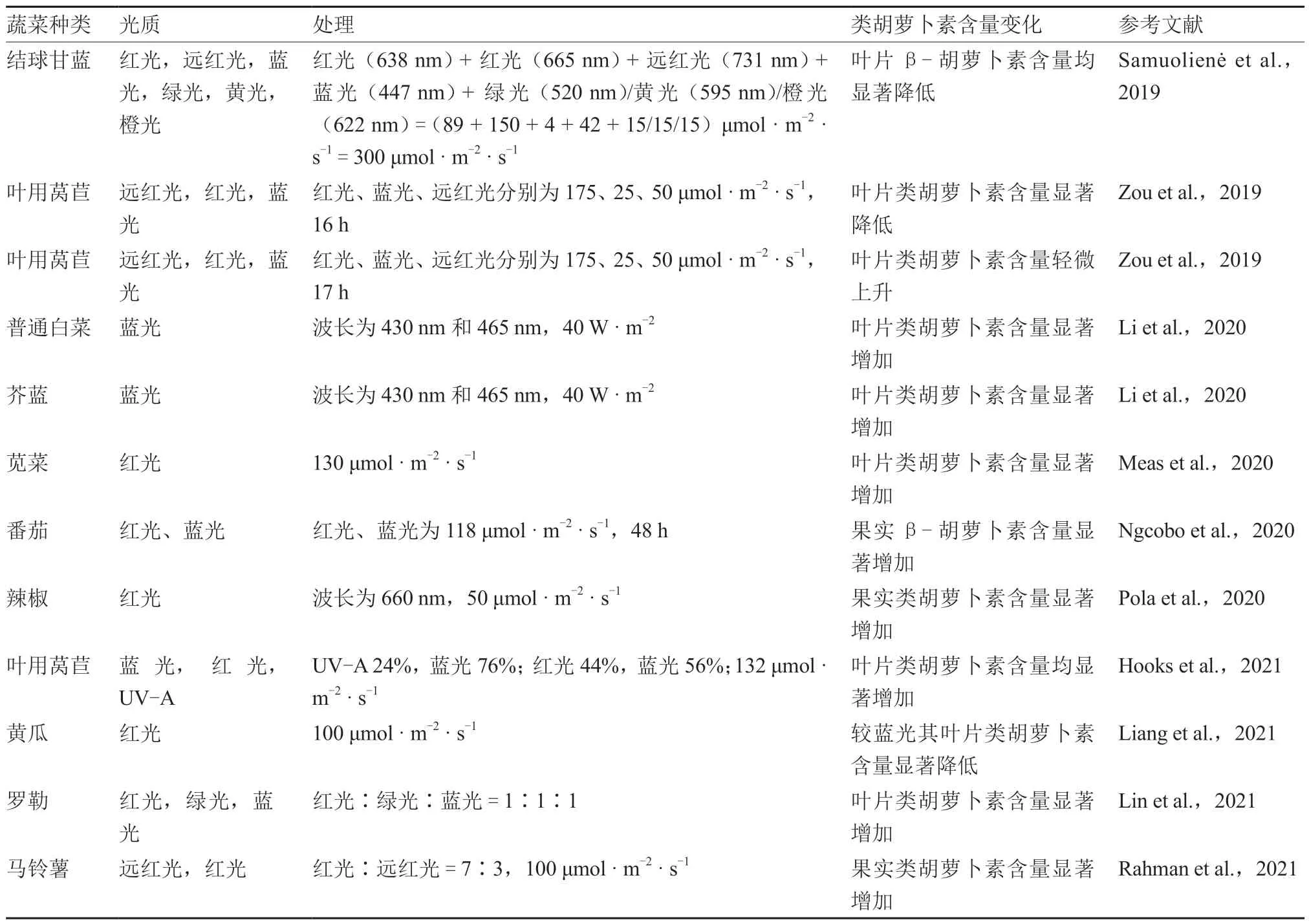
续表
3 光质对蔬菜作物类胡萝卜素合成调控的影响
3.1 生物合成关键基因调控
随着对光调控植物品质研究的深入,越来越多的研究者发现调控光信号的成分是改变植物品质的一种重要方式。植物主要通过4 种光感受器来接收不同波长的光,分别是感知红光及远红光的光敏色素(PHYs)、感知蓝光及UV-A 的隐花色素(CRYs)和向光素(PHOTs),以及感受UV-B 的UVR8(UV RESPONSE LOCUS 8)受体。不同波长的光通过启动光感受器的信号级联而影响植物类胡萝卜素代谢途径中关键酶基因的表达,进而调控类胡萝卜素的生物合成。Li 等(2021a)研究发现,绿豆芽中DXS 在光照和黑暗下具有不同的活性。相比于黑暗,在光照条件下番茄中抑制PSY表达的转录因子ERF.E4(ethylene response factor.e4)水平会显著降低,表明光对类胡萝卜素合成基因具有一定的调控作用;遮光处理显著降低了甜椒果实类胡萝卜素合成基 因PSY、PDS、ZDS、ZISO、LCYB1和CHY2等的表达水平(Yoo et al.,2020)。此外,相较于白光,红光下番茄果实中PSY和CRTISO基因表达水平显著上调,而蓝光下其显著下调,表明不同光质对植物中类胡萝卜素生物合成基因的影响不尽相同(Li et al.,2021b)。
转录因子ELONGATED HYPOCOTYL 5(HY5)及PHYTOCHROME-INTERACTING FACTORs(PIFs)已被确认为光信号通路的关键成分(Chen et al.,2013)。HY5 通过与该通路中一些成分如PIF1a(Llorente et al.,2016)、DAMAGED DNA BINDING PROTEIN 1/HIGH PIGMENT 1(DDB1/HP1)(Wang et al.,2008)、DEETIOLATED 1/HIGH PIGMENT 2(DET1/HP2)(Davuluri et al.,2005)、CULLIN 4(CUL4)(Wang et al.,2008)和COP1-like(Liu et al.,2004)等协同作用以调节番茄中类胡萝卜素的积累。番茄hy5突变体果实中PSY1、Z-ISO、Crt-ISO1、LCY-B和VDE基因显著下调,表明番茄果实中类胡萝卜素的合成受到抑制(Wang et al.,2021)。在UV-B 诱导的信号通路中,拟南芥光受体UVR8 直接与CONSTITUTIVELY PHOTOMORPHOGENIC1(COP1)相互作用以激活转录因子HY5(Yin et al.,2015;Camacho et al.,2019),进而引起PSY、ZDS和BCH1/2等合成类胡萝卜素关键基因表达量升高(Emiliani et al.,2018)。
在拟南芥的光信号转导过程中,BBX 家族基因具有重要的调控作用。如BBX21 通过与HY5启动子中的T/G-box基序结合并激活其表达(Xu et al.,2018);BBX25 通过与HY5 形成失活的异源二聚体,负向调控BBX22基因的表达(Gangappa et al.,2013);而BBX22 参与了拟南芥光形态建成(Chang et al.,2011)。Xiong 等(2019)在番茄中发现一种名为BBX20 的蛋白,它直接与PSY1启动子结合并激活其表达,并且BBX20 蛋白受CRL4(由CUL4、DDB1 和DET1 组成)复合体的调控。Lira 等(2020)也发现光敏色素(PHY)介导的光信号参与了番茄BBX20基因的转录调控。PHY 通过感知红光/远红光比例来调节自身的活性状态。Bou-Torrent 等(2015)发现在低比例红光/远红光下,拟南芥中PIF1 及相关PIFq 蛋白抑制了PSY基因的表达;此外,PHYTOCHROME-RAPIDLY REGULATED 1(PAR1)可与PIF1 相互作用从而诱导PSY表达。
目前,虽然部分光质影响植物类胡萝卜素积累的信号通路已较为明晰(图3),但还存在一些不足。一是光质介导植物类胡萝卜素生物合成是一系列复杂的过程,其中涉及许多转录因子的参与。目前也发现了很多类胡萝卜素调控相关的转录因子,但是它们是通过什么样的方式来发挥作用以及光对这些转录因子是如何进行调控的在很大程度上是未知的。二是类胡萝卜素最终在质体中的净积累量是合成、降解和稳定储存的结果。并且一些营养元素对类胡萝卜素积累也有影响。因此,光质对植物体内类胡萝卜素调控相关转录因子及相关离子转运蛋白的影响还需要进一步的研究。

图3 部分光质调控植物类胡萝卜素合成的机制
3.2 翻译后调控
翻译后调控对于维持植物中类胡萝卜素生物合成关键酶的功能至关重要,其主要是通过蛋白水解及蛋白与蛋白相互作用从而对该代谢途径中的酶进行调控,进而影响植物体中类胡萝卜素的含量及组成。光参与了植物类胡萝卜素合成关键酶基因(如PSY和DXS)的翻译后调控。拟南芥PSY基因具有两种不同的5′UTRs 差异剪接体(ASV)。光照下ASV2 诱导水平显著增加,并伴随着PSY 蛋白水平及PSY活性的提升;ASV1 在幼苗脱黄化过程中具有对PSY诱导的作用(Álvarez et al.,2016)。Welsch 等(2000)研究发现与黑暗中生长的白芥相比,远红光下白芥植株中PSY 蛋白的水平显著升高,但PSY的活性却没有明显改变;此外,光照条件下PSY与类囊体膜结合,其活性迅速增加,这主要归功于NADPH 作用。高光照强度条件下,植物蛋白质会发生错误折叠或形成蛋白聚集体。拟南芥中J-蛋白J20 可与错误折叠的DXS 相互作用,并将其靶向HSP70 系统进行重新折叠以恢复功能(Pulido et al.,2013)。ORANGE(OR)蛋白作为PSY翻译后主要的调控因子,可与其直接结合以稳定类胡萝卜素的生物合成。与DXS 类似,发生错误折叠或形成蛋白聚集体的PSY 可被Clp 蛋白酶所降解,而OR 蛋白通过与这些PSY 蛋白相互作用促使其进行重新折叠(D′Andrea &Rodriguez-Concepcion,2019)。
3.3 库源调控
蔬菜作物中类胡萝卜素的积累不仅取决其生物合成,细胞内类胡萝卜素的储存方式及空间分布对其也有很大的影响。质体的超微结构对类胡萝卜素的积累具有重要的作用(Llorente et al.,2017),它决定了植物各组织中类胡萝卜素的相对含量(Sun et al.,2018)。在番茄红熟期,果实中叶绿体逐渐转化为有色体并且颜色发生转变,由绿色变成黄色或红色(Suzuki et al.,2015)。这主要是红光及远红光促使质体的不同分化所致(Quian-Ulloa &Stange,2021)。Usami 等(2004)发现,PHYs 和CRYs 可以调节拟南芥根系中白色体向叶绿体的转化,并且蓝光的诱导作用比红光更强。蛋白质组学研究定位了大部分的类胡萝卜素合成酶在叶绿体被膜上,部分酶存在于类囊体中(Ytterberg et al.,2006;Joyard et al.,2009)。叶绿体信号识别颗粒(scSRP54,ffc)和(scSRP43,Chaos)及其受体(cpFtsY)可将捕光蛋白靶向类囊体膜。与野生型相比,拟南芥cpftsy突变体中叶黄素含量下降了67%,β-胡萝卜素含量下降了约80%,ffc/chaos突变体发生相似变化(Tzvetkova-Chevolleau et al.,2007)。细胞器中类胡萝卜素合成酶的定位与代谢产物并不完全重叠,质体小球与类囊体合成密切相关,可能参与代谢物的输送(van Wijk &Kessler,2017)。
一般来说,有色体是类胡萝卜素储存的主要场所。在胡萝卜根部和番茄果实中,叶绿体可以分化为有色体,叶绿体的发育及有色体的分化为高水平的类胡萝卜素积累提供了可能,并伴随着PSY基因活性的提高(Llorente et al.,2017)。Arias 等(2020)发现PHYA、PHYB、PIF3、PAR1、CRY2、FAR1和COP1等基因参与胡萝卜贮藏根中有色体分化的调控。从花椰菜突变体中分离出的OR基因编码一种富含半胱氨酸的DnaJ 锌指蛋白,它在多种植物中参与有色体的合成(Lu et al.,2006;Li et al.,2012;Ellison et al.,2018)。基于甜瓜品种golden的SNP,拟南芥OR 蛋白相同位点单个氨基酸的取代显著提高了β-胡萝卜素含量,并且与有色体的合成有关(Yuan et al.,2015b)。Arias 等(2020)研究表明,光负调控OR基因的表达,导致胡萝卜贮藏根中有色体丰度降低。
Bianchetti 等(2018)研究发现PHYA 通过CRFs、TRRs 和COP1 促进番茄果实中质体的分化与成熟,并伴随着类胡萝卜素生物合成增加。番茄hp1和hp2突变体中质体的数量和大小显著增加,同时类胡萝卜素的含量显著上升(Cruz et al.,2018;Wang et al.,2019)。番茄hp2突变体中,光通过调控乙烯和生长素途径转录因子ERF.E4 和ARF2a/b 进而调控类胡萝卜素的生物合成(Cruz et al.,2018)。此外,拟南芥根部质体的发育还受到生长素和细胞分裂素信号的调控,PHYs 及一些转录因子(如HY5 和GLK2 等)参与了该信号途径(Kobayashi et al.,2013;Kobayashi et al.,2017)。植物中质体的分化与类胡萝卜素积累除了受到光的调控外,二者间同样存在复杂的关系。Fraser 等(2007)指出,在过表达PSY1番茄植株中,随着果实的发育,有色体结构过早形成。而Lu 等(2017)在研究中却发现通过补光增加柑橘中胡萝卜素的积累,可以引起果肉中有色体的发育,类胡萝卜素的积累可能受到植物自身新陈代谢的限制。
4 结论与展望
近些年来,有关蔬菜作物中类胡萝卜素代谢机理得到了广泛的研究。但是,对于光质调控植物类胡萝卜素代谢的分子机制研究甚少。本文综述了光质对蔬菜作物类胡萝卜素的影响及调控机制,并得出以下结论:①适量的蓝光有益于大多数蔬菜作物类胡萝卜素积累。可以根据一定的需求,有目的地补充蓝光以提升植株类胡萝卜素的含量。②其他光(红光/远红光、UV 等)对蔬菜作物类胡萝卜素积累的影响存在一定差异性,需结合作物品类、植株生长阶段及处理方式具体而定。③可以根据作物品种采取相应的补光措施,以维持植物产量器官采后类胡萝卜素含量的稳定。④PSY是蔬菜作物类胡萝卜素生物合成的关键酶基因,通过不同的作用方式上调PSY的表达,可促进类胡萝卜素的生物合成。类胡萝卜素主要在有色体中进行合成。有色体的发育保证了类胡萝卜素的储存。适量的光照有益于PSY的表达及白色体向有色体的转化。此外,高光照强度会抑制类胡萝卜素生物合成关键酶的活性,导致其发生错误折叠或不进行折叠。
因此,光源的强度、光质及光周期等是决定植物中类胡萝卜素生物合成的关键因素。但目前,关于光质调控蔬菜作物类胡萝卜素积累方面的研究还存在许多问题仍待解决:①番茄作为研究蔬菜作物类胡萝卜素代谢的模式植物,近些年来对它的研究不断深入。但是有关该方面的研究结果并不都是集中在番茄上,所得结论并不具有代表性。②目前大部分的研究集中在红蓝光及UV-B 上,有关UV-A 及其他波长的光(绿光、黄光等)研究甚少。虽然已经有人提出了绿光光受体的设想,但这些光影响植物类胡萝卜素合成的分子机制还需进一步研究。③光质调控植物类胡萝卜素生物合成是一系列复杂的过程,其中涉及多个转录因子的参与。HY5 作为植物光形态建成的关键调控因子,参与了植物体内多种代谢过程,包括类胡萝卜素的生物合成。在番茄中也发现了除HY5 以外的类胡萝卜素代谢通路。因此,亟需寻找类似于HY5 的转录因子,并阐明其分子机制。④有关光质方面的研究大多是单一因素,设施园艺中LED 的使用不可避免的改变了植物局部温度,故可以考虑光温耦合方面的研究,并寻找相应的转录因子或转录因子复合体。⑤目前,有关光质对蔬菜作物类胡萝卜素合成关键酶基因翻译后调控及库源调控方面的研究甚少。OR 蛋白作为PSY翻译后的主要调控者,还参与了质体的发育。然而,OR 蛋白是如何对这些过程进行调控的目前还没有明确的结论。
以上几方面的研究将更全面地解析光质调控蔬菜作物类胡萝卜素积累的分子机制,从而为实现定向调控植物中类胡萝卜素等次生代谢物积累的目标提供一定的可能性,并为蔬菜作物高品质选育策略提供理论参考。
猜你喜欢
学与玩(2022年6期)2022-10-28
四川林业科技(2022年1期)2022-03-22
上海农业学报(2022年1期)2022-03-08
——记嘉荫县红光乡燎原村党支部
奋斗(2020年24期)2021-01-16
活力(2019年17期)2019-11-26
新农村(浙江)(2019年5期)2019-05-29
浙江农业学报(2017年1期)2017-05-17
中国酿造(2016年12期)2016-03-01
中国果菜(2016年9期)2016-03-01
中国果业信息(2015年12期)2015-01-24
