
侵染假酸浆的泰国番茄黄化曲叶病毒全基因组结构特征
2023-05-30 09:29张水英赵丽玲钟静李婷婷丁铭
植物保护 2023年2期
关键词:重组
张水英 赵丽玲 钟静 李婷婷 丁铭


摘要 为明确假酸浆Nicandra physalodes叶片黄化、皱缩症状是否由菜豆金色花叶病毒属病毒侵染引起,本研究利用分子检测方法和生物信息学技术鉴定了假酸浆样品中的病毒种类。从采集的病样中克隆并获得了2条菜豆金色花叶病毒属病毒DNA-A全序列和1条beta卫星全序列,经全序列分析发现,该双生病毒的两条DNA-A全序列与泰国番茄黄化曲叶病毒(tomato yellow leaf curl Thailand virus, TYLCTHV)云南分离物TYLCTHV-YN1732一致性最高,达99.3%,亲缘关系较近;beta卫星的全序列与云南番茄曲叶beta卫星(tomato leaf curl Yunnan betasatellite, TLCYnB)的分离物YN5230一致性最高,达99.3%,亲缘关系较近。重组分析显示,假酸浆上分离的TYLCTHV-YN5735-12是一个重组病毒,有两个重组事件,一个主要发生在AV1的编码区,由中国番茄黄化曲叶病毒(tomato yellow leaf curl China virus, TYLCCNV)和广西大戟曲叶病毒(euphorbia leaf curl Guangxi virus, EuLCGXV) 重组形成,另一个重组事件主要发生在基因间隔区(IR区),由泰国烟草卷叶病毒(tobacco leaf curl Thailand virus, TbLCTHV)和TYLCTHV重组形成。研究结果表明,假酸浆是TYLCTHV的新寄主,这是单组分菜豆金色花叶病毒属病毒及其伴随beta卫星侵染假酸浆的首次报道。
关键词 假酸浆; 泰国番茄黄化曲叶病毒; 基因组特征; 重组
中图分类号: S 432.1
文献标识码: A
DOI: 10.16688/j.zwbh.2021666
Abstract Molecular detection and bioinformatics analysis were performed to determine whether Nicandra physalodes plants in Yunnan province displaying yellow and crumple symptoms were associated with begomovirus infections. Two complete begomovirus DNA-A molecules and one betasatellite molecule were obtained from infected samples. Sequence comparison revealed that the two DNA-A sequences shared the highest identity (99.3%) with the tomato yellow leaf curl Thailand virus (TYLCTHV) isolate TYLCTHV-YN1732, while the betasatellite shared the highest sequence identity (99.3%) with the tomato leaf curl Yunnan betasatellite (TLCYnB) isolate YN5230. Recombination analysis revealed that the begomovirus infecting Nicandra physalodes was a recombinant virus derived from two potential recombinant events. One recombination event was detected in the coding region of AV1, contributed by tomato yellow leaf curl China virus (TYLCCNV) and euphorbia leaf curl Guangxi virus (EuLCGXV). The other recombination event occurred mainly in the intergenic region (IR), with the two parents being tobacco leaf curl Thailand virus (TbLCTHV) and TYLCTHV. This study recorded Nicandra physalodes as a new host for TYLCTHV. This is the first report of a monopartite begomovirus in association with a betasatellite infecting Nicandra physalodes.
Key words Nicandra physalodes; tomato yellow leaf curl Thailand virus; genome organization; recombination
煙粉虱Bemisia tabaci传播的菜豆金色花叶病毒属Begomovirus病毒是双生病毒科Geminiviridae中种类最多、分布最广、造成危害最严重的一类病毒[1]。按基因组结构分为双组分病毒和单组分病毒。双组分病毒由两条大小2.5~2.8 kb的DNA-A和DNA-B组成,单组分病毒只含有1条大小约为2.7 kb的基因组(DNA-A)。其全基因组含有6个开放阅读框,正义链上的AV1和AV2基因分别编码CP蛋白和Pre-CP蛋白,反义链上的AC1、AC2、AC3和AC4基因分别编码Rep、TrAP、REn和参与病毒系统运动的蛋白[2]。单组分病毒通常与beta卫星或与beta卫星和alpha卫星相伴随复合侵染植物[3-4]。
Beta卫星的基因组(DNAbeta)大小约为1.35 kb,是其辅助的病毒基因组大小的一半左右。DNAbeta保守区含有保守的9碱基序列“TAATATT↓AC”[5-6]。DNAbeta的一个开放阅读框编码βC1蛋白。βC1蛋白能够引起典型的病毒侵染症状,并能提高所辅助病毒在寄主植物中的积累量[7]。2020年Hu 等[8]研究发现,很多beta卫星的病毒链还可编码βV1蛋白,βV1蛋白有助于病毒的侵染。
泰国番茄黄化曲叶病毒(tomato yellow leaf curl Thailand virus, TYLCTHV)于1990年首次发现于泰国田间的番茄上[9]。其基因组由两条大小均约为2.8 kb的DNA-A和DNA-B组成,是一种双组分菜豆金色花叶病毒属病毒[10-11]。2004年Li等在云南从表现为卷叶的番茄上发现的TYLCTHV是单组分病毒,有3个样品还检测到DNAbeta序列[12]。2007年Jan等首次在台湾番茄上发现的TYLCTHV是双组分病毒,但并未发现有卫星DNA[13]。2009年Guo等在云南保山番茄上分离到的TYLCTHV是一种单组分病毒,并伴随有beta卫星[14]。2010年Shih等在台湾从表现为卷叶、小泡、畸形、叶脉黄化和萎缩的辣椒上首次发现的TYLCTHV是双组分病毒,但并未检测到DNAbeta[15]。
假酸浆 Nicandra physalodes (L.) Gaertner 又名大千生、田珠、冰粉、水晶凉粉、蓝花天仙子、鞭打绣球等,属茄科Solanaceae假酸浆属Nicandra。假酸浆原产于南美洲,在我国作为药用或观赏植物栽培,四川、云南、广西等地均有分布,生于田边、荒地、篱笆边、屋院周围,其全草可药用,有镇静、祛痰、清热解毒之效[16]。目前假酸浆上报道的病毒有番茄严重皱纹病毒(tomato severe rugose virus, ToSRV)[17]、马铃薯A病毒(potato virus A, PVA)[18]和苜蓿花叶病毒(alfalfa mosaic virus, AMV)[19],还未见其他病毒的报道。本研究采集了表现叶片黄化和皱缩的假酸浆病株,从中分离、鉴定出了菜豆金色花叶病毒属病毒,并对分离到的病毒核苷酸序列进行了比对和进化重组分析。
1 材料与方法
1.1 样品采集和DNA提取
2016年9月,在保山陇川县户撒阿昌族乡对田间双生病毒病害进行调查时发现,田间部分假酸浆植株表现出叶片黄化、皱缩等疑似双生病毒侵染引起的典型症状(图1),随机采集病株的叶片编号为YN5735,采用CTAB法[20]提取病株叶片的总DNA。
1.2 假酸浆病样的PCR检测
为了检测病叶中是否含有菜豆金色花叶病毒属病毒,分别利用该属病毒DNA-A基因組部分序列的通用简并引物BegoAForl/BegoARevl[21]以及DNA-B通用简并引物PCRc1/PBLv2040[22]对假酸浆总DNA进行PCR检测,同时利用beta和alpha卫星DNA通用引物β01/β02[23]和UNA101/UNA102[24](表1)分别检测是否含有病毒伴随的卫星DNA。
PCR反应体系:10×PCR反应缓冲液(含 Mg2+)2.5 μL,dNTPs(2.5 μmol/L)2 μL,上、下游引物(20 μmol/L)各0.5 μL,Taq Plus DNA 聚合酶(5 U/μL,上海申能博彩生物科技有限公司)0.5 μL,约100 ng总DNA,加双蒸水至25 μL。扩增条件:94℃预变性2 min;94℃变性45 s,50℃退火45 s,72℃延伸2 min,30个循环;最后72℃延伸10 min。PCR产物经1.0%琼脂糖凝胶电泳检测后割取目的片段,利用Axygen DNA凝胶回收试剂盒分离纯化,回收PCR产物连接至pGEM-T Easy载体,再转入大肠杆菌DH5α中,挑选阳性克隆,送上海立菲生物技术有限公司测序。
1.3 假酸浆中菜豆金色花叶病毒属病毒基因组克隆
为获得高纯度的菜豆金色花叶病毒属病毒及其卫星的基因组,采用Templiphi TM Kit(GE Healthcare)对假酸浆病样总DNA进行了滚环扩增(rolling circle amplification,RCA)。具体步骤为:将1 μL植物总DNA样品(约50 ng)加入到5 μL样品缓冲液中,均匀混合后95℃变性3 min,立刻转至冰上放置10 min;然后加入5 μL反应缓冲液和0.2 μL酶(phi29 DNA聚合酶),30℃反应18 h;最后65℃加热10 min终止反应。
根据测序后获得的部分DNA-A序列,设计全长引物TYLCTHV-BeF/TYLCTHV-BeR(表1)扩增该病毒DNA-A基因组全长序列。PCR反应体系:2×PCR反应缓冲液(含Mg2+)25 μL,dNTPs(2 μmol/L)10 μL,上、下游引物(20 μmol/L)各1 μL,KOD FX DNA聚合酶(1 U/μL,日本东洋纺)2 μL,RCA产物0.8 μL,加双蒸水至50 μL。扩增条件:94℃预变性2 min;94℃变性45 s,62℃退火45 s,72℃延伸3 min,30个循环;72℃延伸10 min。按 1.2 中的方法进行克隆测序。
1.4 假酸浆中病毒分离物基因组序列分析
采用DNASTAR Lasergene 7.1 对假酸浆中分离到的DNA-A及卫星DNA的核苷酸序列进行病毒基因组全序列拼接,利用DNAMAN 5.22 (Lynnon Biosoft, Quebec, Canada)和DNASTAR Lasergene 7.1 确定开放阅读框(ORF)。获得的全基因组序列采用BLAST(https:∥blast.ncbi.nlm.nih.gov/Blast)进行核苷酸序列比较,选择亲缘关系较近、同属代表种序列,利用SDT软件进一步进行比较分析,根据SDT比较结果确定所分离病毒的分类地位[25],运用MEGA 6软件中的Clustal W方法进行深入比对。采用MEGA 6软件中的邻近法(neighbor-joining)构建进化树,可信度使用1 000次自导复制验证[26]。采用RDP 4.46软件的默认设置进行序列的重组分析[27]。
2 结果与分析
2.1 假酸浆中菜豆金色花叶病毒属病毒PCR检测
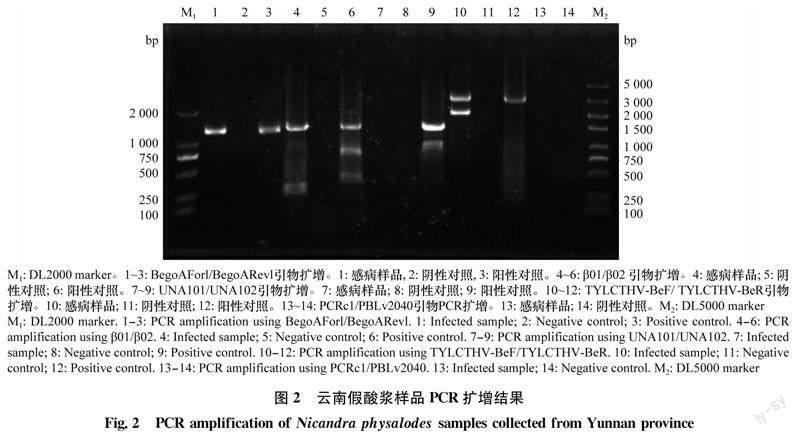
利用DNA-B的简并引物PCRc1/PBLv2040对YN5735样品进行PCR扩增,样品中未扩增到相应条带。以BegoAForl/BegoARevl为引物从YN5735样品中扩增到约1 200 bp的目的片段(图2),将获得的目的条带进行克隆,并选取2个阳性克隆(YN5735-1、YN5735-6)测序。结果显示:分离物序列与泰国番茄黄化曲叶病毒分离物的序列一致性高达99.2%,推测YN5735样品被菜豆金色花叶病毒属病毒侵染。
2.2 假酸浆中菜豆金色花叶病毒属病毒全基因组结构分析
2.2.1 TYLCTHV的鉴定及基因组结构
以YN5735样品的DNA为模板进行RCA扩增,再以RCA扩增产物作为模板,利用TYLCTHV-BeF/TYLCTHV-BeR扩增病毒全长序列,筛选获得2个阳性克隆YN5735-12和YN5735-15,经测序、分析发现2个克隆的基因组序列一致性为100%,选择YN5735-12进行进一步分析。
基因组结构分析显示:TYLCTHV-YN5735-12(GenBank 登录号:MF040760)全长2 738 bp,其基因组结构具有典型的单组分双生病毒结构,编码6个开放阅读框,其AV1(293—1 063 nt)、AV2(133—471 nt)、AC1(1 512—2 597 nt)、AC2(1 205—1 609 nt)、AC3(1 060—1 464 nt)和AC4(2 147—2 446 nt),其基因间隔区(intergenic region,IR)包含273 bp(2 598—132 nt),在该区域内含有菜豆金色花叶病毒属病毒复制和转录所必需的相关结构。
2.2.2 TYLCTHV全基因组相似性比较
运用SDT软件进行相似性分析,发现TYLCTHV-YN5735-12分离物的DNA-A全长序列与参与比对的TYLCTHV分离物的一致性均在91.0%以上,并且与云南分离物TYLCTHV-YN1732(登录号:KX290922)的一致性达到99.3%(表2)。
根据菜豆金色花叶病毒属病毒的分类标准:DNA-A的核苷酸一致性大于91.0%则认为是同种病毒,同种成员间的核苷酸一致性小于94.0%则表示这些成员属于不同的株系,大于94.0%则属于同一株系[28]。因此,假酸浆样品中分离到的菜豆金色花叶病毒属病毒是泰国番茄黄化曲叶病毒的一个分离物(TYLCTHV-YN5735-12)。与TYLCTHV-YN1732為同一株系。运用MegAlign软件进一步分析TYLCTHV-YN5735-12分离物DNA-A序列与其他菜豆金色花叶病毒属病毒的相似性,发现TYLCTHV-YN5735-12的编码区和非编码区,除AV2的氨基酸序列外,IR区、AV1、AC1、AC2、AC3和AC4的核苷酸和氨基酸序列均与TYLCTHV-YN1732的一致性最高,达97.1%~100.0%(表2)。
2.2.3 TYLCTHV全基因组进化与重组分析
选择与TYLCTHV-YN5735-12一致性较高的菜豆金色花叶病毒属病毒序列进行系统进化分析,发现所有TYLCTHV的DNA-A分离物都聚为1个大分支,TYLCTHV-YN5735-12与云南分离物TYLCTHV-YN1732聚为一个小分支,与缅甸分离物TYLCTHV-TLCV-MM聚于一个稍大的分支,表明本研究分离到的分离物与TYLCTHV-YN1732和TYLCTHV-TLCV-MM亲缘关系较近(图3)。
重组分析显示,RDP、GENECONV、BootScan、MaxChi、Chimaera、SiScan和3Seq 7种分析方法都能检测到两个重组事件发生,一个重组事件主要发生在AV1编码区,起始位置为510 nt,结束位置为1 056 nt,其主要亲本为TYLCCNV-Y11,次要亲本为EuLCGXV-G35-1(RDP 值:3.44×10-15;GENECONV值:2.754×10-8;BootScan值:4.687×10-14;MaxChi值:6.055×10-12;Chimaera值:3.162×10-11;SiScan值:9.520×10-11;3Seq值:1.414×10-13)。另一个重组事件主要发生在基因间隔区(IR区),起始位置为2 704 nt,结束位置为58 nt,其主要亲本为 TbLCTHV-KKN-13G,次要亲本为TYLCTHV-WF-SPN-Tom2015(RDP 值:2.289×10-12;GENECONV值:3.765×10-14;BootScan值:1.430×10-5;MaxChi值:3.157×10-5;Chimaera值:1.258×10-3;SiScan值:2.762×10-10;3Seq值:2.602×10-7)。根据RDP分析方法的判断标准,为了保证RDP分析结果的可靠性,每个方法所得P值需小于0.05且至少有3个方法都检测到同一重组事件才认为有重组事件发生[27,29],由此可见,TYLCTHV-YN5735-12是一个重组病毒。
2.3 病毒伴随的卫星DNA分子结构及其进化重组分析
alpha卫星DNA通用引物UNA101/UNA102(表1)未能从YN5735样品中扩增到相应条带。利用beta卫星DNA通用引物β01/β02(表1)扩增到约1 300 bp的目的片段。经克隆测序后得到一条beta卫星序列(YN5735-42),其具有典型的单组分病毒伴随性beta卫星的基因组结构:在反义链的212—592 nt位置编码一个βC1蛋白,包含一个卫星保守区(satellite conserved region,SCR),位于1 221—1 360 nt,1—10 nt,一个A富含区(A-rich),位于891—977 nt,A含量达67.8%。
经SDT比对,YN5735-42与云南番茄曲叶beta卫星分离物TLCYnB-YN5230(登录号:MF461727)一致性最高,达99.3%,与其他云南番茄曲叶beta卫星分离物的一致性在92.3%以上,与其他病毒beta卫星分离物的一致性在78.0%以下(表3)。根据ICTV beta卫星分子的分类标准:全序列核苷酸一致性小于78.0%则认为是新beta卫星,大于78.0%则认为属于同一种beta卫星[30]。上述结果表明假酸浆上分离得到的beta卫星DNA属于TLCYnB,命名为TLCYnB-YN5735-42。
系统进化树分析表明,YN5735-42与TLCYnB的其他几个分离物均聚在一个较小的分支上,亲缘关系较近,且这些beta卫星DNA都是从云南分离获得的(图4),具有明显的地理分布特征。表明保山陇川假酸浆上分离得到的TYLCTHV有beta卫星伴随。
重组分析发现,TLCYnB-YN5735-42有两个重组事件,一个重组事件的起始位置为418 nt,结束位置为558 nt,其主要亲本为CLCuMuB-BH12,次要亲本为CLCuBuB-LG-RCA-bI-r(GENECONV值:2.904×10-8;MaxChi值:6.289×10-3;SiScan值:2.292×10-19)。另一个重组事件的起始位置为906 nt,结束位置为1 322 nt,其主要亲本为TYLCCNB-G32,次要亲本为CLCuBuB-C-32(RDP 值:4.300×10-11;MaxChi值:2.582×10-5;Chimaera值:3.687×10-5;SiScan值:9.420×10-18;3Seq值:1.239×10-4)。根據RDP分析方法的判断标准[27],TLCYnB-YN5735-42是一个重组beta卫星。
3 结论与讨论
本研究从表现黄化、皱缩的假酸浆样品中分离到泰国番茄黄化曲叶病毒,利用分子检测方法和生物信息学技术对该病毒进行分析,结果表明,该病毒与云南番茄曲叶beta卫星相伴随,这是假酸浆上泰国番茄黄化曲叶病毒伴随云南番茄曲叶beta卫星的首次报道。
TYLCTHV刚发现时被认为是一种双组分病毒[10-11],后来发现,TYLCTHV既是一种单组分病毒,也是一种双组分病毒,单组分TYLCTHV主要在中国云南[12,14]和缅甸的番茄上发现[31-32],双组分TYLCTHV主要在泰国、中国南部[33]、越南、柬埔寨和中国台湾[13]一些地区的番茄、辣椒、甜椒[15]和烟粉虱[34]上发现。在研究双组分病毒斯里兰卡木薯花叶病毒(Sri Lankan cassava mosaic virus, SLCMV)时,发现其DNA-A具有单组分begomovirus病毒的生物学特征,其DNA-B可能是从与其共同侵染木薯的印度木薯花叶病毒(Indian cassava mosaic virus, ICMV)中获得的DNA-B组分进化而来,因此SLCMV被认为是从单组分到双组分病毒进化的中间体[35-36]。综上所述,一些单组分双生病毒在田间具有捕获DNA-B组分的能力,从而变成双组分双生病毒。2009年Guo等[14]研究表明,泰国的TYLCTHV DNAbeta是由单组分病毒从另一个双组分begomovirus病毒中获取DNA-B进化而来的。TYLCTHV可能代表了介于多数类型和少数类型之间的进化中间体[14]。
突变和重组是菜豆金色花叶病毒属病毒进化的主要动力,重组能够导致菜豆金色花叶病毒属病毒产生新种和新分离物[29]。菜豆金色花叶病毒属病毒复合侵染容易造成该属病毒之间重组,从而可能扩大寄主范围,造成更加严重的危害[37]。之前对双生病毒重组热点的研究发现,病毒重组通常发生于IR区,但也有报道CP、AV2、AC1、AC4和BC1基因也存在潜在的重组位点[38-39]。本研究发现的TYLCTHV-YN5735-12分离物有两个重组区域,一个主要发生在IR区(2 704—58 nt),其主要亲本为在泰国辣椒上分离得到的TbLCTHV-KKN-13G,次要亲本为在泰国烟粉虱上分离得到的TYLCTHV-WF-SPN-Tom2015,说明本研究分离得到的TYLCTHV很有可能是由泰国的烟粉虱传过来的。另一个重组事件主要发生在AV1编码区(510—1 056 nt),其主要亲本为中国云南烟草上分离得到的TYLCCNV-Y11,次要亲本为中国广西一品红上分离得到的EuLCGXV-G35-1,由此可以看出,本研究分离得到的TYLCTHV-YN5735-12的重组亲本可能与地理位置有一定的关系。本研究获得的TLCYnB-YN5735-42分离物有两个重组事件,一个位于βC1的编码区(418—558 nt),其主要亲本为在巴基斯坦旁遮普棉花上分离得到的CLCuMuB-BH12,次要亲本为在印度旁遮普棉花上分离到的CLCuBuB-LG-RCA-bI-r;另一个位于SCR区及A-rich区(906—1 322 nt),其主要亲本为在中国广西番茄上分离到的TYLCCNB-G32,次要亲本为在巴基斯坦费萨拉巴德棉花上分离到的CLCuBuB-C-32。
目前泰国番茄黄化曲叶病毒的寄主主要有番茄、辣椒、甜椒和烟粉虱,暂还未有其他寄主的报道。目前假酸浆上分离的病毒很少,尤其是双生病毒只有番茄严重皱纹病毒,此病毒是一种双组分双生病毒[18],本研究发现的TYLCTHV是一种单组分病毒,说明单组分和双组分菜豆金色花叶病毒属病毒均可侵染假酸浆。因此我们还应该扩大双生病毒在观赏植物和药用植物上的监测,这样才能更有效地预防双生病毒病的危害。
参考文献
[1] VARMA A, MALATHI V G. Emerging geminivirus problems: a serious threat to crop production [J]. Annals of Applied Biology, 2015, 142(2): 145-164.
[2] 楊秀玲.中国番茄黄曲叶病毒抵御RNA沉默的机制和番茄黄曲叶病毒的分子变异研究[D]. 杭州: 浙江大学, 2011.
[3] ZHAO Liling, ZHONG Jing, ZHANG Xiaoyun, et al. Two distinct begomoviruses associated with an alphasatellite coinfecting Emilia sonchifolia in Thailand [J]. Archives of Virology, 2018, 163(6):1695-1699.
[4] TAHIR M N, HAMEED A, AMIN I, et al. Characterization of a begomovirus-betasatellite complex, producing defective molecules in spinach (Spinacia Oleracea L.), a new host for begomovirus and betasatellite complex in Pakistan [J]. The Plant Pathology Journal, 2017, 33(5): 514-521.
[5] CUI Xiaofeng, TAO Xiaorong, XIE Yan, et al. A DNAβ associated with tomato yellow leaf curl China virus is required for symptom induction [J]. Journal of Virology, 2004, 78(24): 13966-13974.
[6] ZHOU Xueping, XIE Yan, TAO Xiaorong, et al. Characterization of DNAβ associated with begomoviruses in China and evidence for co-evolution with their cognate viral DNA-A [J]. Journal of General Virology, 2003, 84(1): 237-247.
[7] TAHIR M N, MANSOOR S. βC1 of chili leaf curl betasatellite is a pathogenicity determinant [J/OL]. Virology Journal, 2011, 8(1): 509. DOI: 10.1186/1743-422X-8-509.
[8] HU Tao, SONG Yu, WANG Yaqing, et al. Functional analysis of a novel βV1 gene identified in a geminivirus betasatellite [J]. Science China Life Sciences, 2020, 63(5): 688-696.
[9] ATTATHOM S, CHIEMSOMBAT P, SUTABUTRA T, et al. Characterization of nucleic acid of tomato yellow leaf curl virus [J]. Kasetsart Journal, Natural Sciences, 1990, 24 (5): 1-5.
[10]ATTATHOM S, CHIEMSOMBAT P, KOSITRATANA W, et al. Complete nucleotide sequence and genome analysis of bipartite tomato yellow leaf curl virus in Thailand [J]. Kasetsart Journal, Natural Sciences, 1994, 28(4): 632-639.
[11]ROCHESTER D E, DEPAULO J J, FAUQUET C M, et al. Complete nucleotide sequence of the geminivirus tomato yellow leaf curl virus, Thailand isolate [J]. Journal of General Virology, 1994, 75(3): 477-485.
[12]LI Zhenghe, ZHOU Xueping, ZHANG Xiongwen, et al. Molecular characterization of tomato-infecting begomoviruses in Yunnan, China [J]. Archives of Virology, 2004, 149(9): 1721-1732.
[13]JAN F J, GREEN S K, SHIH S L, et al. First report of tomato yellow leaf curl Thailand virus in Taiwan [J]. Plant Disease, 2007, 91(10): 1363.
[14]GUO Wei, YANG Xiuling, XIE Yan, et al. Tomato yellow leaf curl Thailand virus-[Y72]from Yunnan is a monopartite begomovirus associated with DNAbeta [J]. Virus Genes, 2009, 38(2): 328-333.
[15]SHIH S L, TSAI W S, LEE L M, et al. First report of tomato yellow leaf curl Thailand virus associated with pepper leaf curl disease in Taiwan [J]. Plant Disease, 2010, 94(5): 637.
[16]中國科学院中国植物志编辑委员会. 中国植物志: 第67卷第一分册[M]. 北京: 科学出版社, 1978: 6-8.
[17]BARBOSA J C, BARRETO S S, INOUE-NAGATA A K, et al. Natural infection of Nicandra physaloides by tomato severe rugose virus in Brazil [J]. Journal of General Plant Pathology, 2009, 75(6): 440-443.
[18]RAJAMAEKI M L, VALKONEN J. The 6K2 protein and the VPg of potato virus A are determinants of systemic infection in Nicandra physaloides [J]. Molecular Plant Microbe Interaction, 1999, 12(12): 1074-1081.
[19]GALLO J, CIAMPOR F. Transmission of alfalfa mosaic virus through Nicandra physaloides seeds and its localization in embryo cotyledons [J]. Acta Virologica, 1977, 21(4): 344-346.
[20]DOYLE J J, DOYLE J L, DOYLE J A, et al. A rapid DNA isolation procedure for small quantities of fresh leaf tissue [J]. Phytochemical Bulletin, 1987, 19(1): 11-15.
[21]HA C, COOMBS S, REVILL P, et al. Corchorus yellow vein virus, a New World geminivirus from the Old World [J]. Journal of General Virology, 2006, 87(4): 997-1003.
[22]ROJAS M R, GILBERTSON G L, RUSSELL D R, et al. Use of degenerate primers in the polymerase chain reaction to detect whitefly-transmitted geminiviruses [J]. Plant Disease, 1993, 77(4): 340-347.
[23]BRIDDON R W, BULL S E, MANSOOR S, et al. Universal primers for the PCR-mediated amplification of DNAβ a molecule associated with some monopartite begomoviruses [J]. Molecular Biotechnology, 2002, 20(3): 315-318.
[24]BULL S E, BRIDDON R W, MARKHAM P G. Universal primers for the PCR-mediated amplification of DNA 1: a satellite-like molecule associated with begomovirus-DNAβ complexes [J]. Molecular Biotechnology, 2003, 23(1): 83-86.
[25]MUHIRE B M, VARSANI A, MARTIN D P. SDT: a virus classification tool based on pairwise sequence alignment and identity calculation [J/OL]. PLoS ONE, 2014, 9(9): e108277. DOI:10.1371/journal.pone.0108277.
[26]TAMURA K, STECHER G, PETERSON D, et al. MEGA 6: molecular evolutionary genetics analysis version 6.0[J]. Molecular Biology and Evolution, 2013, 30(12): 2725-2729.
[27]MARTIN D P, LEMEY P, LOTT M, et al. RDP 3: a flexible and fast computer program for analyzing recombination [J]. Bioinformatics, 2010, 26(19): 2462-2463.
[28]BROWN J K, ZERBINI F M, NAVAS-CASTILLO J, et al. Revision of begomovirus taxonomy based on pairwise sequence comparisons [J]. Archives of Virology, 2015, 160(6): 1593-1619.
[29]赵丽玲, 施章吉, 李婷婷, 等. 野茼蒿黄脉病毒的田间寄主范围及其基因多样性特征[J]. 植物保护学报, 2020, 47(3): 647-656.
[30]BROWN J K, FAUQUET C M, BRIDDON R W, et al. Virus taxonomy: Ninth report of the international committee on taxonomy of viruses [M]. San Diego: Elsevier Academic Press, 2012: 351-373.
[31]GREEN S K, TSAI W S, SHIH S L, et al. Molecular characterization of begomoviruses associated with leaf curl diseases of tomato in Bangladesh, Laos, Malaysia, Myanmar, and Vietnam [J]. Plant Disease, 2001, 85(12): 1286.
[32]KENYON L, TSAI W S, SHIH S L, et al. Emergence and diversity of begomoviruses infecting solanaceous crops in East and Southeast Asia [J]. Virus Research, 2014, 186: 104-113.
[33]SEEPIBAN C, CHAROENVILAISIRI S, WARIN N. Development and application of triple antibody sandwich enzyme-linked immunosorbent assays for begomovirus detection using monoclonal antibodies against tomato yellow leaf curl Thailand virus [J/OL]. Virology Journal, 2017, 14(1): 99. DOI: 10.1186/s12985-017-0763-2.
[34]YULE S, CHIEMSOMBAT P, SRINIVASAN R. Detection of tomato yellow leaf curl Thailand virus transmitted by Bemisia tabaci Asia I in tomato and pepper [J]. Phytoparasitica, 2019, 47(5): 143-153.
[35]BLAWID R, VAN D T, MAISS E. Transreplication of a tomato yellow leaf curl Thailand virus DNA-B and replication of a DNA component by tomato leaf curl Vietnam virus and tomato yellow leaf curl Vietnam virus [J]. Virus Research, 2008, 136(1/2): 107-117.
[36]SAUNDERS K, SALIM N, MALI V R, et al. Characterisation of Sri Lankan cassava mosaic virus and Indian cassava mosaic virus: evidence for acquisition of a DNA B component by a monopartite begomovirus [J]. Virology, 2002, 293(1): 63-74.
[37]趙丽玲, 钟静, 施章吉, 等. 复合侵染水茄的两种菜豆金色花叶病毒属病毒基因组结构特征分析[J]. 植物保护学报, 2020, 47(2): 355-364.
[38]郭维. 三种双生病毒及其伴随卫星DNA的致病性研究[D]. 杭州: 浙江大学, 2008.
[39]钟静, 赵丽玲, 尹跃艳, 等. 一种侵染鳢肠的双生病毒基因组特征[J]. 植物病理学报, 2017, 47(4): 479-486.
(责任编辑:杨明丽)
猜你喜欢
文艺生活·下旬刊(2016年12期)2017-01-11
北方文学·下旬(2016年9期)2016-12-20
智富时代(2016年12期)2016-12-01
智富时代(2016年12期)2016-12-01
中国市场(2016年22期)2016-07-07
商(2016年16期)2016-06-12
商(2016年14期)2016-05-30
