
α1,3 FUCT Ⅶ mRNA 及Slex抗原在非小细胞肺癌组织中的表达及其与转移的关系
2010-11-07 08:28文秀芳
中国医药指南 2010年5期
沈 庆 黄 静 文秀芳 黄 方 刘 霄 李 华
α1,3 FUCT Ⅶ是一种糖链加工酶,能催化岩藻糖基转移到糖蛋白糖链底物,它是产生Slex(又称为唾液酸化的路易斯X)的重要物质,大量文献报道Slex在大肠癌、胃癌、膀胱癌等肿瘤组织中呈明显增强表达[1-3],并与肿瘤的发生、发展、侵袭转移和预后密切相关。但α1,3 FUCT Ⅶ其与肿瘤转移的关系尚不十分清楚。本研究应用RT-PCR法及免疫组织化学催化放大(CSA)检测α1,3 FUCTⅦ基因mRNA及Slex在非小细胞肺癌组织、癌旁正常肺组织中的表达,探讨α1,3 FUCT Ⅶ基因mRNA及Slex的表达与临床病理因素的关系,以及其在肺癌发展过程中可能的作用。
1 材料与方法
1.1 标本
收集重庆市第三人民医院及协作单位2008年6月至2009年6月期间手术切除肺癌新鲜标本66例(男38例,女28例),平均年龄(51±16)岁。其中腺癌36例,鳞癌30例。癌旁正常肺组织选择远离癌组织(>5cm)的正常支气管黏膜和肺组织20例作为对照观察。所有标本均分为2份,另一份投入液氮速冻之后,3h内转移至-80℃低温冰箱冻存;一份经10%福尔马林固定,石蜡包埋、切片。
1.2 试剂
Rever Tre Ace-a-逆转录试剂盒购自TOYOBO公司,PCR System 9700购自Applied Biosy Stems,PCR Mix Taq购自天跟公司。鼠抗Slex单克隆抗体购自美国Millipore公司,SP试剂盒及0.3% Triton X100均购自北京中杉生物技术有限公司。
1.3 逆转录-聚合酶链式反应(RT-PCR法)
取标本20~100mg,以TRNzol试剂提取总RNA。将2μg RNA置于10µL逆转录反应体系中逆转录为cDNA,再行PCR扩增。根据已知α1,3 FUCT Ⅶ基因序列,由上海生物技术有限公司合成引物,上游:5'-CCC CGC AGC CAA GTC TAT GA -3',下游:5'-CCA CCC CTG CTT CCT GAC CT -3'(181bp)。GAPDH(内参照)引物上游:5'-TG GGG TGA TGC TGG TGC TGA GT- 3',下游:5'-AG GTT TCT CCA GGC GGC ATG TC -3'(500bp)。PCR反应条件:94℃预变性10min;94℃变性45s,63℃退火1min,72℃延伸1min。35个循环;72℃终延伸10min,扩增产物10g/LTBE琼脂糖凝胶电泳。溴化乙锭显影,凝胶成像系统(UVITec公司)成像。Image-Pro Plus 6.0软件行半定量分析。目的基因的相对表达量以目的条带积分光密度(IOD)值与内参照条带IOD值的比值表示。
1.4 免疫组织化学染色法
采用催化放大法(Catalyzed signal ampli fi cation,CSA)。①标本组织常规石蜡包埋切片,切片脱蜡、水化;②放入 0.3%的过氧化氢(H2O2)溶液阻断内源性过氧化物酶,室温30min; ③0.3% Triton X100室温30min,以增加细胞的通透性;④滴加小鼠抗人Slex单克隆抗体溶液(一抗),4℃过夜;滴加生物素标记的IgG抗体(二抗)室温孵育2h;⑤0.05%二氨基联苯胺(DAB)显色8~12min。苏木素衬染,蓝化。常规封片。以非特异性小鼠IgG代替一抗作为阴性对照。结果判断:Slex阳性信号为细胞胞膜和(或)细胞胞质内出现棕黄色或棕褐色染色颗粒。根据免疫组织化学染色的每例切片中显色有无及显色癌细胞的比例判定阳性或阴性结果:凡无阳性染色细胞或阳性染色细胞数<5%者为阴性(-),阳性染色细胞≥5% 者为阳性(+)。
1.5 统计学分析
采用SPSS12.0统计学软件,α1,3 FUCT Ⅶ基因mRNA表达所得数据以±s表示,两组均数的比较采用t检验,两组以上均数的比较采用单因素方差分析。对Slex表达的数据进行χ2检验,P<0.05为差异有统计学意义。
2 结 果
2.1 α1,3 FUCT Ⅶ基因mRNA在非小细胞肺癌组织中的表达
α1,3 FUCT Ⅶ基因mRNA在腺癌组织表达最强(0.79±0.19),在鳞癌组织中表达(0.51±0.12),差异有统计学意义(P*<0.05)。在癌旁正常肺组织表达量为(0.45±0.11),与腺癌比较差异有统计学意义(P*<0.05),与鳞癌比较差异无显著性(P#>0.05)。此外,α1,3 FUCT Ⅶ基因mRNA表达与年龄无关(P>0.05),与TNM分期、分化程度、淋巴结转移、组织学类型及性别有关(P<0.05),见表1和图1。

图1 α1,3 FUCT ⅦmRNA在癌旁正常组织及非小细胞肺癌组织中的表达

表1 α1,3 FUCT ⅦmRNA的表达与肺癌临床和病理学特征的关系
2.2 Slex的表达
66例肺癌组织中有50例Slex的表达阳性,阳性率为75.7%。癌旁正常肺组织20例Slex的表达均为阴性,差异有统计学意义(P<0.05)。Slex的表达在腺癌阳性率为86%(31/36),鳞癌为63.3%(19/30),差异有统计学意义(P*<0.05),见图2。Slex的表达与年龄无关(P>0.05),与TNM分期、分化程度、淋巴结转移、组织学类型及性别有关。Slex的表达与肺癌临床和病理学特征的关系见表2。
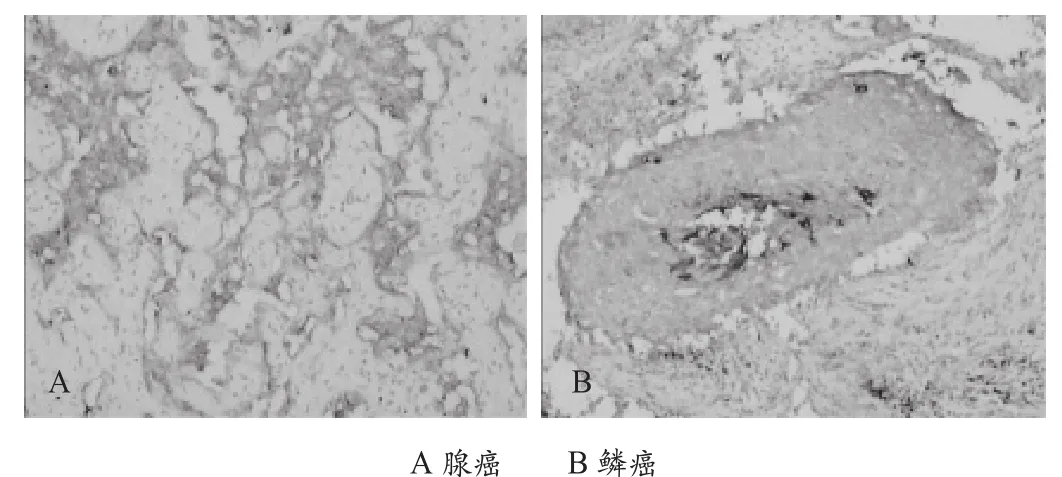
图2 Slex在非小细胞肺癌中的表达(×100)
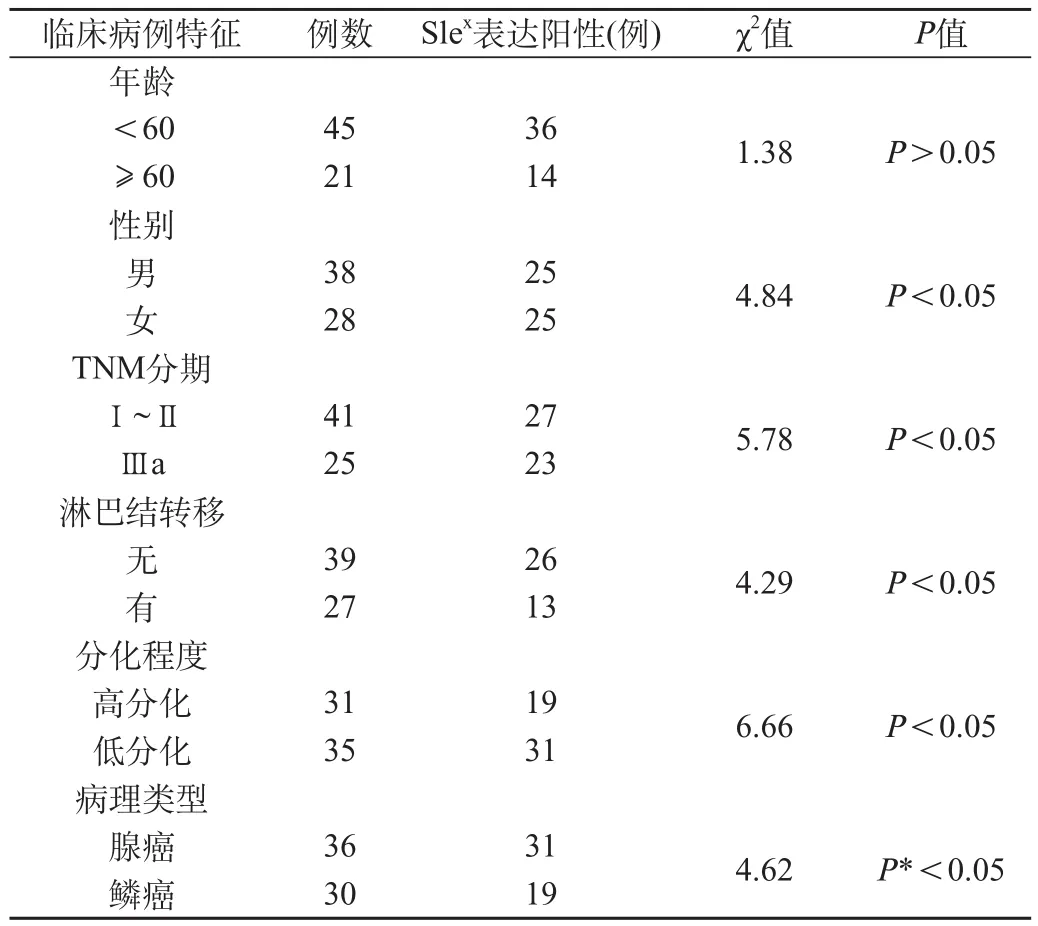
表2 Slex的表达与肺癌临床和病理学特征的关系
3 讨 论
FUCT是一类负责将GDP岩藻糖中的岩藻糖基(Fucose)转移至糖链上N乙酰氨基乳糖单链(GlcNAc)上并以α岩藻糖苷键相连接的酶。其主要作用底物是糖蛋白的O糖或乳糖系、新乳糖系的糖脂,也可以是N糖链,产物主要是含有α岩藻糖基的血型抗原,如ABO血型抗原和Lewis糖链抗原(Le)。FUCT Ⅶ是岩藻糖基转移酶(fucosyltransferase,FUCT)的一个亚型,它是Ⅱ类跨膜拓扑结构,N端有14个氨基酸位于细胞质,接着为19个氨基酸的疏水跨膜区以及位于高尔基体内的309个氨基酸催化结构域。1994年Sasaki等[4]在THP-C细胞cDNA文库中克隆出来的。其cDNA全长为1.7kb,开放阅读框架编码342个氨基酸,相对分子质量为39.2×103,基因定位在染色体9q34.3。目前的研究发现,岩藻糖基化参与多种生物学过程,包括组织生长、血管生成、细胞增殖、黏附、侵袭和肿瘤转移。至少有18种人基因疾病起因于N-糖基化蛋白改变[5-7]。Proro Hamon等[8]的研究显示,高亲和力的选择素和其配体的相互作用需要core2基础的0聚糖,它的合成需要岩藻糖转移酶Ⅶ(FUCT Ⅶ)和磺基转移酶,以core1前提为底物进行修饰,最终形成侧链为Slex的core2基础的0聚糖。而FUCT Ⅶ及其产物Slex对肝癌细胞H7721的转移能力起至关重要的作用[9]。
Slex(又称为唾液酸化的路易斯X,Sialyl Lewis-X)是表达在胚胎组织及肿瘤细胞表面糖脂及糖蛋白上的含有寡聚糖的一组碳水化合物抗原,为细胞黏附分子P选择素所识别的配基。其与P选择素特异性结合以帮助癌细胞穿过内皮细胞基底膜,与恶性肿瘤的恶化和转移呈正相关。早在1999年,陈继华等[10]研究发现肺癌组织中除了Slex抗原以外,可能还存在有Lex及SDlex。其中,SDlex与恶性肿瘤转移的关系要比Slex及Lex更密切。目前Hakomori[11]研究提出癌细胞表达糖链的两种机制为:①正常细胞本来就必须表达的糖链,在癌细胞则出现糖链表达机制的障碍,称为“糖链不全”。②新糖链的合成,指的是牵涉糖类决定子合成的肿瘤相关基因。如通过实验表明唾液酸6-硫代Lex是Slex的N-乙酰葡萄糖胺(GlcNac)基的第6位上有硫酸基结合结构,在正常的大肠黏膜上表达较多,但是随着恶性程度的逐渐增加,该N-乙酰葡萄糖胺(GlcNac)基的第6位硫酸基修饰能力下降,唾液酸6-硫代Lex的表达逐渐消失,Slex增加。
我们的研究发现,α1,3 FUCT Ⅶ基因mRNA在腺癌组织表达最强(0.79±0.19),在鳞癌组织中表达(0.51±0.12),差异有统计学意义(P*<0.05),见表1。Slex的表达在腺癌阳性率为86%(31/36),鳞癌为63.3%(19/30),差异有统计学意义(P*<0.05),见表2。而且α1,3 FUCT Ⅶ基因mRNA及Slex表达与TNM分期、分化程度、淋巴结转移、组织学类型及性别有关,与年龄无关。表明α1,3 FUCT Ⅶ基因mRNA及Slex表达可能与非小细胞肺癌的发生及转移相关,其检测可作为判断患者肺癌转移一项重要参考指标。而Slex可能是在非小细胞肺癌中表达的主要的Lewis抗原,其在非小细胞肺癌中的高表达与肿瘤的侵袭转移能力有关。
众所周知,α岩藻糖转移酶大家族目前已发现有9种亚型,其中FUCT-III(Lewis血型,Le)主要分布在肾、胆囊、及乳汁中。FUCT-IV(类髓型)产物为Lex及VIM-2,存在于5~10周胚胎各组织,在成人仅在髓系组织及大脑中表达。FUCT-VI(血浆型)主要存在于肝脏和血浆,其产物为Lex和Slex。其中FUCT-VI催化合成Slex的能力可以是FUCT Ⅶ的1.5倍[12]。FUCT Ⅶ(白细胞型)在白细胞中表达,只能以唾液酸化的糖链为底物,合成Slex[13]。前期的研究发现,FUCT Ⅶ的活性可以直接影响Slex的表达量并进而影响肝癌细胞的黏附,趋化性迁移和侵袭等转移表型[14]。用佛波脂(TPA,一种促癌剂,也是PKC蛋白激酶激活剂)处理Jurkat细胞观察FUCT Ⅶ及其产物Slex表达的改变,结果发现处理后的细胞增加FUCT Ⅶ的转录及其产物Slex的表达,并能诱导合成E-选择素配体合成[15]。我们的研究中发现,在腺癌组织,α1,3 FUCT Ⅶ基因mRNA表达量为(0.79±0.19),Slex表达阳性率为86%(31/36),均较强,表明在腺癌组织α1,3 FUCT Ⅶ可能是催化合成Slex抗原的主要FUCT。在鳞癌组织,α1,3 FUCT Ⅶ基因mRNA表达较弱(0.51±0.12),且与癌旁正常肺组织表达无显著性差异(P#>0.05),但Slex呈较强表达(阳性率为63.3%,19/30),表明在鳞癌组织中可能存在其他亚型的FUCT,以催化合成Slex。
然而,我们的实验仅限于对非小细胞肺癌组织标本中α1,3 FUCTⅦ基因mRNA及Slex进行研究,验证其与临床病理因素的关系,至于α1,3 FUCT ⅦmRNA及Slex在肺癌中的作用机制尚未涉及,肺癌组织中是否还存在有其它的Lewis抗原,它们与α1,3 FUCT ⅦmRNA关系如何尚不清楚。因此,我们进一步需要研究在肺癌组织中Lewis抗原与其合成酶FUCT之间的关系,并结合相关细胞系及临床结果来证实这种Lewis抗原及相关的糖链加工酶是否能成为肿瘤转移的新的生物学指标和防止肿瘤转移可能的治疗靶点。
[1]Paganuzzi M,Bobbio B,Marroni P,et al.Prognostic role of serum sialyl Lewisx (CD15s)in colorectal cancer[J].Oncology,2003,65(1):52-59.
[2]Sumikura S,Ishigami S,Natsugoe S,et al. Disseminated cancer cells in the blood and expression of sialylated antigen in gastric cancer[J].Cancer letters,2003,200(1):77-83.
[3]Kenji N,Makoto S,Kazuko H,et al.Sialosyl-Le(x)expression de fi nes invasive and metastatic properties of bladder carcinoma[J].Cancer,2002,94(3): 673-685.
[4]Sasaki K,Kurata K,Funayama K,et al.Expression cloning of a novel alpha1,3-fucosyltransferase that is involved in biosynthesis of the sialyl lewis X carbonhydrate determinants in leukocytes[J].J Biol Chem,1994,269(20):14730-14737.
[5]Komrower JJ. Lecture.Congenital disorder of glycosylayion (CDG):it' s all in it[J].J Inherit Metab Dis,2003,26 (2/3):99-118.
[6]Marquardt T,Denecke J.Congenital disorder of glycosylation: review of their molecular bases .Clinical presentations and speci fi c therapies[J]. Eur J Pediatr,2003,162(6):359-379.
[7]Freeze HH,Aebi M.Altered glycan structure:the molecular basis of congenital disorders of glycosylation[J].Curr Opin Struct Biol,2005,15(5):490-498.
[8]Prorok Hamon M,Notel F,Mathiew S,et al.N-glycans of Core2β(1,6)-N-acetylglucosaminy-ltransferase-I(C2GnT-1)but not those of α(1.3)-fucosyltransferase-Ⅶ(FUCTⅦ)are Required for the Synthesis of Function P-Selectin Glycoprotein Ligand(PSGL-1)Effects on P-,L-,and E-selectin Binding[J].Biochem J,2005,391(Pt 3):491-502.
[9]Hiraiwa N,Dohi T,Kawakami-kimura N,et al.Suppression of sialyl Lewis X expression and E-selectin-mediated cell adhesion in cultured human lymphoid cells by tranfection of antisense cDNA of an alphal-3 fucosyltranferase (FUCTⅦ)[J].J Biol Chem,1996,2 71(49):31556-311561.
[10]陈继华,刘飞,陈惠黎.肺癌组织中岩藻糖化链结构免疫组化研究[J].实验生物学报,1999,32(1):7-12.
[11]Hakomori S.Tumor-associated glycolipid antabolism and organization[J].Chem Phys Lipids,42(1/3):209-233.
[12]Togayachi A,Kudo Y,Ikehara Y,et al.Up-regulation of Lewis enzyme(α1.3- FUCTIII)and plasma-typeα1.3-fucosyltransferase (FUCTVI)expression of sialyl lewis X antigen in non-small cell lung cancer [J].Int J Cancer,1999,84(1):70-83.
[13]Narimats U.Human fucosyltransferase:tissue distribution of blood group antigens,cancer-associated antigen and fucosltransferase[J].Tanpakushitsu Kakusan Koso,1998,43(16):2394-2403.
[14]Liu F,Qi HL,Chen HL.Regulation of differentiation and proliferationinduce on Lewis antigens,αfucosyltransferase and metastatic potential in hepatocarcinoma cell[J].Br J Cancer,2001,84(11):1556-1563.
[15]Knibbs RN,Craig RA,Natsuka S,et al.The fucosyltransferase FUCTⅦ regulates E-selectin ligand synthesis in human T cells [J].J Cell Biol,1996,133(4):911-920.
猜你喜欢
食品与发酵工业(2022年7期)2022-04-18
云南医药(2021年3期)2021-07-21
食品研究与开发(2020年16期)2020-08-11
天然产物研究与开发(2019年4期)2019-04-27
中成药(2018年7期)2018-08-04
中华老年口腔医学杂志(2016年2期)2017-01-15
中国实验动物学报(2015年3期)2015-12-16
中国当代医药(2015年17期)2015-03-01
癌变·畸变·突变(2015年3期)2015-02-27
中华皮肤科杂志(2014年3期)2014-12-19
