
小兴安岭南部2种红松天然林优势树种的空间结构和生态位特征
2023-11-25 10:05王亚飞蒋乙东段文标朱海媖王秋宇
中南林业科技大学学报 2023年10期
王亚飞,蒋乙东,段文标,朱海媖,王秋宇
(东北林业大学 林学院,黑龙江 哈尔滨 150000)
1993年12月29日正式生效的《生物多样性公约》和2021年10月12日在中国昆明召开的《生物多样性公约》第十五次缔约国大会(CBD COP15)通过的《昆明宣言》,旨在实现维系生物多样性的目标。目前,我国的森林生态系统通过多年的可持续经营取得了重大的成效,但是,以人工林为主的森林经营模式降低了森林生态系统内的物种多样性,进而影响了森林的功能多样性和可持续性[1-2],而维系森林生态系统内的物种多样性是森林生态系统经营的关键和核心所在。德国学者提出的近自然经营为维系森林生态系统的生物多样性提供了良好的方向,即以借鉴天然林的演替及发展趋势,通过建立混交林、计划性抚育伐等措施,加速森林生态系统物种多样性和功能多样性的恢复[3]。而弄清天然林内植物的资源分配特征及空间分布特征是近自然经营的技术关键[4]。
生态位特征是描述群落中不同种群时空资源分配的定量分析的重要指标,能够说明种群在群落中的资源分配状况及不同种群间资源占有的相似度等,反映了种群的地位级及种群间的资源利用关系[5]。随着生态位研究的逐渐深入,出现了多种多样的生态位理论,如经典的生态位理论(空间生态位[6]、功能生态位[7]、n维超体积[8])、资源利用函数[9]以及扩展的生态位理论[10]。通过对区域内各种群的生态位特征进行分析,可以更好地对该区域内种群的当前态和未来态进行分析和预测,进而更加合理地利用和保护自然资源。生态位研究表明,高水平的物种多样性使群落功能性更加丰富,这也意味着不同功能性的植物之间更容易形成资源生态位和功能生态位的互补,从而增强了群落对资源的利用效率,并且使种群之间的关系更加稳定[11-12]。
生态位特征仅能从资源分配角度对群落的维系进行揭示,但是缺乏对森林结构的解析。而空间结构特征体现了森林中林木的点格局及其相关属性的分布特征,因而可与生态位特征形成互补,从资源和空间两方面对群落动态进行表征。在空间结构特征参数中,角尺度反映了森林中林木的分布格局,混交度体现了群落内种群的混交状态,而大小比数则说明了森林内树木的大小分化特征[13-14]。目前,国内在生态位特征[15—16]和空间结构特征[17-19]方面对不同气候带和不同林型进行了较多的研究,但是在小兴安岭地区的研究较少,特别是对生态位特征和空间结构特征二者之间关系的研究更为少见。
20世纪以来,历史的原因造成大量红松针阔混交林受到破坏。小兴安岭亦是如此,该区域内地带性针阔混交林和针叶林遭到破坏。本研究选择未经人为扰动的阔叶红松林和云冷杉红松林为研究对象,基于样地调查数据,对其优势树种的生态位宽度、生态位重叠和相似程度、角尺度、大小比数以及混交度进行分析,阐释2种林型红松天然林优势树种的生态位特征及空间结构特征。通过生态响应及其速率预测2种林型红松天然林发展趋势,并通过相关分析揭示其生态位特征和空间结构特征之间的关系,为红松林生态系统的抚育和保护等提供基础数据和理论依据。
1 研究地概况
样地位于小兴安岭南部的凉水国家级自然保护区内,该保护区是以保护红松林生态系统为目的而建立的。小兴安岭属北温带大陆性季风气候区,四季分明,年平均气温为±1 ℃[20]。所选云冷杉红松林地处海拔490 m,其优势树种为红松、臭冷杉和红皮云杉等,伴生树种为白桦、色木槭、青楷槭和紫椴等,灌木层主要为毛榛子Corylus mandshurica和刺五加Acanthopanaxssenticossus等,草本主要有苔草Carexspp.、酢浆草Oxalis corniculata和荨麻Urticafissa等。所选阔叶红松林地处海拔390 m,上层树种以山杨Populus davidiana、枫桦、白桦、云冷杉和红皮云杉等为主,灌木层和草本层植物与云冷杉红松林相似。
2 研究方法
2.1 样地设置与调查
2017年8月进行踏查,选择林隙干扰下有代表性的2种类型的典型红松天然林(阔叶红松林和云冷杉红松林,详见表1)作为试验对象,在阔叶红松林的区域内设置了一块1 hm2(100 m×100 m)的样地,在云冷杉红松林的区域内设置了一块1.2 hm2(100 m×120 m)的样地。对每块样地内胸径≥5 cm的林木进行每木检尺。采用网格法将每块样地划分为10 m×10 m的小网格,对样地内每棵树木进行准确定位。使用布鲁莱斯测高器测量其树高,并记录每株检尺木的种类和胸径等。

表1 样地基本信息Table 1 Information about the sample sites
2.2 生态位特征值计算方法
在进行生态位特征值计算时,考虑到天然林内具有林隙,设置的研究单元较小时可能被林隙完全覆盖,所以按照20 m×20 m将样地重新划分网格,然后以此来进行生态位特征的分析。
2.2.1 生态位宽度的计算
分别计算了样地内所有种群和林隙内所有种群两部分的生态位宽度。生态位宽度采用Levins的计算方法[21],其公式为:
式中:Bi为种群i的生态位宽度,nij为种群i在样方j中的资源利用(以种群i在样方j中的重要值之和表示),Ni为种群i的总数量。
2.2.2 生态位重叠的计算
生态位重叠的计算方法采用Levins重叠指数[22],其计算公式为:
式中:Lih和Lhi分别为树种i相对于树种h以及树种h相对于树种i的生态位重叠指数,B(L)i和B(L)h分别为树种i和树种h的生态位宽度,Pij和Phj与上节Pij同义。
2.2.3 生态位相似的计算
生态位相似计算公式为[23]:
式中:Cih表征了物种i和h的生态位相似比例,取值范围为[0,1]。
2.2.4 生态响应及响应速率的计算
生态位响应反映了植被在样地内处于进攻型或者淘汰型,进而对种群动态进行预测,具体计算公式为[24]:
式中:ΔLih为树种i占用其他树种的资源总量,即生态响应。R为树种的生态响应速率,B(L)为树种的生态位宽度。公式中(1)表征了树种i占用其他树种的总资源量,(2)表征了树种i被其他树种所占用的资源量,因此,当(1)>(2)时,树种i为进攻性树种,即树种i相较于其他树种更加适应此样地,未来一定时间内树种i在样地中的资源占用量、数量等会越来越多;反之,树种i则为淘汰性树种,随时间推移数量减少,资源占用量更加少;(1)=(2)时,树种i为中性树种,无强烈的外力干扰会保持在样地中的状态。
2.3 空间结构特征指标的计算方法
空间结构特征指标计算采用Winkelmass 1.0软件完成,具体的计算公式如下:
式中:Wi为角尺度,Mi为混交度,Ui为大小比数,Zij表征对象木与相邻木的夹角是否小于标准角α0,Vij表征对象木与相邻木是否为同种树种,Kij表征对象木是否大于相邻木。角尺度、混交度和大小比数的取值范围均为[0,1]。其中,当Wi为0、0.25、0.5、075、1时,其分布处于非常均匀、均匀、随机、不均匀、非常不均匀5个程度。当Ui为0、0.25、0.5、075、1时,其在样地中的地位分别处于优势、亚优势、中庸、劣汰、非常劣汰5个等级。当Mi为0、0.25、0.5、075、1时,其混交程度分别处于零度、弱度、中度、强度和极强度5个水平[25-26]。
2.4 生态位特征和空间结构的相关性分析
为了表征生态位特征和空间结构特征的关系,采用SPSS 21.0软件对其相关性进行了分析。首先检验2个样地内优势树种的生态位特征和空间结构特征指标的正态分布,如果符合正态分布,后续相关性分析使用Pearson相关系数,否则使用Spearman相关系数。
3 结果与分析
3.1 2种林型的乔木组成特征
阔叶红松林和云冷杉红松林是2种重要的天然红松林。由表2可知,云冷杉红松林内,红松的重要值最高,为15.58%。其次为白桦、红皮云杉、色木槭等。榆树在此样地的重要值最低。重要值排名前八的树种的重要值之和为71.95%。由表3可知在阔叶红松林中,红皮云杉的重要值最高,为15.49%,枫桦次之。鼠李、黄檗、花楷槭、红松、水曲柳等树种的重要值在此样地中偏小。重要值排名前八的树种重要值之和为70.07%,分别为红皮云杉、枫桦、暴马丁香、毛赤杨、色木槭、臭冷杉、春榆、紫椴。

表3 阔叶红松林内乔木各种群的重要值Table 3 Importance values of tree groups in natural P.koraiensis coniferous and broad-leaved mixed forest
3.2 2种林型的生态位宽度特征
在云冷杉红松林的优势树种当中(图1),紫椴的生态位宽度最大,青楷槭、色木槭、臭冷杉、红松次之,但是其生态位宽度值均大于0.500,说明这些树种此样地内的适应性较强。水曲柳、白桦、红皮云杉的生态位宽度偏小,说明其在此样地内的资源分配较少,对此样地的适应性较差。而在阔叶红松林中(图2),红皮云杉和暴马丁香的生态位宽度明显高于其他6种优势树种的生态位宽度,并且其他树种的生态位宽度均小于0.500,说明红皮云杉和暴马丁香在该样地的占据了较多的资源。

图1 云冷杉红松林内优势树种的生态位宽度Fig.1 Niche width of dominant tree species in P.koraiensis-A.nephrolepis-P.koraiensis forest
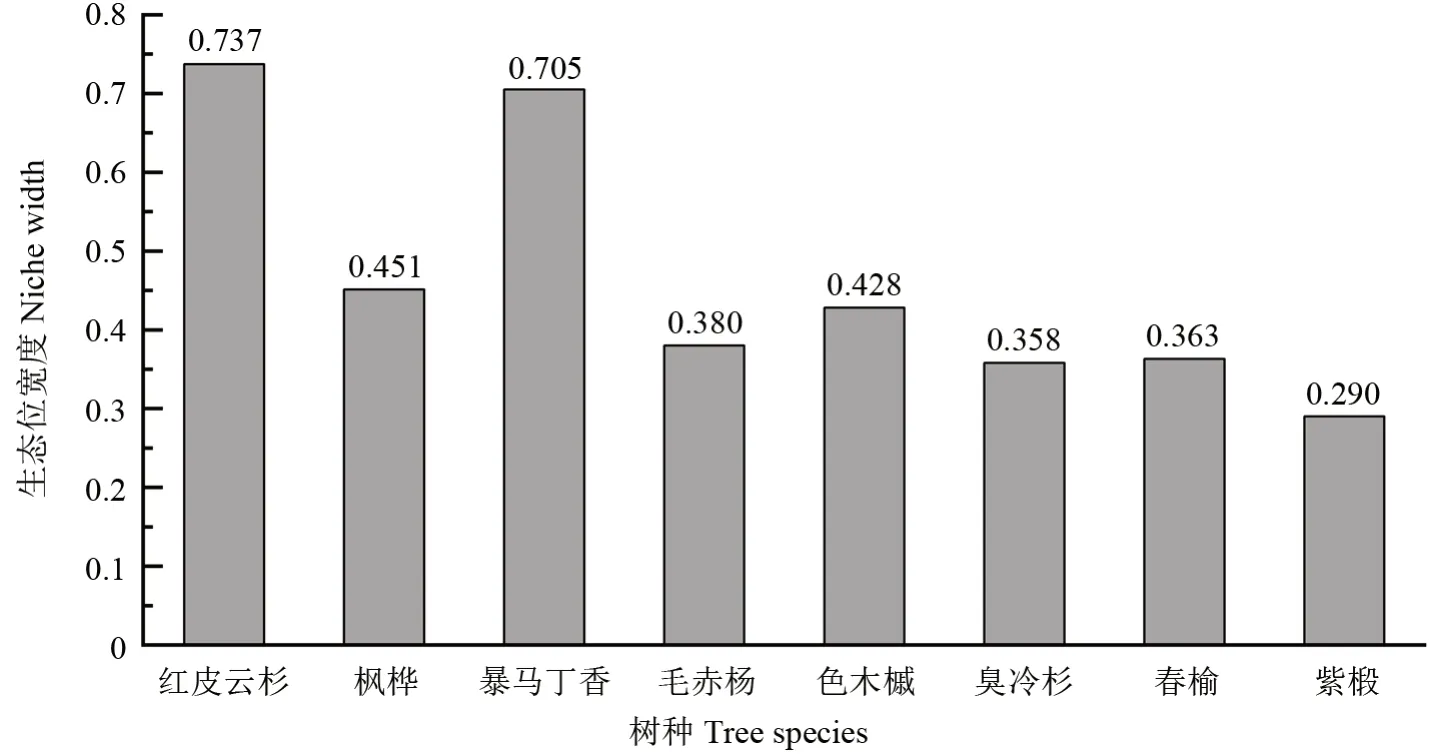
图2 阔叶红松林内优势树种的生态位宽度Fig.2 Niche width of dominant tree species in natural P.koraiensis coniferous and broad-leaved mixed forest
3.3 2种林型的生态位重叠特征
在云冷杉红松林中,红松与臭冷杉、青楷槭、紫椴、色木槭之间的生态位重叠度较高(表4)。白桦的生长并未占据其他树种的生态位,但是红皮云杉、臭冷杉、紫椴和青楷槭占据了白桦的生态位。紫椴、青楷槭、臭冷杉的生态位相对于其他优势树种的生态位重叠度最大,说明紫椴、青楷槭、臭冷杉的生长占据其他树种的较多。色木槭与其他树种相互的重叠度都较高。而水曲柳并未侵占其他优势树种的生态位,但是其他树种相对于其生态位的重叠程度最高。在此样地中,生态位重叠程度最高的为紫椴相对于水曲柳和青楷槭相对于红松,分别达到了0.036和0.031。

表4 云冷杉红松林内优势树种间的生态位重叠特征†Table 4 Niche overlap among dominant tree species in P.koraiensis-A.nephrolepis-P.koraiensis forest
而在阔叶红松林内优势树种的生态位重叠程度均较高,并且其他树种对于红皮云杉的生态位也有一定的重叠(表5)。枫桦与暴马丁香、色木槭、臭冷杉、紫椴的生态位重叠程度较高,暴马丁香相对于其他树种的生态位重叠程度较高,但是与其他树种的生态位重叠程度较低。色木槭除了与红皮云杉、枫桦、暴马丁香的生态位重叠程度较高外,与紫椴和春榆也具有较高的生态位重叠程度,其中与紫椴的生态位重叠程度最高,达到了0.037,紫椴对色木槭的重叠程度和与其他树种的重叠程度相比也最高,达到了0.025。

表5 阔叶红松林内优势树种间的生态位重叠特征Table 5 Niche overlap among dominant tree species in natural P.koraiensis coniferous and broad-leaved mixed forest
3.4 2种林型的生态位相似特征
由表6可知,在云冷杉红松林内,红皮云杉和水曲柳的生态位相似度最高,为0.864,紫椴—红皮云杉、臭冷杉—白桦等种对的生态位相似度最低,均低于0.700。通过对优势树种种对的生态位相似度进行统计分析发现,在云冷杉红松林当中,其均值较为相似。其中红松最高,与其他优势树种的生态位相似度的均值为0.790,紫椴的最低为0.740。样地内优势树种整体的均值为0.760。由表7可知,在阔叶红松林内,各优势种相似度均值为0.790。毛赤杨与紫椴的生态位相似度最高,达到了0.988,说明毛赤杨和紫椴在本样地内的资源需求最为相似。其他生态位相似度较高的种对有臭冷杉—紫椴、春榆—紫椴、毛赤杨—色木槭等,其相似度均大于0.900。而红皮云杉与春榆的生态位相似度最低,为0.639。并且对优势种与其他优势种的生态位相似度的统计发现,红皮云杉与其他优势树种的生态位相似度最低,均值仅有0.690。而毛赤杨、臭冷杉和色木槭与其他树种的生态位相似程度都较高,均值均为0.820。

表6 云冷杉红松林内优势树种生态位相似特征Table 6 Niche similarity of dominant tree species in P.koraiensis-A.nephrolepis-P.koraiensis forest

表7 阔叶红松林内优势树种生态位相似特征Table 7 Niche similarity of dominant tree species in natural P.koraiensis coniferous and broad-leaved mixed forest
3.5 2种林型的生态响应及响应速率
由表8可知,在云冷杉红松林中,红松、白桦、红皮云杉和水曲柳的资源被色木槭、臭冷杉、紫椴和青楷槭占据。其中,红松的衰退速率较快,达到了-204.95;白桦的衰退速率较慢,仅为-0.51;色木槭和臭冷杉的发展速度较快,生态响应速率分别达到了1 764.84和300.79。从整体来看,生态响应之和为-0.001 1,说明优势树种的资源在被其他非优势树种略微占据,但是优势树种整体具有较高的发展速度,为1 908.26。在阔叶红松林内,优势树种占据非优势树种的资源较多,达到0.515 2,发展速率达到237.86。在阔叶红松林的优势树种中除了紫椴以外,其他优势树种均为发展中的树种,其中暴马丁香和红皮云杉占据其他树种的资源较多,分别为22.91和21.80,但是春榆、臭冷杉、毛赤杨和色木槭的发展速度都较快。整体而言,云冷杉红松林处于稳定阶段,阔叶红松林处于发展阶段。
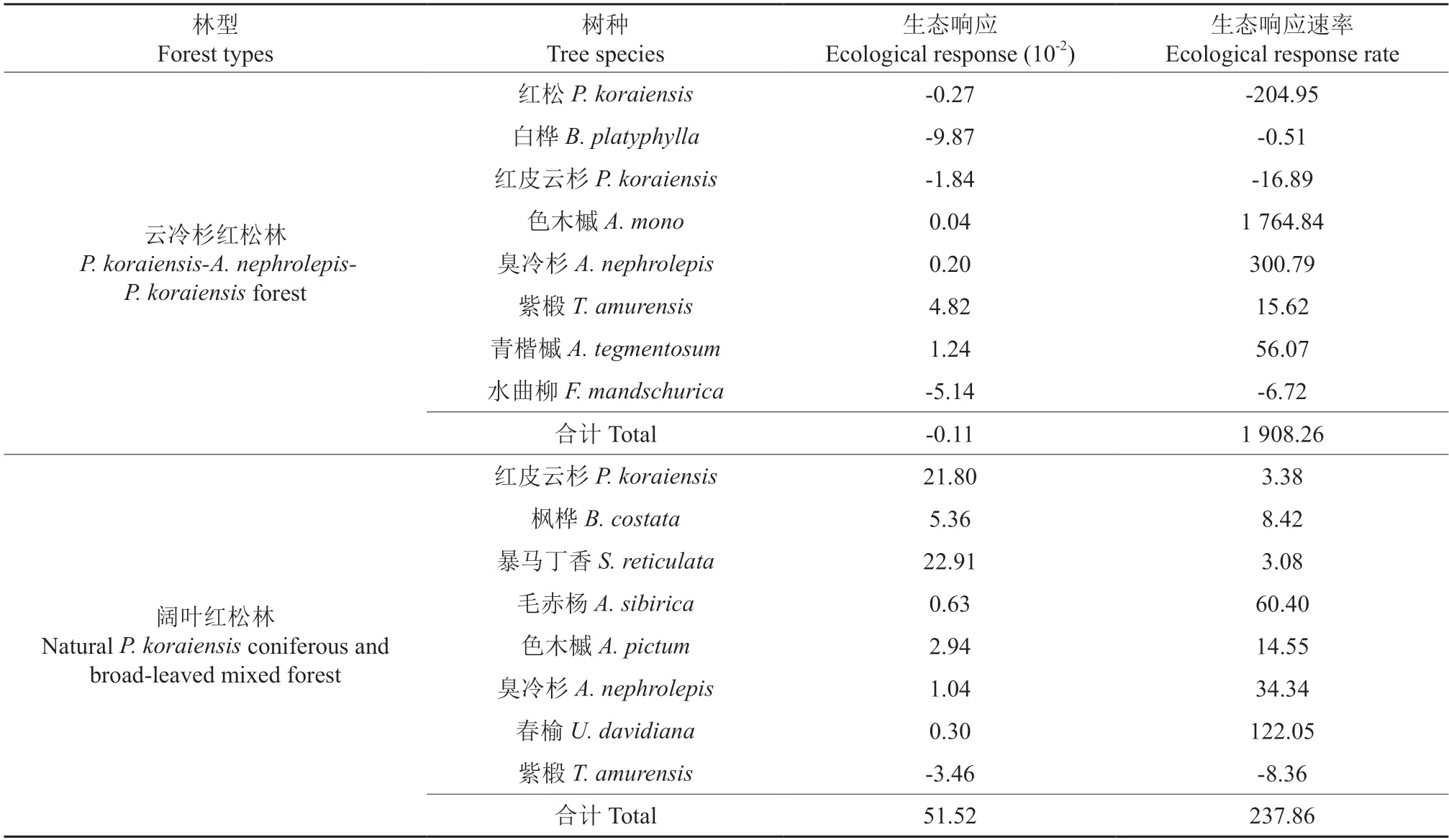
表8 优势树种的生态响应及速率Table 8 Ecological response and rate of dominant tree species
3.6 2种林型内优势树种的空间结构特征
由表9可知,云冷杉红松林和阔叶红松林内优势树种的角尺度均分布在0.5左右,为随机分布。在大小尺度方面,云冷杉红松林的红松的大小比数最小,为0.10,是该样地中的优势树种。白桦、红皮云杉、水曲柳和紫椴在云冷杉红松林内大小比数均大于0.4且小于0.5,更倾向于中庸树种,臭冷杉的大小比数为0.5,为亚优势树种。色木槭和青楷槭的大小比数分别为0.58和0.63,处于中庸和亚优势之间。在阔叶红松林中,枫桦的大小比数最小,为0.19,倾向于亚优势树种;毛赤杨、色木槭、臭冷杉、春榆和紫椴的大小比数分布在0.5左右,为该样地的中庸树种;暴马丁香的大小比数最大,为0.64,在大小方面相较于其他树种是该样地的劣汰树种。在混交度方面,云冷杉红松林内有红松、水曲柳和臭冷杉3种树种的混交度介于强度混交和极强混交之间,其余5种树种介于中度混交和强度混交之间。在阔叶红松林内,红皮云杉和枫桦介于强度混交和极强混交之间,臭冷杉和春榆属于强度混交,色木槭和紫椴介于强度和中度混交之间,而暴马丁香和毛赤杨的混交度最低,仅为0.48,介于中度混交和弱度混交之间。
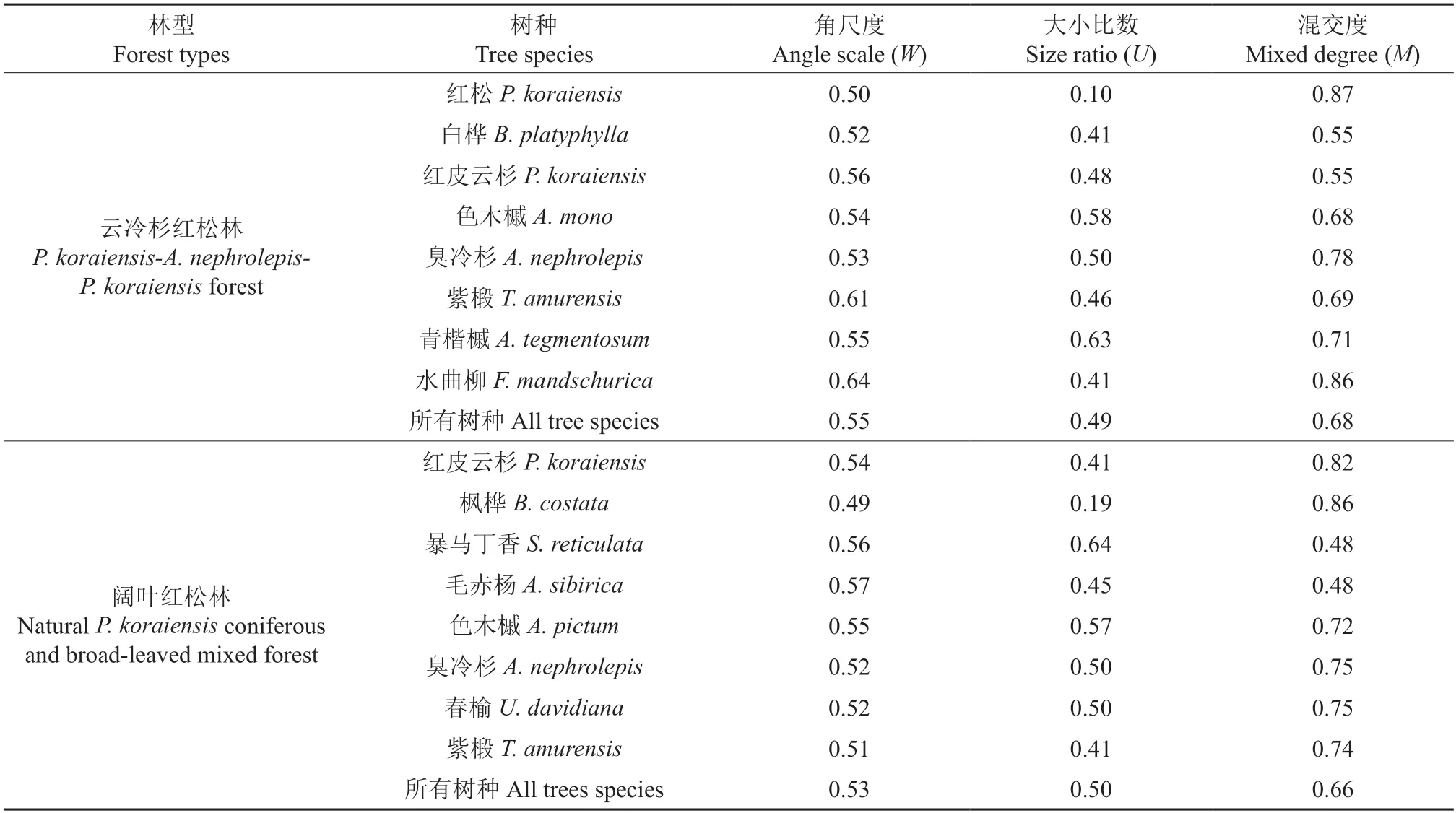
表9 2种林型红松天然林内优势树种的空间结构特征Table 9 Spatial structure characteristics of dominant tree species in two types of natural P.koraiensis forest
通过对比2种林型红松天然林的空间结构特征发现角尺度、大小比数和混交度均相近。其中,角尺度结果显示2种林型的点格局均为随机分布,2种林型大小比数等级均为中庸强度,并且其混交度均介于强度和中度之间。
3.7 2种林型内生态位宽度特征与空间结构特征之间的关系
对各指标数据的正态分布进行检验,结果如表10所示,生态响应速率的偏度值不处于±1.96之间,因而其不服从正态分布。因而,使用Spearman相关系数对其他指标的相关性进行分析,指标之间的相关系数如表11所示,其中混交度与大小比数具有显著的负相关关系,相关系数为0.522,混交度与生态位响应速率之间具有显著的正相关关系,相关系数为0.572。生态位宽度和生态响应之间具有极显著的正相关关系,相关系数为0.691。

表10 所选指标的正态分布检验结果Table 10 Normal distribution test results of selected indicators

表11 生态位和空间结构特征各指标间相关性†Table 11 Correlation between niche and spatial structure characteristics
4 讨 论
4.1 2种林型红松天然林内优势树种的生态位特征
生态位特征反映了林分中资源的分配模式,而生态位宽度特征则反映了种群由于自身的生物学特征而对不同状态的林分产生的相应的适应能力[27]。小兴安岭内的天然林以红松针阔混交林系统为主,而阔叶红松林和云冷杉红松林分别处于该系统演替的中期和后期,因而其内的优势物种及优势物种出现了明显的差异性。在云冷杉红松林内,其优势树种以耐阴树种红松、臭冷杉、红皮云杉、紫椴、水曲柳等及主要的伴生树种青楷槭、色木槭等为主。白桦为阳性先锋树种,其生物学特性对此样地并没有太高的适应性,因而其生态位宽度较低。红松为该地区的顶级优势树种,而云冷杉红松林属于暗针叶林,也是顶级优势群落,据此可以看出,在稳定的顶级群落阶段,林中生态位宽度的调配受建群种的生物学特性驱动。在其他研究中,以南方铁杉为建群种形成的顶级群落中也表现出了南方铁杉及其伴生树种的生态位特征较宽的现象[28]。而在阔叶红松林中,阳性先锋树种和耐荫树种均有优势分布,并且其生态位宽度没有出现明显的差异性。阔叶红松林是阔叶林向针叶林演替的中间阶段[29],随着该林分中耐阴树种的生长,阳性先锋树种的资源利用谱变窄,逐渐失去其在林中的优势地位。
生态位重叠反映了种群之间在某区域内资源利用的重合度[30],生态位相似反映了种群在研究区内的资源利用的相似度。在以往的研究当中,研究者发现生态位越宽的树种,其生态位重叠值越小[16]。但是由于本研究选择的生态位重叠度方法不同,说明了目标树种相对于其他树种的生态位重叠特征。在物种分布较为稳定的云冷杉红松林中,生物学特性决定了树种间的分布及生态位重合度,例如红松与具有伴生关系的青楷槭和色木槭等。并且在该样地内,红松与其他优势种群较高的相似度也反映了生物学特性是该样地内资源分配中的主导。但是在阔叶红松林中,物种之间对资源的竞争较为激烈,从而形成了生态位宽度较宽的树种对于生态位宽度较窄的树种的生态位重叠度更高。并且阔叶红松林内,生态位宽度越高的树种与其他树种的生态位相似度越低,这与黄绍辉等[31]、杨俊等[32]和龙翠玲[33]研究结果相似,这是优势树种种群自我保护的一种策略。研究发现在资源压力较为紧张时,种群的聚集分布能够保护种群的发育,保证种群占据更宽的资源谱。
综合生态位宽度、生态位重叠和生态位相似这3种特征可以发现,不同演替阶段林分内资源分配的驱动因素各不相同。在群落发育阶段,资源分配主要是靠种群间竞争来驱动,而在稳定的顶级群落,主要是靠建群种的生物学特性来驱动。
4.2 2种林型红松天然林的空间结构特征
林分的空间结构是森林发展和维系的重要驱动因子[34-36]。林分内林隙等现象就是通过改变林分的空间结构来防止林分处于单一的顶级群落,增加了林分内的异质性斑块,为其他树种的生长创造空间和机会。本研究结果表明,云冷杉红松林和阔叶红松林均处于随机分布的阶段,但是阔叶红松林的结果与前人的结果不符。前人的研究结果均表明,当群落处于发育阶段时,种群分布格局为聚集分布,但是当群落达到顶级群落时,种群分布格局达到随机分布。本研究中,阔叶红松林属于发育中的群落,但是其分布格局也是随机分布,这可能是由于林内具有大量的林隙,复杂的斑块构造使树种在该样地内更新的点位发生变化,从而造成了其分布格局的变化[37]。在大小比数方面,云冷杉红松林内的红松和天然针阔林内的白桦为优势级别,这是由于其平均胸径较大但是数量较少[38-39]。2种林型内其余树种均处于中庸,这也说明了未经人为干扰的天然林的大小比数均为中庸,具有良好的径级特征。在混交度方面,可以看出耐阴树种的混交度强于阳性先锋树种[40]。王大伟等[41-42]研究发现,小树围绕大树聚集生长产生环形结构,这一现象不仅在热带雨林中存在,在中国东北部的红松异龄混交林中同样存在。成熟的耐阴树种体型较大,树高较高,因而其下更容易生长小树,从而形成林分的垂直结构,加强光资源的利用效率。
4.3 种群生态响应速率预测
生态响应反映了调查时期种群生态位侵占或者侵占的程度,而生态响应速率反映了种群的发展或者衰退速率。2种林型内具有大量的林隙,因而2种林型的优势树种都处于发展阶段,都具有较高的生态位响应速率。其中,色木槭为红松的重要伴生树种,并且母树较多,因而其具有较高的发展速度。红松虽然是样地的建群种,母树较多,但是其种子是林中动物的重要食物,幼苗更新较少,因而其处于强烈的衰退阶段。臭冷杉和红松同属于松科,红松的减少加速了臭冷杉对其的取代作用,因而其具有较高的生态响应速率。相对于这3个树种,其他优势树种的发展和衰退速率均较慢。因为白桦为阳性先锋树种,其生态位被其他树种侵占较严重,这与上面的生态位重叠结果相似。因而未来该区域的云冷杉红松林中红松减少,云冷杉增加,有可能逐渐转化为云冷杉林[43]。
在阔叶红松林中,毛赤杨和云冷杉的发展速度最快。虽然阳性先锋树种的生态响应速率高于耐阴树种,但是耐阴树种的发展会填充主林冠层,造成阳性先锋树种的光资源减少。枫桦虽然是耐阴树种,但是其更新较少。综上,该样地未来的发展趋势为云冷杉林。
5 结 论
发展中的阔叶红松林的资源分配受种群竞争驱动,而稳定的云冷杉红松林的资源分配受建群种即红松的生物学特性驱动。2种红松天然林的空间结构均表现出点格局为随机分布,大小比数为中庸强度,混交度介于强度和中度之间,混交度与大小比数具有显著的负相关关系,混交度与生态位响应速率之间具有显著的正相关关系;生态位宽度和生态响应之间具有极显著的正相关关系。但是,本研究缺乏结合林下更新以及林隙的详细调查数据,未能详细地说明林隙存在对样地的空间和资源分配特征的影响,未来将对林隙年龄、大小及其对林分空间和资源分配特征的作用方面进行研究。
猜你喜欢
建筑与预算(2022年6期)2022-07-08
格言·校园版(2022年17期)2022-07-06
小哥白尼(野生动物)(2021年9期)2022-01-17
中国林副特产(2020年5期)2020-10-09
防护林科技(2017年7期)2017-09-01
现代营销(创富信息版)(2016年6期)2016-11-21
现代营销(创富信息版)(2016年12期)2016-02-08
建筑与预算(2014年7期)2014-10-14
满族文学(2014年5期)2014-09-22
建筑与预算(2014年5期)2014-04-11
