
转录因子TaMADS2在小麦-叶锈菌互作中的功能研究
2023-05-11 13:26王率伍赵世佳刘子豪路亚南马保站肖继斌郑文明
中国农业大学学报 2023年5期
王率伍 秦 昭 赵世佳 刘子豪 路亚南 马保站 肖继斌 程 琨 郑文明 刘 娜
(河南农业大学 生命科学学院/小麦玉米作物学国家重点实验室,郑州 450002)
小麦叶锈病是由叶锈菌(Pucciniatriticina)侵染引起的在世界范围内广泛分布的真菌病害,发生严重时可造成小麦5%~45%的减产[1]。小麦叶锈菌是专性寄生的活体营养型真菌,该菌分布范围广,发生频繁,在世界范围内造成小麦产量的严重损失,包括亚洲、北美洲、欧洲、澳洲以及非洲等地区[2]。锈病防治最经济有效的方法是抗病品种的培育[3],因此新型叶锈病抗性基因的挖掘对减轻叶锈病的危害以及实现叶锈病的持久防治具有重要意义。
在长期进化过程中,植物形成了一套多层次的识别防御系统,从而抵御病原菌的侵染。植物可以在病原菌的侵染部位启动超敏反应(Hypersensitive response,HR)和局部程序性细胞死亡[4],从而阻止病原菌的进一步繁殖与扩散。超敏反应通常包括病程相关基因(Pathogenesis-related,PR)表达、活性氧(Reactive oxygen species,ROS)爆发以及信号分子的传递等[5]。病程相关蛋白(Pathogenesis-related proteins,PRs)是一类水溶性蛋白,主要功能包括攻击病原物、降解病原物毒素以及抑制病毒外壳蛋白同植物受体结合等[6]。PR1被证实具有抑制病毒扩展、抵抗真菌入侵以及保护植物抵御逆境胁迫等功能[7-8]。PR2基因编码β-1,3-葡聚糖酶,可以与几丁质酶一起降解真菌细胞壁,从而造成菌丝体顶端膨胀、破碎,最终死亡[9-10]。病程相关基因PR1和PR2是植物抗病反应的标志基因,检测这些基因的表达可侧面反应植物抗病反应的强弱。病原菌侵染引起的HR反应会促进活性氧(O2-和H2O2)的产生,不断积累的活性氧可以进一步刺激产生效应子触发的免疫反应(Effectors-triggered immunity,ETI)。ROS在细胞中积累能够控制和抑制病原菌的生长,同时可作为细胞壁内的抗菌小分子抑制病原菌的侵入,并作为次级信使触发额外的免疫反应,如基因表达或者气孔关闭[11-12]。
病毒诱导的基因沉默(Virus-induced gene silencing,VIGS)是一种利用反向遗传学的原理研究基因功能的方法,将目的基因特异性片段插入病毒载体后侵染宿主植物,导致目的基因局部双链RNA形成,从而引起目的基因的沉默[13]。VIGS技术避免了对寄主植物进行遗传转化,研究周期短,是一种简单有效的基因功能研究方法,从而极大的推动了植物基因组功能研究的发展。最早追溯到1995年,Kumagai等[14]首次将插有一段八氢番茄红素脱氢酶(Phytoene desaturase,PDS)的重组烟草花叶病毒(Tobacco mosaic virus,TMV)导入本氏烟中,成功的沉默了烟草的PDS基因。由于PDS是类胡萝卜素合成所必需的酶,其基因沉默会引起所感染植物的叶片产生光漂白表型[15],因此常被用来作为VIGS体系的指示基因。张蕊等[16]利用VIGS技术沉默TaSPX3基因的表达,发现TaSPX3基因正向调控小麦对叶锈菌的抗性;宋姗姗等[17]通过BSMV-VIGS技术证明TaRanGAP2基因在小麦抵抗叶锈菌侵染的HR反应中发挥重要的作用;蒋军等[18]利用VIGS技术沉默TaCTSB基因的表达,减弱了小麦对条锈菌的抗性。Guo等[19]通过VIGS技术沉默TaBln1的表达增强了小麦对条锈菌的防御反应;而Wan等[20]发现瞬时沉默TaRaf46可以抑制小麦叶片中条锈菌的侵染。另外有报道称,在寄主小麦中通过VIGS能够沉默小麦条锈菌致病基因的表达,从而显著降低条锈菌的危害[21]。
转录因子在生物胁迫与非生物胁迫的信号通路中发挥关键作用。MADS-box转录因子是一类广泛存在于真核生物中的古老转录因子家族,参与植物生长的各个环节,在植物的生长发育、信号转导以及胁迫响应中发挥不可替代的作用[22]。普通六倍体小麦的全基因组表达谱分析发现,MADS-box家族多种转录因子在病原菌(赤霉菌、叶枯菌、条锈菌以及白粉菌)诱导下呈差异表达,因此MADS-box转录因子可能在小麦抵抗病原菌的侵染过程中发挥作用[23]。Guo等[24]研究发现I型MADS-box基因TaMADS2受条锈菌侵染后,在小麦与条锈菌亲和/非亲和互作早期上调表达,36和48 h之后出现下调表达,推测该基因可参与调控小麦对条锈菌的抗性。
本试验前期发现叶锈菌与小麦的亲和/非亲和互作早期TaMADS2基因的表达量均受到叶锈菌的诱导呈上调表达,且在非亲和互作中TaMADS2的上调表达时期较亲和互作更早。目前,关于TaMADS2参与小麦抗叶锈病过程的作用机制研究尚未见报道。因此,本研究拟利用BSMV-VIGS技术沉默小麦中TaMADS2基因的表达,对沉默植株接种叶锈菌后进行研究,旨在明确TaMADS2在小麦受叶锈菌侵染过程中的功能,为进一步阐明TaMADS2基因的功能机制奠定基础。
1 材料与方法
1.1 试验材料
供试小麦叶锈菌生理小种为Pt15,单孢菌系,该菌系为实验室前期鉴定并保存;供试小麦品种为‘郑麦9023’(ZM9023,Pt15非亲和互作)和‘中国春’(CS,Pt15亲和互作)。
1.2 大麦条纹花叶病毒(Barley stripe mosaic virus, BSMV)介导的TaMADS2基因沉默
本试验中所用BSMV的α、β和γ载体由河南农业大学农学院王道文教授课题组惠赠。TaMADS2基因cDNA全长822 bp,由试验前期从‘郑麦9023’中克隆得到。利用带有NheⅠ酶切位点的引物TaMADS2-VIGS-F/R从本研究已有的TaMADS2-T重组质粒上扩增一段204 bp的特异性片段作为VIGS的靶标序列,使用NheⅠ将γ载体酶切线性化,使用一步克隆法构建TaMADS2的VIGS重组载体γ-TaMADS2。
将病毒载体α、γ(Empty vector)、γ-PDS以及重组载体γ-TaMADS2使用Mlu Ⅰ进行线性化,β使用Spe Ⅰ进行线性化,使用RiboMAXTM-T7试剂盒进行体外转录。转录产物定量后分别将γ、γ-PDS、γ-TaMADS2与α、β等体积混合,组装为BSMV:EV、BSMV:PDS和BSMV:TaMADS2这3种病毒,采用摩擦接种的方法接种于二叶一心期小麦的第二片叶。接种BSMV:PDS的小麦作为阳性对照来观察沉默效果,PDS为八氢番茄红素脱氢酶基因,若PDS被沉默则小麦叶片会出现白化表型;接种BSMV:TaMADS2的小麦作为试验组,检测TaMADS2基因沉默小麦对叶锈菌的抗性变化;空质粒BSMV:EV为阴性对照。病毒接种后第14天观察接种BSMV:PDS小麦叶片的白化现象。将叶锈菌夏孢子Pt15均匀接种于BSMV:TaMADS2和BSMV:EV小麦叶片上。在接种叶锈菌后分别取样进行后续试验,并在接种叶锈菌后的第10天观察小麦叶片感病表型,分析TaMADS2基因沉默对于小麦抗病性的影响。
1.3 qRT-PCR检测
接种叶锈菌后第12天分别对沉默组和对照组取样,使用Trizol Reagent(CW0580S)和HiScript Ⅱ QRT SuperMix for qPCR(Vazyme,R223-01)分别提取样品RNA并反转为cDNA。使用qRT-PCR检测TaMADS2、PR1和PR2的相对表达量,分析TaMADS2的沉默效率以及叶锈菌侵染早期TaMADS2基因沉默对病程相关基因的转录水平影响。
接种叶锈菌10 d后分别对试验组和阴性对照组取样。提取样品DNA后通过绝对定量测定各样品中叶锈菌单拷贝基因PtRTP1和小麦单拷贝基因TaEF1的CT值,代入前期测定的特异性标准曲线[25]得到2个基因的拷贝数,计算PtRTP1和TaEF1基因的比例,获得样品的真菌生物量[26]。
1.4 组织学与活性氧染色观察
接种叶锈菌后的24和48 h分别对沉默组和对照组小麦叶片进行随机取样,参考肖继斌[27]所列方法使用WGA-Alexa-488荧光染料对沉默组和对照组24及48 h样品进行处理,用二氨基联苯胺(DAB)活性氧染料对沉默组和对照组的48 h样品进行处理,在荧光显微镜下观察菌丝生长与活性氧的积累情况。
2 结果与分析
2.1 VIGS重组质粒的构建
以TaMADS2-T载体为模板,使用引物TaMADS2-VIGS-F/R进行目的片段的扩增,克隆得到204 bp的特异靶标序列。对扩增产物进行回收,将回收产物与线性化γ载体连接并转化大肠杆菌,挑取单克隆菌落,使用VIGS-text-F/R进行单克隆测序,与靶标序列一致即重组质粒构建完成可进行下一步操作。将验证成功的菌落的质粒进行酶切线性化,重组质粒片段小于线性化质粒,说明重组质粒成功线性化。将线性化的质粒进行体外转录,获得病毒α、β、γ、γ-PDS以及γ-TaMADS2的RNA,用于后续试验。
2.2 BSMV-VIGS沉默体系效果检测
为验证本试验BSMV-VIGS沉默体系是否有效,在感染病毒14 d后观察BSMV:PDS沉默表型。如图1(a)所示,‘中国春’和‘郑麦9023’小麦叶片在接种BSMV:PDS病毒后出现白化,而作为阴性对照的BSMV:EV未出现变化,证明本研究所用沉默体系能够有效沉默小麦基因。利用qRT-PCR检测感染BSMV病毒的小麦在接种叶锈菌后12 h叶片中TaMADS2基因的表达水平,进一步证实TaMADS2基因是否被成功沉默。如图1(b)和(c)所示,与感染BSMV:EV的小麦相比,感染BSMV:TaMADS2病毒的小麦的TaMADS2基因表达量显著降低,结果表明感染BSMV:TaMADS2病毒有效的沉默了‘郑麦9023’和‘中国春’中的TaMADS2基因表达。
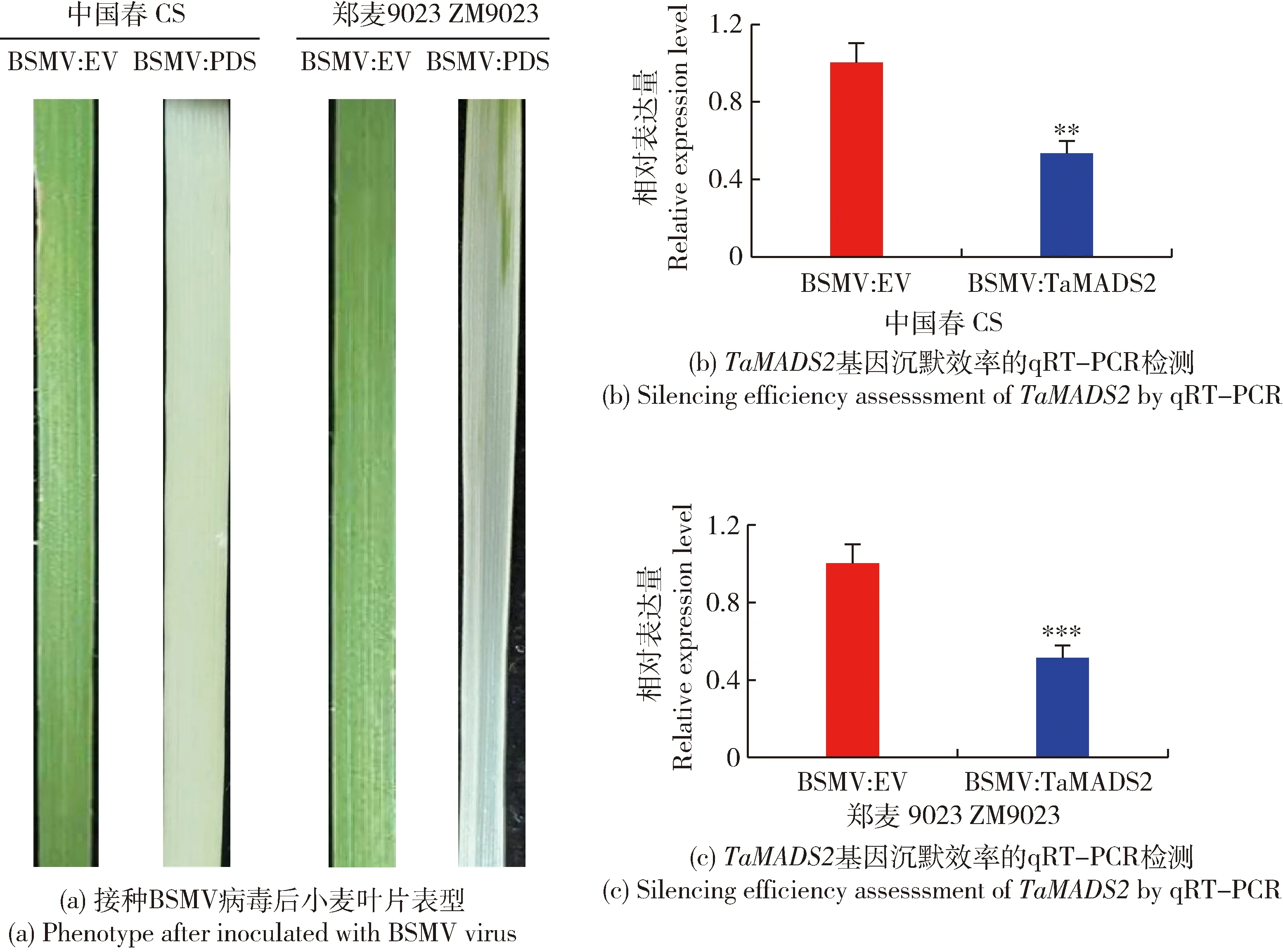
BSMV:EV为阴性对照,BSMV:PDS为PDS基因沉默的小麦植株,BSMV:TaMADS2为TaMADS2基因沉默的植株。显著性分析(*P<0.05,**P<0.01,***P<0.001)。下同。BSMV: EV is the negative control, BSMV: PDS is the PDS-silenced wheat plants, BSMV: TaMADS2 is the TaMADS2-slienced wheat plants. Significance analysis (*P<0.05, **P<0.01, ***P<0.001). The same below.图1 接种BSMV病毒的小麦叶片表型及沉默效率检测Fig.1 Phenotype and silencing efficiency of wheat leaves inoculated with BSMV virus
2.3 TaMADS2基因沉默后小麦抗病性分析
为解析TaMADS2基因在小麦抗叶锈病中的功能,分别对感染BSMV:EV和BSMV:TaMADS2病毒的小麦叶片接种叶锈菌,在第10天,观察叶片感病表型。如图2(a)所示,2个不同品种小麦感染病毒后,BSMV:TaMADS2的小麦叶片明显比BSMV:EV的小麦叶片产生更多的叶锈菌孢子堆。对2种不同小麦叶片进行分子病情指数检测,与表型观察结果相同,BSMV:TaMADS2小麦叶片的真菌生物量高于BSMV:EV的小麦叶片(图2(b)和(c)),表明TaMADS2基因的沉默降低小麦对叶锈病的抗病性。
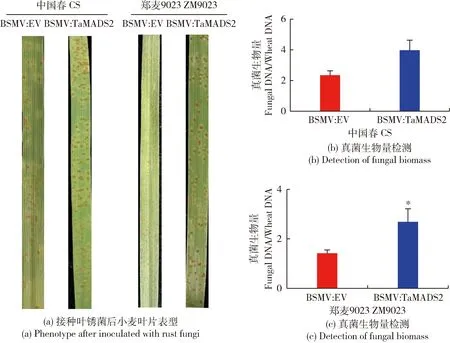
图2 TaMADS2沉默植株及对照植株接种叶锈菌后的叶片表型及真菌生物量检测Fig.2 Leaf phenotype and detection of fungal biomass of TaMADS2-slienced plants and control plants
对叶锈菌侵染的小麦叶片经DAB染色后在组织学水平观察活性氧积累上的变化,经WGA染色,在组织学水平观察叶锈菌在TaMADS2沉默植株中的生长情况。DAB染色后的活性氧观察结果如图3所示,在48 h时BSMV:EV样品内活性氧积累量显著大于BSMV:TaMADS2样品。WGA染色后的组织学观察结果如图4所示,在24和48 h,处理组BSMV:TaMADS2和对照BSMV:EV均可观察到气孔下囊和入侵菌丝,但处理组BSMV:TaMADS2中的气孔下囊较对照BSMV:EV相比更大且入侵菌丝更长,表明TaMADS2基因的沉默可能促进叶锈菌侵染初期入侵菌丝的扩展。以上结果表明沉默TaMADS2基因可降低小麦对叶锈病的抗性。
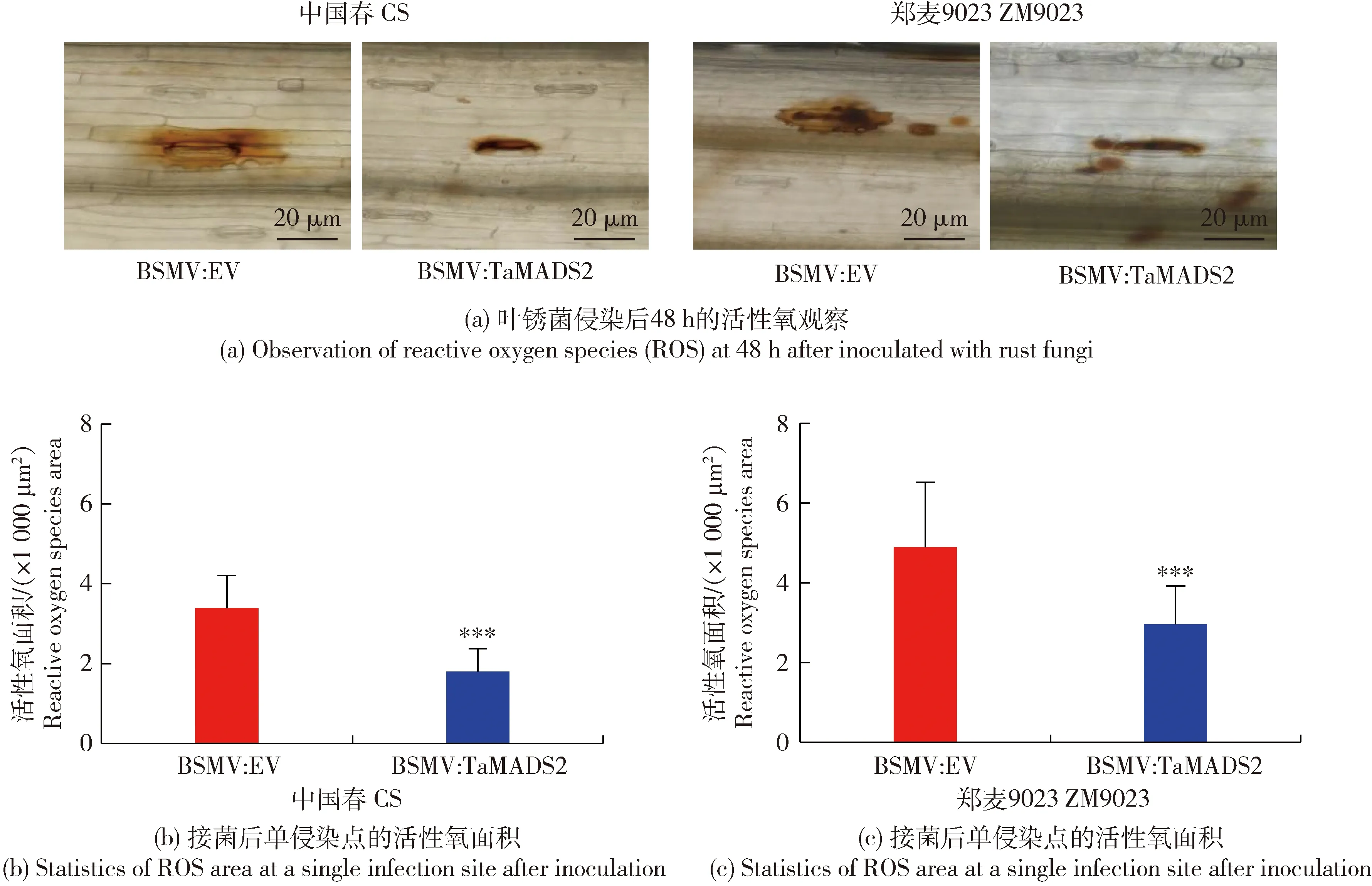
图3 叶锈菌侵染后的活性氧观察及数据统计Fig.3 Observation of reactive oxygen species (ROS) and data statistics after inoculated with rust fungi
在接种叶锈菌后的12 h取样检测病程相关基因以及活性氧清除相关基因的转录水平变化,其中PR1、PR2和CAT基因的表达量变化显著。如图5所示,同对照组BSMV:EV相比,感染BSMV:TaMADS2的2种小麦的PR1基因的表达量均降低,其中亲和小麦品种‘中国春’中PR1表达量显著降低90.22%,非亲和小麦‘郑麦9023’降低14.73%;感染BSMV:TaMADS2的2种小麦中PR2基因的表达量显著低于感染BSMV:EV的小麦,分别下降46.67%和36.38%,表明TaMADS2的沉默显著抑制了2个品种小麦中PR2的表达。CAT基因的表达量在感染BSMV:TaMADS2的小麦中相对于感染BSMV:EV的小麦分别降低17.80% 和24.15%,表明在叶锈菌侵染早期,TaMADS2基因可能通过活性氧清除酶(TaCAT)维持植株体内活性氧的动态平衡。对这2种小麦表达分析发现,TaMADS2基因沉默后在亲和/非亲和小麦中PR1、PR2和CAT的表达量均呈下调,表明TaMADS2基因可能通过PR1、PR2和CAT调控小麦叶锈菌抗性。
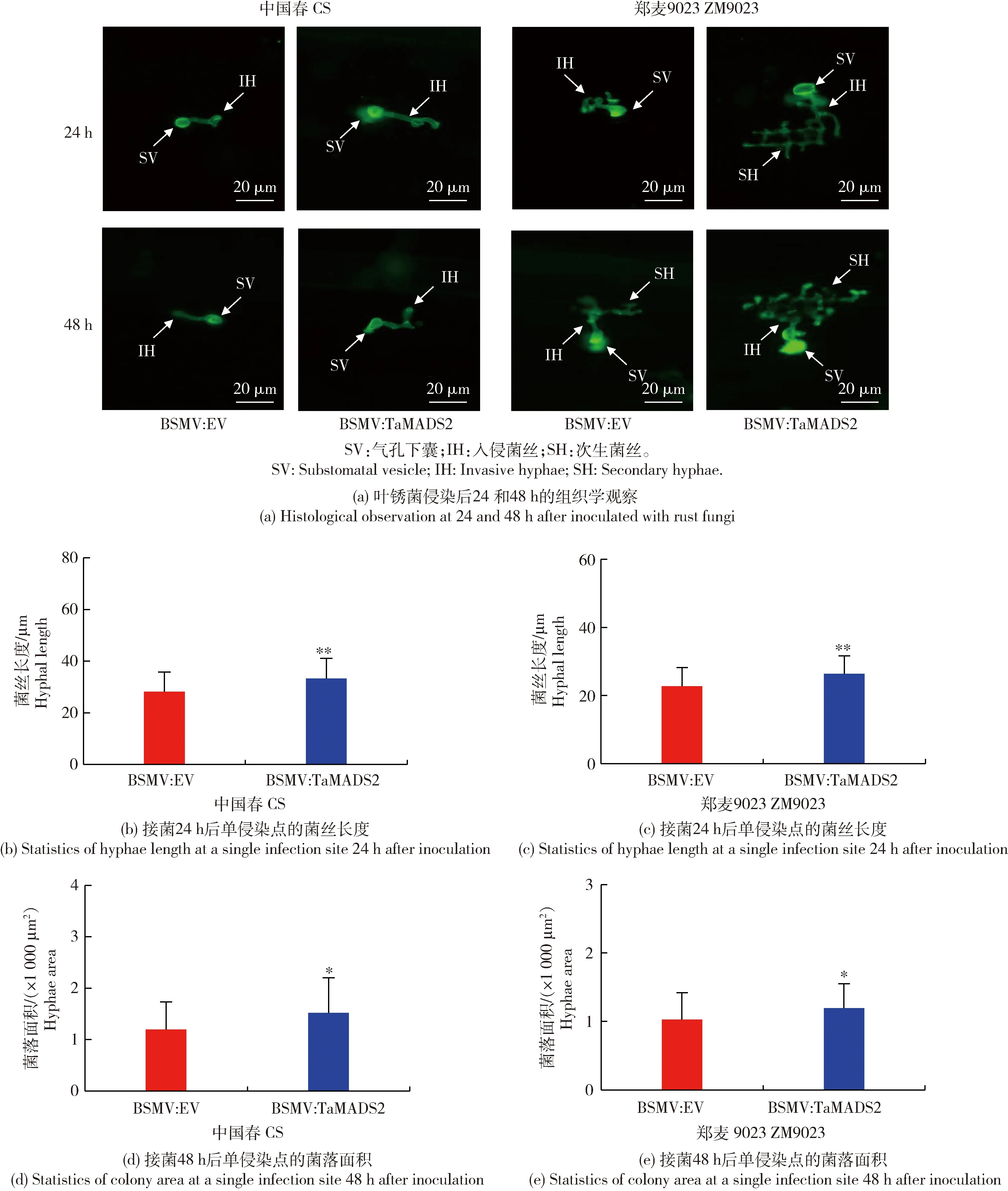
数据为40个数据的平均值。Values are the average of 40 data.图4 沉默TaMADS2后叶锈菌的组织细胞学分析Fig.4 Histocytology analysis of rust fungi growth in TaMADS2-silenced plants
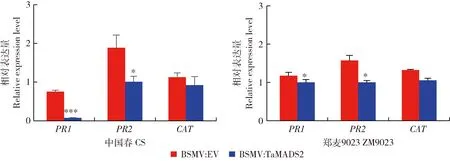
图5 TaMADS2沉默植株和对照植株接种叶锈菌后PR1、PR2和CAT基因的相对表达量Fig.5 The relative expression levels of PR1、PR2 and CAT genes in TaMADS2-silenced plants compared to control plants after inoculated with rust fungi
3 结论与讨论
植物在生长发育的过程中为应对多变的环境进化出了复杂的调控网络,各种转录因子在调控网络中发挥着至关重要的作用。MADS-box转录因子不仅是植物生长发育各个环节的关键调控因子,同时也在植物响应逆境胁迫中发挥重要作用[28]。MADS-box转录因子在植物抗病过程中的作用越来越受到广大研究者的关注,Khong等[29]发现水稻OsMADS26基因负调控水稻抗寒性以及对稻瘟病、白叶枯病的抗性。Guo等[24]发现小麦与条锈菌亲和/非亲和互作过程中,I型MADS-box基因TaMADS2在受到条锈菌的侵染早期上调表达,在36和48 h分别出现下调表达,推测该基因可能参与调控小麦的条锈菌抗性。Ma等[23]通过六倍体小麦的全基因组表达谱分析发现MADS-box家族多种转录因子在赤霉菌、条锈菌和白粉菌等病原菌诱导下差异表达。本研究前期发现叶锈菌与亲和/非亲和小麦早期互作过程中,TaMADS2的表达量均受到叶锈菌诱导上调表达,且在非亲和小麦中TaMADS2的响应时期较亲和小麦更早。本研究选取叶锈菌小种Pt15的非亲和品系‘郑麦9023’以及亲和品系‘中国春’,利用BSMV-VIGS技术分别沉默TaMADS2基因表达,解析TaMADS2基因在小麦抗叶锈病中的功能。在接种病毒14 d后检测TaMADS2基因的表达量,感染BSMV:TaMADS2的小麦中TaMADS2基因的表达水平显著降低,TaMADS2基因沉默株系叶片上的叶锈菌孢子堆数量增多,分子病情指数增加。组织学观察和活性氧观察结果发现感染BSMV:TaMADS2的小麦叶片中病原菌生长速度更快,菌丝分枝数增多,活性氧积累量减少,说明沉默TaMADS2基因的表达降低了小麦对叶锈菌的抗性。
目前,病程相关蛋白(PRs)在植物抗病性以及系统获得抗性中的作用已有广泛报道,已经发现的PRs中PR1、PR2和PR5是植物抗病反应和SAR(Systemic acquired resistance)的标记基因[30]。PR1是在受到病原菌和SA大量诱导表达的PR蛋白[31],受侵染的组织中PR1的表达水平可以提高10 000倍,占整个叶片蛋白含量的1%~2%[32],因此被广泛认为是植物防御反应的标记基因。β-1,3-葡聚糖酶属于PR2家族成员,而β-1,3-葡聚糖是很多植物病原菌细胞壁的重要组成成分,β-1,3-葡聚糖酶能够催化β-1,3-葡聚糖多聚体的水解,从而抑制真菌的生长与增殖[33]。本研究中TaMADS2的沉默抑制了PR1和PR2基因的表达,表明TaMADS2可能通过病程相关蛋白途径抑制病原菌的生长,提高小麦对叶锈菌的抗性。
在早期的HR反应中病原菌的侵染会引起活性氧的爆发,活性氧可以直接作为抗菌物质发挥作用[34]。过氧化氢酶(CAT)是生物防御系统中的关键的活性氧清除酶类,能够清除超氧化物阴离子自由基,将H2O2降解为无毒的H2O和O2-,保护植物免受活性氧毒害,在植物防御、胁迫应答及控制细胞的氧化平衡中发挥重要的作用[35-36]。病程的发展伴随着ROS的大量产生,而活性氧清除酶基因CAT的表达量也会随之变化,常做为调控植物抵御病原菌的抗性参照指标[37]。本研究在叶锈菌侵染早期,TaMADS2沉默植株中活性氧积累量减少,CAT的表达量亦下降,表明TaMADS2基因可能通过调控侵染点活性氧的积累影响叶锈菌的侵染过程。本试验测定了侵染早期(12 h)TaCAT转录水平的相对变化,呈现下调趋势但未达显著差异水平,表明其受到了TaMADS2表达变化的影响,可能参与了这一抗性调控。但更准确描述CAT在TaMADS2调控小麦对叶锈菌抗性中作用,尚有待进一步的研究。
本研究利用BSMV-VIGS技术初步验证了TaMADS2基因在小麦抗叶锈病中的功能,叶锈菌侵染下TaMADS2基因抑制表达植株的叶锈菌孢子堆增多,病菌生物量增加,菌丝生长速度显著加快,活性氧积累量显著减少,病程相关基因PR1和PR2表达量显著减少,表明TaMADS2基因的沉默表达降低了小麦对叶锈菌的抗性。研究结果为进一步解析TaMADS2基因在小麦抗叶锈病中的作用机理奠定基础,也为抗病性小麦品种的遗传改良和利用提供了基因资源。
猜你喜欢
农业技术与装备(2021年12期)2021-12-02
湖北农业科学(2021年10期)2021-06-25
世界农药(2020年4期)2020-12-22
农业科技与信息(2020年20期)2020-12-18
现代园艺(2017年22期)2018-01-19
林业科学(2017年7期)2017-08-30
陕西林业科技(2016年1期)2016-08-04
山东医药(2015年38期)2015-12-07
中国学术期刊文摘(2015年2期)2015-10-27
食品工业科技(2014年13期)2014-03-11
