
miR-361-3P调控转化生长因子β1诱导成骨细胞凋亡的研究
2023-08-21 01:36王帆王鹏皓
中国医科大学学报 2023年7期
王帆,王鹏皓
(中国医科大学附属第一医院骨科,沈阳 110001)
人体骨骼的正常代谢依靠成骨细胞与破骨细胞的动态平衡维持。这种动态平衡受骨组织周围微环境中存在的多种细胞因子调节,其中,转化生长因子β1 (transforming growth factor-β1,TGF-β1) 作为骨组织局部微环境中至关重要的调节因子,在成骨活动中发挥十分重要的作用[1]。细胞凋亡使细胞在基因控制下程序性死亡,进而维持体内细胞数量动态平衡,影响组织周围内环境相对稳定。研究[2]证明,微RNA (micro RNA,miRNA) 对成骨细胞的再生、转化、分化等代谢过程起至关重要的作用。目前对miR-361-3P在骨代谢相关疾病中的研究较少见,且其对成骨细胞自我程序性死亡的作用机制尚未清楚。因此,本研究拟通过分子生物学技术检测人成骨细胞内miR-361-3P对TGF-β1和细胞凋亡标志分子caspase 3蛋白活性表达的影响,以探讨miR-361-3P/TGF-β1通路在人成骨细胞凋亡中的作用机制。
1 材料与方法
1.1 细胞培养
用DMEM培养液 (含10 % 胎牛血清、300 mg/L G418) 于37 ℃、5% CO2培养箱中培养人成骨细胞系hFob1.19 (北京307-青藤转化医学中心)。
1.2 细胞分组及转染
miR-361-3p mimic (mimic)、miR-361-3p inhibitor(inhibitor) 和 TGF-β1 siRNA (si-TGF-β1) 均购自上海吉凯基因医学科技股份有限公司。将hFob1.19细胞随机分为control组、mimic组及其阴性对照组 (mimic-NC组)、inhibitor 组及其阴性对照组 (inhibitor-NC组)、si-TGF-β1及其阴性对照组 (si-NC组)。采用Lipofectamine 2000脂质体转染系统 (北京中杉金桥生物有限公司),于37 ℃、5% CO2、95%湿度条件下行细胞转染8 h。
1.3 CCK-8实验
将hFob1.19细胞转移至6孔板,培养48 h,每孔加入10 μL CCK-8 (5 mg/mL,北京中杉金桥生物有限公司),孵育2 h,450 nm 处检测吸光度。
1.4 实时PCR (real-time quantitative PCR,RT-qPCR)
用TRIzol试剂提取细胞总RNA。采用Nanodrop 2000测定RNA样品的浓度和纯度。用primematipt混合RT 随机引物对mRNA行逆转录。用TRIzol reagent行miRNA逆转录。采用PrimeScript RT Master Mix及ABI Prism7500序列检测系统进行cDNA扩增。以GAPDH和U6作内参照。采用2-ΔΔCt法分析miRNA和mRNA的相对表达量。采用不同引物检测环状RNA的后剪接,采用聚合引物检测线性mRNA。miRNA和mRNA引物见表1。
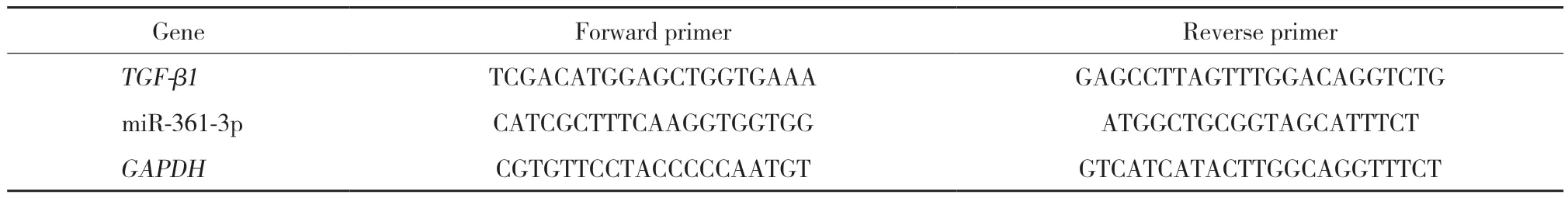
表1 miRNA和mRNA引物设计 (5’-3’)Tab.1 Primer design for the miRNA and mRNA expression assays (5’-3’)
1.5 Western blotting
采用放射免疫沉淀法提取细胞总蛋白,用8%或10% SDS-PAGE分离后,转移至PVDF膜。加入TGF-β1一抗(稀释1 ∶250,美国赛默飞公司),37 ℃孵育过夜。用PBS洗膜后,加入过氧化物酶偶联的二抗 (美国赛默飞公司),37 ℃孵育2 h。PBS洗膜,采用增强化学发光法显色。用GAPDH (稀释1 ∶2 500) 作内参照。
1.6 报告基因荧光素酶活性测定
将miR-361-3p和TGF-β1的3’-UTR插入pmirGLO质粒,获得荧光素酶报告载体 (pmirGLO-miR-361-3p-wt,pmirGLO,TGF-β1-wt)。构建突变质粒 (pmirGLOmiR-361-3p-mut和pmirGLO-TGF-β1-mut)。96孔板中培养hFob1.19细胞24 h,分别转染上述pmirGLO质粒48 h,使用双荧光素酶报告基因检测系统 (美国Promega公司) 测定荧光素酶活性。
1.7 统计学分析
采用Prism7.0软件进行统计学分析。计量资料以±s表示。采用Shapiro-Wilk检验分析数据分布,Levene检验分析方差齐。采用未配对的Student’st检验 (正态分布和方差齐数据)、Welcht检验 (方差不齐数据) 或Mann-WhitneyU检验 (非正态分布数据) 进行组间比较。P< 0.05为差异有统计学意义。
2 结果
2.1 miR-361-3p对成骨细胞内TGF-β1的调节作用
miR-361-3p mimic作用成骨细胞48 h后,细胞内miR-361-3p mRNA表达水平明显升高 (图1A),TGF-β1mRNA表达水平显著下调,TGF-β1蛋白表达水平受到明显抑制 (图1B、1C),差异均有统计学意义 (P< 0.05)。

图1 miR-361-3p 对成骨细胞内TGF-β1表达的影响Fig.1 Effects of miR-361-3p on TGF-β1 expression in osteoblasts
2.2 miR-361-3p对成骨细胞增殖及凋亡的影响
miR-361-3p mimic作用成骨细胞48 h后,CCK-8结果显示细胞增殖被显著抑制 (图2A);同时,酶联免疫吸附实验和Western blotting均发现,成骨细胞内caspase 3蛋白表达水平明显升高 (图 2B、2C),差异均有统计学意义 (均P< 0.05)。

图2 miR-361-3p对成骨细胞增殖及caspase 3 活性的调控作用Fig.2 Regulation of miR-361-3p on osteoblast proliferation and caspase 3 activity
2.3 miR-361-3p负向调控靶基因TGF-β1对成骨细胞增殖和凋亡的影响
miR-361-3p与TGF-β1mRNA的3’-UTR区域存在多个结合位点 (图3A)。荧光素酶报告基因实验显示,miR-361-3p mimic组中 WT-TGF-β1的荧光素酶活性明显降低 (图3B),证实miR-361-3p可与TGF-β1mRNA结合并干预其表达;与control组和inhibitor-NC组比较,miR-361-3p inhibitor可明显抑制成骨细胞内miR-361-3p mRNA的表达和caspase 3的蛋白活性 (图 3C、3F),显著增强成骨细胞增殖效应 (图3E);si-TGF-β1可显著抑制成骨细胞增殖,而显著增强caspase 3的蛋白活性 (图3D、3F),差异均有统计学意义 (均P<0.05)。以上均证实miR-361-3p/TGF-β1对成骨细胞增殖和凋亡具有调控作用。

图3 miR-361-3p/TGF-β1调控成骨细胞增殖和凋亡的作用Fig.3 Role of miR-361-3p/TGF-β1 in regulating osteoblast proliferation and apoptosis
3 讨论
miRNA是基因表达调控的重要组成部分,通常由20~24个核苷酸组成,通过多种途径 (包括直接抑制翻译过程,或通过烯基化使mRNA靶点降解) 参与细胞增殖、凋亡、迁移等过程及炎症、感染、固有免疫的调控,在机体的免疫应答、肿瘤发生发展等方面发挥重要作用[3]。近年来,miRNA对骨代谢的影响受到广泛关注。研究表明,miR-150-3p 通过β-catenin对间充质干细胞的成骨分化产生影响[4];miR-23b通过影响Smad3抑制MC3T3-E1细胞的成骨化[5];miR-103a通过在转录过程后调控Runx2,抑制成骨细胞的活性,从而降低细胞外基质的矿化[6];miR-204和miR-211通过抑制Runx2蛋白在成骨活动中的负反馈调节,增加人骨髓间充质干细胞内的脂肪生成[7]。本研究结果显示,miR-361-3p可能通过抑制靶基因TGF-β1的转录和蛋白活性,抑制成骨细胞的增殖,并促进成骨细胞凋亡。
miRNA通过与靶基因相关序列位点结合发挥作用。此序列通常仅包含6~8个碱基,因此,1个miRNA可能对多个mRNA产生作用,而1个 mRNA可能受多个miRNA调控[8]。某些miRNA的缺失可能不会影响机体正常的生长发育,但可能引发疾病,很多慢性疾病的发生均与miRNA特殊的作用方式有关。miR-361-3p作为一种抗炎mi-RNA,可通过靶向抑制活化性T淋巴细胞核因子5调节核因子κB[9];miR-361-3p在肺组织中存在表达,通过抑制癌细胞的生成、转移,加速癌细胞的凋亡,发挥抗癌的功能[10];miR-361-3p过量表达可抑制高糖环境下的肾小管上皮细胞凋亡,降低乳酸脱氢酶含量,提高肾小管上皮细胞存活率,降低细胞内丙二醛和caspase 3 的表达水平[11]。miR-361-3p能调控多种靶基因,如胰岛素样生长因子1、TNF、p73和促卵泡激素等[12]。本研究表明,人成骨细胞内的miR-361-3P与靶基因TGF-β1的启动区有8个碱基结合位点,可靶向抑制TGF-β1基因转录,参与成骨细胞凋亡。
TGF-β1具有多效性及多向性,通过自分泌或旁分泌等方式与细胞表面受体信号结合,调节细胞增殖、分化及凋亡。研究[13]表明,TGF-β1对心肌细胞增殖、凋亡及成纤维化等有调控作用;TGF-β1高表达通过旁分泌机制使心脏成纤维细胞的胶原形成增加,导致心肌肥厚、心肌收缩功能降低[14]。骨组织微环境中,TGF-β1由成骨细胞产生,通过控制成骨细胞调节骨代谢活动,在骨组织形成和骨组织吸收的动态平衡中发挥至关重要的作用。对小鼠成骨细胞的研究发现,TGF-β1可通过降低RANKL、增加OPG的表达,减少破骨细胞的生成和骨吸收降低[15]。本研究通过检测凋亡蛋白caspase 3的活性表达,探讨miR-361-3p/TGF-β1和成骨细胞凋亡间可能存在的相关性。结果发现,当阻断miR-361-3p/TGF-β1通路时,成骨细胞凋亡和caspase 3蛋白活性明显增强,表明miR-361-3p/TGF-β1可负向调控成骨细胞的凋亡。
综上所述,本研究探讨了miR-361-3p通过TGF-β1影响成骨细胞增殖及凋亡的作用机制,发现TGF-β1是miR-361-3p的靶基因,且miR-361-3p通过下调TGF-β1的表达,促进成骨细胞增殖,抑制成骨细胞凋亡。
猜你喜欢
天津医科大学学报(2021年4期)2021-08-21
中日友好医院学报(2021年1期)2021-04-14
山东医药(2020年9期)2020-05-20
山东陶瓷(2019年2期)2019-02-17
实用口腔医学杂志(2017年6期)2017-09-19
安徽医科大学学报(2016年12期)2017-01-15
中国现代医生(2016年11期)2016-09-06
中国民族医药杂志(2016年6期)2016-05-09
吉林大学学报(医学版)(2015年5期)2015-12-16
中国医科大学学报(2015年10期)2015-03-01
