
基于高通量测序技术解析中高温制曲真菌群落的演替规律研究
2023-11-23 11:09葛向阳张聪芝李燕荣贾亚伟
酿酒科技 2023年10期
杨 勇,葛向阳,2*,张聪芝,李燕荣,贾亚伟
(1.江苏洋河酒厂股份有限公司,江苏宿迁 223800;2.宿迁学院,江苏宿迁 223800)
大曲作为一种富含多酶多菌的微生态制品,在 中国白酒的酿造过程中起着糖化、生香、发酵、提供菌源的作用,是传统固态发酵蒸馏大曲酒的重要物质保障,被人们评价“曲为酒之骨”[1]。对大曲中真菌微生物的研究最早是通过传统的分离培养方法,葛向阳等[2]经过多级富集、分离、纯化与鉴定,从洋河传统包包曲中分离得到毕赤酵母属、异常汉逊酵母、假丝酵母等酵母菌类群以及卷枝毛霉、寄生曲霉、黑曲霉和黄曲霉等霉菌类群;罗惠波等[3]发现泸州老窖大曲内存在大量的曲霉属、根毛霉属、横梗霉属等霉菌类群以及酿酒酵母、德巴利氏酵母属、异常威克汉姆酵母、锁掷酵母属等酵母菌类群。对中高温制曲发酵过程中可培养微生物的消长及生化指标的变化规律也做了相关研究[4-5],然而,传统的分离培养方法受到培养基的选择性、微生物的生长特性等因素影响[6],很难直接展现大曲中微生物的真实群落结构。
随着分子生物学的发展,基于免培养的高通量测序技术已经越来越广泛地应用于微生物群落的研究。谭崇尧等[7]利用高通量测序发现北方派浓香型白酒大曲优势真菌有热子囊菌属、嗜热真菌属、威克汉姆酵母属等,长江中游派优势真菌为曲霉属、热子囊菌属、嗜热真菌属,江淮派优势真菌有曲霉属、热子囊菌属、耐干霉菌属等,川派优势真菌有热子囊菌属、嗜热真菌属、曲霉属等;吴树坤等[8]利用高通量测序发现四川不同地区浓香型白酒大曲中优势真菌为嗜热真菌属、曲霉属、嗜热子囊菌属、丝衣霉属、节担菌属;施思等[9]利用高通量测序技术发现浓香型白酒大曲在贮存过程中的真菌群落结构不断调整,毕赤酵母属、根霉属及横梗霉属成为最终的优势菌群;Li[10]对不同工艺的中、低温大曲的微生物群落变化进行研究,发现在不同环境作用下,微生物群落结构会发生改变。
基于高通量测序技术,已经对细菌群落演替规律进行了相关分析[11]。本文以中高温大曲为研究对象,采用高通量测序技术对培菌发酵几个主要阶段的真菌群落进行分析,研究各阶段优势真菌的分布、丰度及其在培菌发酵过程中的演替规律,为进一步剖析制曲发酵机理提供可靠的微生物信息。
1 材料与方法
1.1 材料、试剂及仪器
大曲样品:中高温大曲培养过程中入房(编号:RF,发酵0 天)、并房(编号:BF,发酵5 天)、上架(编号:SJ,发酵9 天)、大火(编号:DH,发酵15天)、下架(编号:XJ,发酵21 天)、出房(编号:CF,发酵35 天)各个阶段的混合样,分别粉碎、混匀、装袋,于-80 ℃保存备用。
试剂耗材:TaKaRa Premix Taq®Version 2.0、E.Z.N.A.® Gel Extraction Kit (Omega,USA)、NEBNext® Ultra ™II DNA Library Prep Kit for Illumina®(New England Biolabs,USA)。
仪器设备:Novaseq 6000 PE250 平台、美国Bio-Rad Laboratory S1000 型PCR 仪、Thermo Nano-Drop One。
1.2 实验方法
按照标准流程对大曲样品进行DNA 提取,以ITS5-1737F 和ITS2-2043R 为引物进行PCR 扩增,每个样本进行3 个重复,并将同一样本的PCR 产物进行混合,然后电泳检测、切胶纯化、建库测序。
2 结果与分析
2.1 测序有效性分析
中高温制曲各阶段真菌的richness 曲线如图1所示。结果显示,随着测序深度的增加,各样本的richness 指数曲线都是先增加后逐步趋于平缓,说明测序数据量足够大,可以反映中高温制曲各阶段样本中的绝大多数物种信息。
2.2 Alpha多样性分析
对中高温制曲各阶段的真菌群落进行Alpha多样性分析,结果见表1。所有样本的coverage 均大于0.99,说明该结果能够真实反映真菌群落结构。刚入房时,真菌丰度与菌群多样性都较低,说明自然接种的微生物种类与数量都较低;主发酵期,曲心温度升高,真菌大多不耐热,导致并房后真菌的丰度与菌群多样性都降至最低;此后进入潮火期,随着曲心温度进一步升高,能够适应高温的真菌种类在大曲中不断富集,因此上架时的真菌丰度与菌群多样性都开始升高;随着曲心温度保持60 ℃左右进入占火,嗜热真菌进一步增殖,曲皮水分也丧失殆尽,大火期的真菌丰度与菌群多样性进一步升至最高值;经过大火期的高温驯化与淘汰,下架时的真菌丰度与大火期相当,但菌群多样性降低;此后进入养曲期,曲心水分进一步降低,干燥缺水的环境逐渐不适合真菌的生长,因此出房时的真菌丰度基本不变,菌群多样性也进一步降低。
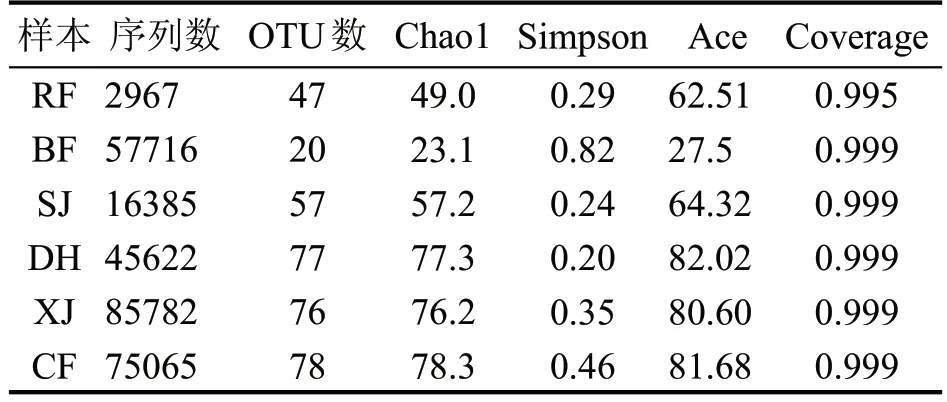
表1 大曲培养各阶段真菌Alpha多样性指数
2.3 OTU分布Venn分析
通过对中高温制曲各阶段的OTU 进行统计分类,Venn 分析结果如图2 所示。结果显示,真菌有11 个OTUs 是共有的;从入房到并房,真菌无变化;上架时缓慢增加;进入大火期,真菌快速增加至最高点;下架时,真菌缓慢下降;出房时,真菌有所回升。

图2 大曲培养各阶段真菌OTUs交叠Venn图
2.4 不同分类水平的物种变化
通过合并相同物种分类的OTUs,从门、纲、目、科、属不同分类水平,统计平均含量前15 的物种,其余的物种归于others 中,根据百分比绘制真菌的分布柱状图,结果见图3与图4。
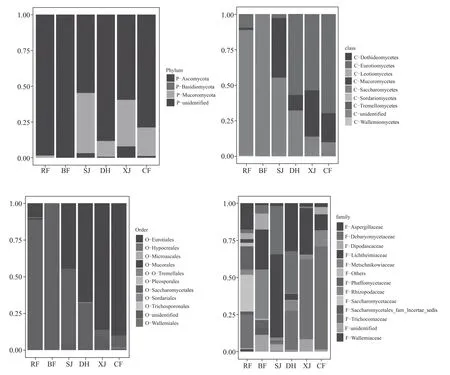
图3 大曲培养各阶段真菌在门、纲、目与科水平上的分布变化
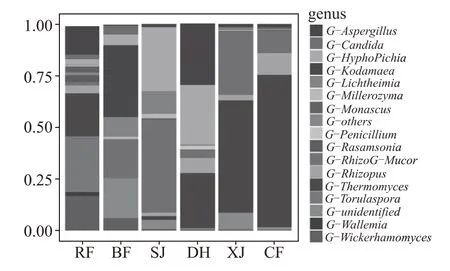
图4 大曲培养各阶段真菌在属水平上的分布变化
从图3 中可以看出,不同培养阶段大曲中的真菌都是以子囊菌门和毛霉门为主,两者占比超过90%以上,其中子囊菌门包括半子囊菌纲的酵母目和散囊菌纲的散囊菌目,毛霉门即毛霉纲的毛霉目。毛霉目前期很少,上架后才大量繁殖,丰度占比高达42 %,大火期间降至11 %,下架后回升至32%,出房后又缓落至20%;酵母目的丰度则先缓升后不断下降,入房时即高达88 %,并房后升至99%以上,之后开始快速下降,上架、大火、下架时分别下降至52%、31%、6%,出房时回升至8%;散囊菌目则先下降后上升,入房时占比10%,而后缓慢下降至3 %,大火后迅速生长繁殖,丰度占比高达57 %,而后维持较高水平,下架、出房时分别为53%、70%。
从科水平看,以相对丰度1 %为下限,共检测到12 个真菌科。入房时以酵母科、发菌科、曲霉科和法夫酵母科为主,占比分别为25%、22%、18%、16 %,此外还有丰度不足5 %的双足囊菌科、德巴列酵母科、根霉科、横梗霉。酵母科在并房后消失殆尽;发菌科先快速减少后逐渐增加,并房时消失殆尽,上架后开始逐渐增加,各个阶段的丰度分别为2%、26%、54%、70%;曲霉科先减少、后增加、而后又减少,仅大火期高达32%,其他阶段丰度都在1%~2%;法夫酵母科呈减少趋势,并房时占比6 %,之后降至1 %以下;双足囊菌科从入房时的4%增加至并房时的11%,而后降至1%以下,出房时回升至5%;德巴列酵母科在大曲培养各阶段呈先增加后减少趋势,上架和大火期丰度较高,分别为32 %、28 %;根霉科则呈波浪式变化,出房时丰度达10 %;属于毛霉目的横梗霉也大致呈先增加后减少趋势,上架时丰度最高达56 %,出房时为11%。
在属分类水平上,以相对丰度1 %为下限,共检测到16 个真菌属,结果如图4 所示。入房时以孢圆酵母属、嗜热真菌属、威克汉姆酵母属和曲霉属为主,占比分别为26%、21%、17%、14%;并房后,微生物群落发生显著变化,主体微生物调整为柯达酵母属、根毛霉属、横梗霉属、威克汉姆酵母属、生丝毕赤酵母属、假丝酵母属,丰度分别为35 %、19%、10%、6%、5%、5%;上架后,根毛霉属、生丝毕赤酵母属、横梗霉属的丰度分别增长至45 %、31 %、11 %;大火期,主体微生物转变为30 %的曲霉属、29%的生丝毕赤酵母属、27%的嗜热真菌属和7%的根霉属;下架后,嗜热真菌属、根毛霉属大量增长至54 %、31 %;出房后,嗜热真菌属占据绝对优势,丰度高达73%,此外还包括11%的根毛霉属和10%的根霉属。
2.5 群落分布特征
通过PCoA 主坐标分析与丰度聚类热图研究中高温制曲各阶段真菌群落分布特征,结果显示,图5 中的第一轴能够解释数据64.6%的变量,第二轴能够解释数据18.5 %的变量,两轴和为83.1 %。结合样品在分布图中的距离及其丰度聚类热图,可将真菌变化趋势的类型分为四类:(1)在入房和并房阶段,距离接近、同处于第二象限且聚为一类,说明这两个阶段的真菌群落结构相似,可归为同一阶段,其中孢圆酵母属主要在入房阶段,柯达酵母属主要在并房阶段;(2)上架时,样本与入房和并房阶段距离较远,且单独位于第三象限,可单独归为一类,此时伴随温湿度等环境因子的变化,大曲中水分营养充足,微生物结构不断进行适应性调整,横梗霉属大量生长至最高丰度,假丝酵母属、粉状米勒氏酵母属等丰度也升高;(3)大火期,样本与入房、并房和上架阶段的距离都较远,且单独位于第四象限,结合丰度聚类热图,也单独归为一类,此时温度等环境因素达到极限,不耐热微生物逐渐被淘汰,生丝毕赤酵母属、曲霉属、节担菌属、青霉属大量生长至最高丰度;(4)下架虽然与大火期距离较近,但下架和出房样本同位于第一象限且距离不远,结合丰度聚类热图可将下架和出房样本聚为一类,两者可归为同一阶段,此时随着品温等环境因子渐趋温和,微生物利用曲心残留的水分和营养进行生长繁殖与演替,根毛霉属、粉状米勒氏酵母属、横梗霉属及未知属在下架时丰度占据绝对优势,柯达酵母属、假丝酵母属、威克汉姆酵母属、根霉属在出房时丰度占据绝对优势,嗜热真菌属、红曲霉属、罗萨氏菌丰度在下架和出房时都较高。
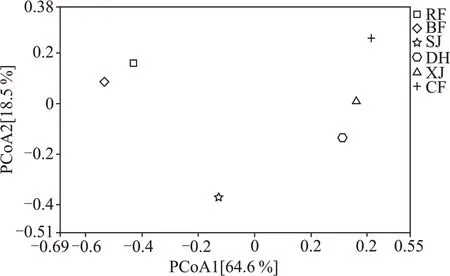
图5 大曲培养各阶段真菌主坐标分析图
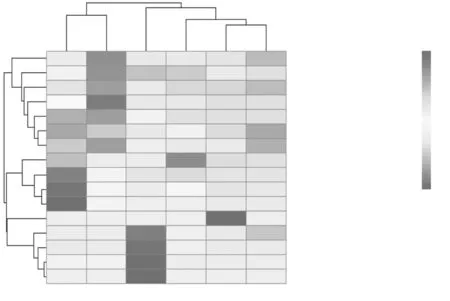
图6 大曲培养各阶段真菌群落丰度聚类热图
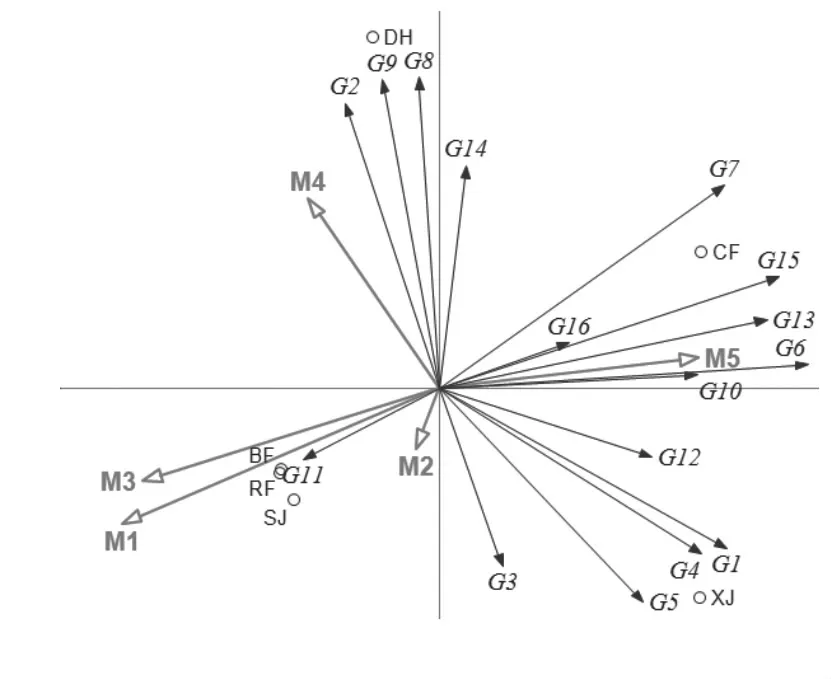
图7 理化因子及酶活指标与真菌优势属的RDA分析
2.6 真菌群落与理化因子关联分析
利用R 语言中RDA 分析和作图,分析中高温制曲过程中的真菌群落结构与理化指标之间的相关性。理化因子关联分析发现,水分、糖化力、酯化力与真菌群落结构的相关性较高,发酵力其次,酸度与真菌群落结构的相关性较低,说明真菌主要对水分和酶活有较大影响,对酸度的影响较小。就主要优势属看,发酵力与生丝毕赤酵母属相关性较高,与曲霉属、青霉属也有一定的相关性;酯化力与嗜热真菌属、红曲霉、威克汉姆酵母属相关性较高,与罗萨氏菌、柯达酵母属、根霉属也有一定的相关性;水分和糖化力则与孢圆酵母属的相关性较高。
3 总结与展望
本研究采用高通量基因测序技术对中高温制曲培养各阶段真菌不同分类水平的组成与丰度进行了系统的研究,结果表明:在门水平上以子囊菌门和毛霉门为主,其中子囊菌门包括半子囊菌纲的酵母目和散囊菌纲的散囊菌目,毛霉门即毛霉纲的毛霉目;在科水平上,以酵母科、发菌科、曲霉科和法夫酵母科为主;属水平上,嗜热真菌属、根毛霉属、根霉属、曲霉属、生丝毕赤酵母属、横梗霉属、威克汉姆酵母属分别在制曲过程不同时期占据主导地位,前期主要是酵母属,为大曲的酿酒和生香提供菌源,后期主要是耐高温霉菌类微生物,为大曲提供糖化、液化以及生香相关的微生物。唐清兰等[12]采用高通量测序技术跟踪浓香型白酒大曲制曲过程真菌群落结构变化,发现伊萨酵母属、嗜热子囊菌属、曲霉属分别在制曲过程不同时期占据主导地位。不难发现,两类大曲的真菌种类及其丰度存在明显差异,这是由于大曲采用开放式生产、自然接种,环境、原料、工艺等对大曲微生物区系均有直接影响。
此外,本文通过RDA 分析首次发现真菌与水分和酶活关系密切。其中发酵力与生丝毕赤酵母属相关性较高,生丝毕赤酵母属是白酒发酵中的重要功能微生物,可以代谢产生风味物质,如乙酸乙酯、4-羟基-2-丁酮等[13-14]。酯化力与嗜热真菌属、红曲霉、威克汉姆酵母属相关性较高,其中嗜热真菌属耐高温,最高生长温度为50 ℃或以上,可产生嗜热酶,包括纤维素酶、蛋白酶、淀粉酶、脂肪酶等[12];红曲霉能分泌大量酯酶,对碳酸十四乙酯和苯乙醇的形成有促进作用[15];威克汉姆酵母属能够将原料中的前体物质转化成酯、酸、醇、醛等风味物质,对白酒风味形成有重要作用[16]。水分与孢圆酵母属的相关性较高,孢圆酵母属是非酿酒酵母菌,但通过其独特的次生代谢物也可对酒的感官特性作出积极贡献[15]。
通过研究,明确了中高温制曲过程中真菌群落结构的演替规律,并首次解析了真菌种类与大曲的发酵和生香之间的潜在关系,有助于中高温大曲真菌多样性形成机制的解析,从而提高对大曲微生物认识的深度和广度,为中高温大曲生产提供参考与借鉴。下一步,我们将结合微生物与非生物因素进行关联性分析,如温度、湿度、氧气、门窗管控等,以全面准确地研究中高温大曲微生态演变的内在驱动力。
猜你喜欢
食品与生物技术学报(2021年9期)2021-09-28
中国调味品(2021年3期)2021-03-23
酿酒科技(2020年7期)2020-12-19
食品与发酵工业(2020年14期)2020-07-29
遗传(2019年11期)2019-11-28
中国调味品(2019年11期)2019-11-15
微循环学杂志(2017年3期)2017-08-30
中国调味品(2017年2期)2017-03-20
化学工业与工程(2016年6期)2016-04-10
湖南农业大学学报(自然科学版)(2015年3期)2015-07-13
