
经前舒颗粒对郁怒反应模型大鼠海马脑区基因表达谱的影响
2011-05-29 12:43郭英慧徐凯勇宋春红乔明琦
中国药理学通报 2011年9期
郭英慧,高 杰,徐凯勇,宋春红,乔明琦
(山东中医药大学1.基础医学院、2.实验动物中心,山东 济南 250355)
怒是影响人体健康的重要情志因素,其在疾病发生、发展中的作用和机制已成为研究的热点之一。前期研究表明:愤怒和郁怒是人们发怒的两种表达方式,分别对焦虑和抑郁及其相关症状产生起重要作用[1]。临床回顾性流行病学调查结果显示:郁怒与抑郁症、心脑血管疾病、癌症等疾病的发生、发展密切相关[2-3]。郁怒的产生及其诱发的情志病证的发病机制是一个复杂的过程,涉及神经、内分泌、免疫等多个系统,具体作用机制尚不清楚[4]。用现代科学技术研究情志病证,这是当今中西医研究的重要课题。基因芯片技术以其快速、高效、高通量的特点已成为研究复杂疾病发生、发展的分子机制的有力工具之一。因此,本研究拟借助基因芯片技术,研究调肝方药经前舒颗粒对郁怒反应模型大鼠海马基因表达谱干预作用并初步探讨其作用机制。
1 材料与方法
1.1材料
1.1.1实验动物健康♂Wistar大鼠,体质量180~220 g,SPF级,由山东中医药大学实验动物中心提供,许可证号 SCXK(鲁)20050015。
1.1.2主要试剂和仪器TRIzol试剂盒(美国Invitrogen公司);经前舒颗粒(主要成分:白芍、当归、柴胡、白术、丹皮、香附、甘草、郁金等药组成。秦皇岛市山海关制药厂生产,生产批号:Z20053587);RT-PCR反应试剂包括 M-MLVReverse、dNTP Mixture、rTaq、DNaseⅠ(大连TaKaRa生物公司)。引物合成由济南博亚生物工程技术服务有限公司完成。大鼠表达谱芯片Affymetrix Rat Genome 230 2.0 Array(上海生物芯片有限公司),A260/A280测定仪,Spectrophotometer ND21000(NanoDrop);Agilent扫描仪。
1.2方法
1.2.1大鼠造模、分组及行为学评价依据文献[5]制备愤怒、郁怒反应大鼠模型。首先筛选旷场实验、糖水偏好得分相近大鼠进入实验,社会隔离加居住入侵造模刺激1周,依据攻击行为区分出愤怒、郁怒组大鼠模型,挑选出模型组中属郁怒反应大鼠,继而将郁模组分为继续造模组和模型给药组。即实验设置正常组、郁怒模型组(简称郁模组)、经前舒给药组(简称郁药组),每组10只。正常组、郁模组(造模同时)灌胃给予灭菌饮用水,郁药组造模同时灌胃给予经前舒颗粒,每天上午9:00灌胃给药1次,给药量0.1 ml·g-1体质量(相当于人临床8倍剂量[6],浓度1 ∶1,即1 ml药液含原生药1 g),连续给药7 d。采用旷场实验和糖水偏好实验对模型进行评价[5]。
1.2.2大鼠脑组织取材评价实验后,每只大鼠迅速断头、开颅、分离脑置于冰上培养皿中,无菌操作于冰上迅速剥离出海马组织,分别放入1.5 ml离心管,置于液氮中冷冻保存。
1.2.3大鼠基因芯片的检测
1.2.3.1组织总RNA提取 取冻存的大鼠海马组织,按组别用组织匀浆机分别混合均匀,TRIzol试剂抽提海马组织的总RNA,进行紫外分析及电泳检测。QIAGEN RNeasy Kit纯化总RNA,测定其浓度后,-80℃保存。
1.2.3.2探针制备、芯片杂交扫描及数据提取 提取的RNA在干冰的保护下交上海生物芯片有限公司,待确认质量后参考Affymetrix基因芯片实验操作手册进行反转录成cDNA,并用荧光Cy3-dCTP标记郁模组和郁药组总RNA,用Cy5-dCTP标记正常组的总RNA。然后用定量的Cy3和Cy5标记的探针,进行芯片杂交、洗涤、干燥。芯片结果用 Affymetrix扫描仪进行扫描,GCOS1.4软件读取数据[7]。
1.2.3.3差异表达基因筛选与生物信息学分析 由上海生物芯片有限公司对扫描数据进行数字化处理和数据统计;差异基因筛选标准为Log-ratio≥0.8为上调基因,Log-ratio≤0.8为下调基因,P<0.5。数据分析软件采用博奥生物公司的GO分子功能注释系统,将差异基因分子功能、生物过程、细胞成分进行归类,并利用公共数据库(BioCarta、KEGG、GenMApp、NCBI Gene Bank等)对差异表达基因涉及的调控路径进行检索分析[7],同时参阅大量相关文献。
1.2.4RT-PCR实验分别取3只正常组、郁模组和郁药组大鼠的海马组织,提取RNA后,逆转录生成cDNA,采用Primer 3软件,选取基因芯片检测结果中Htr3b和GABABR2基因设计引物,以β-actin为内参,进行 PCR 验证[8]。
1.2.5统计学方法采用SPSS 13.0统计软件,各项检测数据以±s记录。采用t检验和χ2检验进行统计学处理。
2 结果
2.1旷场实验及糖水偏好实验由Fig 1可见:造模刺激1周后郁怒大鼠模型旷场实验得分上升(P<0.05),而糖水偏好系数则下降(P<0.05);药物干预后上述指标均得到一定程度恢复,但与郁模组比较差异无显著性(P>0.05)。

Fig 1 Open-field test(A)and sucrose preference test(B)results of control,anger-in model,Jingqianshu treatment rats(¯x ± s,n=10)
2.2组织样品RNA检测和芯片检测质量判定所提取总RNA经紫外分光光度计检测,OD260nm/OD280nm比值介于1.9~2.1;经琼脂糖凝胶电泳分析,28 S、18 S两条带清晰,且无拖尾现象,提示RNA完整性较好,总RNA浓度、纯度及完整性符合实验要求(Fig 2)。基因芯片检测结果显示,Cy3/Cy5比值正常,芯片检出率在0.99以上,均一性良好,符合质控标准,信号强度达到要求。
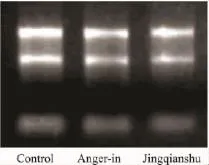
Fig 2 RNA electrophoresis results of control,anger-in model,Jingqianshu treatment.
2.3差异表达基因的功能分布及其调控路径
2.3.1各组差异表达基因的功能分布按Gene Ontology功能分类标准对差异表达基因进行检索分类,由于某些差异表达基因并非仅有一个功能或仅通过一条调控路径。经查基因库,造模组与正常组比较共检出63个表达上调基因和153个表达下调基因,涉及蛋白结合活性等分子功能的基因259个,有关信号转导等生物过程的基因281个,涉及膜组分等细胞成分的基因204个。郁药组与郁模组比较共检出127个不同差异表达基因,其中郁模组/正常组上调而郁药组/正常组基因下调基因有55个,郁模组/正常组下调而郁药组/正常组基因上调基因有72个,主要有各类激素受体、信号转导蛋白、代谢酶、离子通道蛋白、转运蛋白、细胞因子受体等。选取部分郁药组与郁模组比较差异表达基因进行分子功能注释,见Tab 1。
2.3.2各组差异表达基因的调控路径利用KEGG等数据库对差异表达基因涉及的调控路径进行检索分析发现,多基因可通过一条调控路径。造模组与正常组比较发现差异基因共涉及124条调控路径,最多者6个基因/路径,为神经活化配体-受体交互作用路径;其次为4个基因/路径和3基因/路径,为甘油磷脂代谢路径和促分裂原活化蛋白激酶路径。另外还有一基因通过多条调控路径情况,最多的为10条路径/基因,分别是核糖体蛋白S6激酶(Rps6kb1)和磷脂酶Ce1(Plce1)。郁药组与郁模组比较发现差异基因与80条调控路径有关,通过调控基因最多的为4基因/路径,为神经活化配体-受体交互作用路径和离子转运调控路径。同时Rps6kb1和Plce1基因也是涉及调控路径最多的基因,为10条路径/基因。
2.4RT-PCR验证基因芯片检测结果选择基因芯片结果提示的有差异表达的Htr3b和GABABR2基因进行RT-PCR实验,Htr3b和GABABR2基因表达结果与基因芯片数据基本一致(Fig 3),说明基因芯片检测结果是可靠的。
3 讨论

Tab 1 Selected genes differentially expressed in the hippocampus from Jingqianshu treatment and anger-in model rat

Fig 3 Electrophoresis results of Htr3b(A)and GABABR2(B)RT-PCR test from rat hippocampus
居住入侵及社会隔离是社会压力应激领域经典造模方法,旷场实验和糖水偏好为动物行为学领域经典评价方法,国内外医学界应用以上方法已成功制备出抑郁症、经前烦躁易怒等多种情绪类疾病动物模型[5]。本实验采用居住入侵结合社会隔离的方法制备愤怒、郁怒情绪大鼠模型[8]。与正常组相比,郁模组大鼠旷场实验总分明显上升,而糖水偏好系数显著下降,初步判定模型制备成功,用药干预后负性情绪能得到一定程度地改善,但郁药组与郁模组比较差异未见显著性(P>0.05),这与模型用药时间较短有关,但仍能很大程度上说明调肝方药经前舒颗粒能够纠正郁怒反应模型大鼠行为学变化。
中药复方是以多组分、多靶点发挥效应,其对基因的调控可能是通过群体基因表达实现。基因芯片具有大批量、同时筛选和整体信息处理的性质,本研究利用此特点对经前舒颗粒干预郁怒大鼠海马脑区基因表达谱进行初步分析,了解该方应用后的一些基因反应模式,以期从基因水平上探索其可能的药物靶点及作用机制。结果显示该方可使郁模组海马组织中的较多基因产生差异性表达,提示方中的药物作用靶点较多,是通过整体调节基因的高/低表达发挥药物的作用。在上调或下调的一些已知基因中,其功能与保护神经元、抑制细胞凋亡、信号转导突触传递保护、稳定Ca2+通道、拮抗缺氧等有关,这些基因或是药物作用靶点或是药物信号传导通路上的某个环节。差异表达基因多属于神经活化配体—受体交互作用、甘油磷脂代谢和促分裂原活化蛋白激酶等调控路径,说明这些调控路径与经前舒颗粒可能的药物干预分子机制密切相关,这可能是该方干预郁怒情绪反应作用的基础。
举以下几个基因为例讨论:① Htr3b(NM_022189)和催乳素受体Prlr(NM_001034111)基因,有报道Htr3b受体参与调控中枢性恶心、呕吐和应激性肠综合征等过程,基因多态性与女性严重抑郁症密切相关。Prlr受体在泌乳、生长发育、内分泌行为、免疫调节等生理过程中发挥重要生理功能[9-10]。经前舒颗粒可能通过调节这些激素及神经递质受体,进而参与神经—内分泌—免疫系统功能失调调节过程,这与以往报道慢性愤怒应激与神经内分泌免疫功能失调密切相关等研究结果相符[8,11]。② 成纤维细胞生长因子受体 Fgfr2(NM_001109892)和代谢型谷氨酸受体 Grm8(NM_022202),大量研究表明成纤维细胞生长因子bFGF可通过与受体Fgfr2特异性结合,激活酪氨酸激酶,影响谷氨酸受体蛋白、钙结合蛋白等的表达而稳定Ca2+通道,拮抗缺氧、兴奋性氨基酸、自由基等的损害,抑制细胞凋亡的发生、发展,从而保护神经元[12-13]。Grm8(NM_022202)在神经网络的形成、可塑性改变以及学习、记忆等认知的突触传递过程中发挥重要作用,其功能异常可能导致神经系统的损害[14]。经前舒颗粒可致 Fgfr2、Grm8表达上调,可能通过促进抗氧化应激,增加Grm8突触前抑制,减少谷氨酸的释放,对海马神经元起到保护性效应,起到抑制其凋亡作用。③糖皮质激素反应元件结合蛋白Gmeb2(NM_031803),海马是糖皮质受体含量最高的脑区,应激时海马极易受到高浓度的糖皮质激素选择性的攻击,导致海马整体受损,突触可塑性降低,神经元萎缩、丢失,并进一步影响到学习记忆等高级功能[15-16]。经前舒颗粒可上调 Fgfr2、Gmeb2基因表达,推测可能通过拮抗缺氧、自由基的损害,减少游离糖皮质激素含量,从而对应激型海马神经元起保护作用。
郁怒情绪及与其密切相关的情志病证的发病机制非常复杂,本研究虽初步证实经前舒颗粒可通过改变郁怒情绪大鼠海马脑区一些基因反应模式,能够部分纠正模型大鼠行为学变化,但很大部分基因功能还远未完全搞清楚。在后续的研究中,我们将对芯片结果进行进一步验证,并利用相关情志病证动物模型或精神药理学相关研究来佐证基因芯片发现的一些阳性结果,为深入探讨与郁怒情绪密切相关的情志病证的发生和发病机制提供基本依据和研究方向。
[1]Bridewell W B,Chang E C.Distinguishing between anxiety,depression,and hostility:relations to anger-in,anger-out,and anger control[J].Pers Individ Dif,1997,22(4):587-90.
[2]Julkunen J,Gustavsson-Lilius M,Hietanen P.Anger expression,partner support,and quality of life in cancer patients[J].J Psychosom Res,2009,66(3):235-44.
[3]窦学俊,高冬梅,王婧婧,等.护士人群怒表达方式及相关情志病证流行病学调查[J].中医杂志,2009,12:1093-5.
[3]Dou X J,Gao D M,Wang J J,et al.Epidemiological investigation of nurses’emotional diseases related to anger and its expression[J].J Trad Chin Med,2009,12:1093-5.
[4]易正辉,方贻儒,禹顺英.抑郁症脑组织基因差异表达研究进展[J].中国神经精神疾病杂志,2010,36(1):56-9.
[4]Yi Z H,Fang Y R,Yu S Y.The research progress of differential expression genes of depression brain[J].Chin J Nerv Ment Dis,2010,36(1):56-9.
[5]Wei S,Zhang H Y,Gao J,et al.Impact of social isolation and resident intruder stress on aggressive behavior in the male rat[J].Neural Regen Res,2010,5(15):1175-9.
[6]冯 玉,张惠云.经前舒颗粒对经前期综合征肝气郁证大鼠大脑皮层雌激素受体表达的影响[J].中药药理与临床,2010,26(2):71-3.
[6]Feng Y,Zhang H Y.Effects of Jingqianshu granule on the expression of estrogen receptor(ER)of PMS model rats with liver-qi stagnation[J].Pharmacol Clin Chin Mat Med,2010,26(2):71-3.
[7]Li R Y,Zhang Q H,Liu Z,et al.Effect of short-term and longterm fasting on transcriptional regulation of metabolic genes in rat tissues[J].Biochem Biophys Res Commun,2006,344(2):562-70.
[8]王杰琼,张惠云.愤怒、郁怒反应模型大鼠海马5-HTR2C的基因表达[J].中国药理学通报,2011,27(3):325-8.
[8]Wang J Q,Zhang H Y.The expression of 5-HTR2C in the hippocampal of the anger-in and anger-out animal model[J].Chin Pharmacol Bull,2011,27(3):325-8.
[9]Férézou I,Cauli B,Hill E L,et al.5-HT3 receptors mediate serotonergic fast synaptic excitation of neocortical vasoactive intestinal peptide/cholecystokinin interneurons[J].J Neurosci,2002,22(17):7389-97.
[10]Krzywkowski K,Davies PA,Feinberg-Zadek PL,et al.High-frequency HTR3B variant associated with major depression dramatically augments the signaling of the human 5-HT3AB receptor[J].Proc Natl Acad Sci USA,2008,105(2):722-7.
[11]詹向红,李 伟,赵君玫,等.慢性愤怒应激对大鼠衰老进程及其神经内分泌免疫机制的影响[J].中华中医药杂志,2010,25(1):111-3.
[11]Zhan X H,Li W,Zhao J M,et al.Effects of chronic anger stress on rats aging process and neuroendocrine-immune mechanism[J].China J Tradi Chin Med Pharma,2010,25(1):111-3.
[12]Gaughran F,Payne J,Sedgwick P M,et al.Hippocampal FGF-2 and FGFR1 mRNA expression in major depression,schizophrenia and bipolar disorder[J].Brain Res Bull,2006,70(3):221-7.
[13]Rosenblatt S,Irikura K,Caday CG,et al.Basic fibroblast growth factor dilates rat pial arterioles[J].J Cereb Blood Flow Metab,1994,14(1):70-4.
[14]Nakanishi S,Nakajima Y,Masu M,et al.Glutamate receptors:brain function and signal transduction[J].Brain Res Rev,1998,26(2-3):230-5.
[15]Joel sM,Karst H.Effects of chronic stress on structure and cell function in rat hippocampus and hypothalamus[J].Stress,2004,7(4):221-31.
[16]Zhang L,Kokkonen G,Roth G S.Identification of neuronal programmed cell death in situ in the striatum of normal adult rat brain and its relationship to neuronal death during aging[J].Brain Res,1995,677(19):177-9.
猜你喜欢
作文周刊·小学二年级版(2022年20期)2022-05-05
今日农业(2021年4期)2021-06-09
知识文库(2020年5期)2020-05-11
电子技术与软件工程(2019年17期)2019-10-09
创新作文(小学版)(2019年10期)2019-09-25
电子制作(2019年12期)2019-07-16
中国交通信息化(2018年5期)2018-08-21
电子制作(2018年8期)2018-06-26
中国实用医药(2016年1期)2016-01-11
中国药房(2010年12期)2010-02-12
