
披碱草属8种野生牧草居群穗部形态多样性
2011-07-25 08:52德英穆怀彬刘新亮赵来喜王照兰
草业科学 2011年9期
德英,穆怀彬,刘新亮,赵来喜,王照兰
(1.中国农业科学院草原研究所,内蒙古呼和浩特 010010;2.农业部草原资源与生态重点开放实验室,内蒙古呼和浩特 010010;3.中国农业科学院研究生院,北京 100081)
披碱草属(Elymus)是禾本科小麦族内一个重要的属,我国野生披碱草属牧草种质资源丰富,广泛分布于各地,尤其西、北部地区是披碱草属牧草的重要分布区和多样性分化中心[1]。披碱草属的多数物种为草原和草甸的重要组成成分,许多种类是优良的牧草,饲用价值极高[2]。丰富的遗传变异对于培育高产、优质、高抗的栽培牧草品种非常重要,野生牧草种质是育种家改良现有牧草品种的二级基因库,了解野生牧草种质所蕴涵的遗传多样性对于选育优异品种和种质资源保护具有重要意义[3]。
披碱草属牧草作为麦类作物的近缘属,是麦类作物遗传改良的重要优异基因来源,是优质高产的牧草,也是重要固沙植物,具有重要的利用价值[3]。目前国内外关于披碱草属牧草的形态学性状、染色体、RAPD及SSR标记等手段研究披碱草属牧草遗传多样性的较多,形态多样性是检测遗传变异最传统和常用的方法[4-9],但是在种类、分布上大范围内的研究较少。长期以来,种质资源的分类、鉴定及育种材料的选择通常都是依据表型性状来进行的,其中穗部性状的变异是禾本科物种形态多样性的主要表型性状。披碱草属牧草分布广泛,种类繁多,广义的披碱草属包括近150个种,而在我国,则比较接受狭义披碱草属的概念,即有12余种[10]。本研究对分布于我国的披碱草属8种101个野生牧草居群的14项穗部形态学性状进行评价,分析其表型多样性,旨在为我国披碱草属牧草种质资源的保存利用、开发选育提供理论依据,还为拓宽其品种遗传基础,合理选择亲本及其繁育系统研究奠定基础。
1 材料与方法
1.1 材料试验材料全部为我国野生披碱草属种质材料(表1),共8个种101个居群。其中垂穗披碱
草(Elymusnutans)29个居群,短芒披碱草(E.breviaristatus)4个居群,肥披碱草(E.excelsus)6个居群,黑紫披碱草(E.atratus)3个居群,老芒麦(E.sibiricus)24个居群,麦薲草(E.tangutorum)10个居群,披碱草(E.dahuricus)18个居群,圆柱披碱草(E.cylindricus)7个居群。
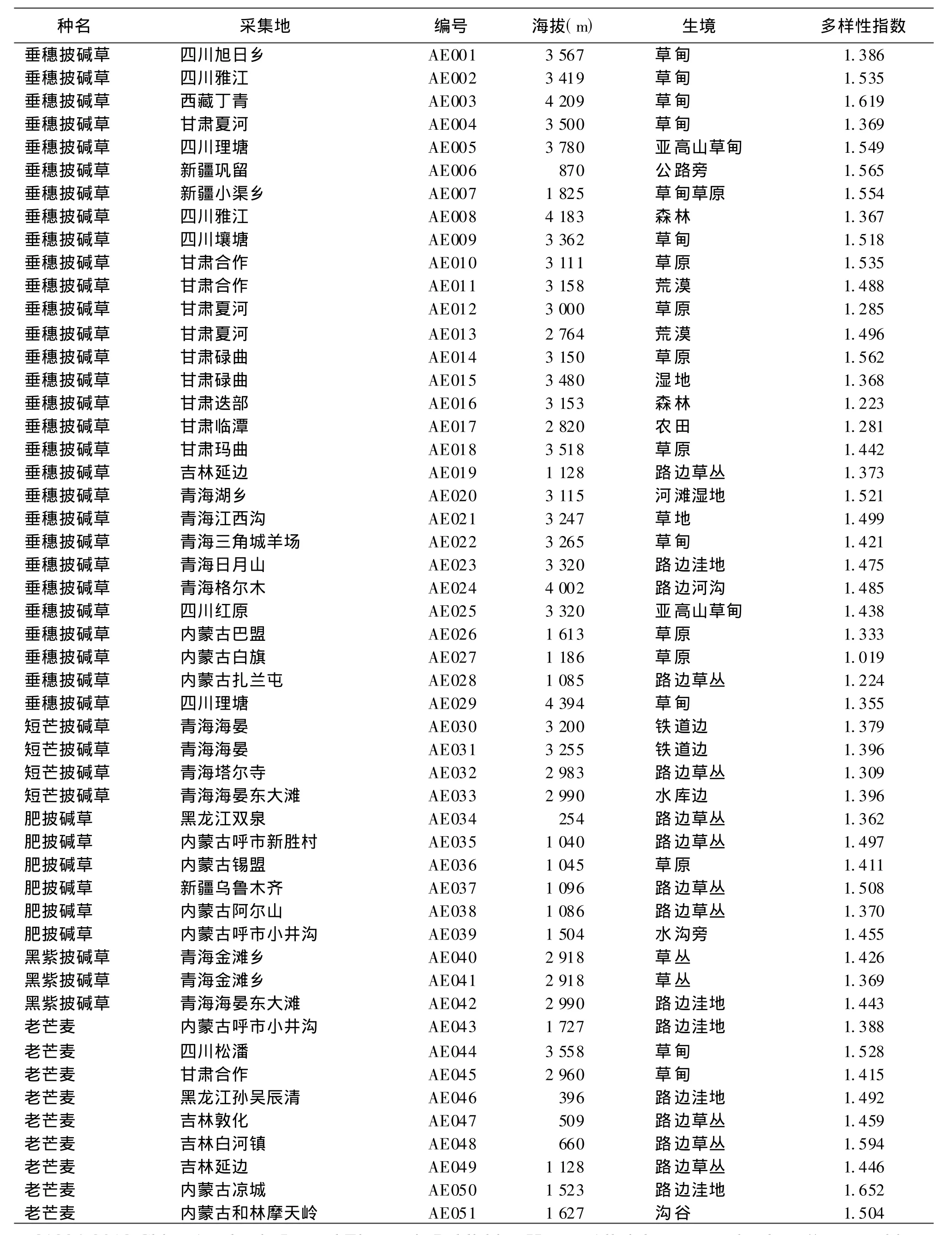
表1 供试材料名录及居群多样性指数

续表1
1.2 试验地概况试验地位于呼和浩特市西南约30余km的土默特左旗沙尔沁乡“农业部沙尔沁牧草资源重点野外科学观测试验站”,处在大青山及和林格尔丘陵之间的广阔平原地区,土壤以淡栗钙土为主 ,海拔 1 055 m,40°36′N,111°45′E,为干旱、半干旱温带大陆性气候,冬季长而寒冷,夏季短而炎热,寒暑变化剧烈,降水量少,蒸发量大,气候干燥,无霜期短。
1.3 试验设计2009年4月15日,选用大田的土壤,过筛,去掉石块、杂质,土∶腐熟羊粪的比例是5∶1,用育苗盘装土,均匀的将种子撒在盘中,再轻轻的用土覆盖,然后用喷头浇透,置于温室中。6月13日,单株移栽到试验地,采用随机设计,各材料的小区面积为2 m×5 m。
1.4 田间调查2010年7-9月,在植株成熟期测定,包括穗长、穗宽、小穗长、小穗宽、小穗数、每穗轴小花数、第一外稃长、第一内稃长、第一外稃芒长、第一颖长、第一颖芒长、第一颖宽、旗叶与穗基部长度、主穗轴第一节间长共计14项指标,其中旗叶与穗基部长度、主穗轴第一节间长用直尺测量,其余均用游标卡尺测量,每个指标均为10个重复,具体测量方法参照《老芒麦种质资源描述规范和数据标准》[11]。
1.5 数据处理试验数据采用Excel和SPSS 11.5软件进行多样性指数计算、方差分析。多样性指数的计算采用Shannon-weaver指数。

式中,Pi为某性状第i个代码值出现的概率[12]。数量性状如穗长、穗宽、小穗长等进行 10级分类,1级<x-2s,10级≥x+2s,中间每级间差0.5s,s为标准差。
居群总多样性指数为同一种所有居群一起计算得出的多样性指数;居群平均多样性指数为同一种不同居群多样性指数的平均值。
2 结果与分析
2.1 披碱草属8种牧草种质资源的形态性状变异分析以居群为单位,统计披碱草属101个居群植株的14项穗部性状,并按种进行整体平均统计,计算平均数、标准差、变异系数。结果表明,披碱草属8种牧草种间存在差异(表2)。穗长平均值以肥披碱草最大;第二为披碱草、老芒麦及黑紫披碱草,这3种无显著差异(P>0.05);第三是垂穗披碱草;第四为麦薲草和圆柱披碱草,二者间无显著差异(P>0.05),短芒披碱草最小,其分为5个显著差异性组合(P<0.05)。穗宽平均值肥披碱草最大,黑紫披碱草最小,分为4个显著差异性组合(P<0.05);小穗长平均值肥披碱草最大,短芒披碱草最小,分为5个显著差异性组合(P<0.05);小穗宽平均值肥披碱草最大,圆柱披碱草最小,分为4个显著差异性组合(P<0.05);小穗数平均值肥披碱草最大,垂穗披碱草最小,分为5个显著差异性组合(P<0.05);每穗轴小花数平均值肥披碱草最大,短芒披碱草最小,分为4个显著差异性组合(P<0.05);第一外稃长平均值老芒麦最大,黑紫披碱草最小,分为5个显著差异性组合(P<0.05);第一内稃长平均值老芒麦最大,圆柱披碱草最小,分为4个显著差异性组合(P<0.05);第一外稃芒长平均值肥披碱草最大,短芒披碱草最小,分为6个显著差异性组合(P<0.05);第一颖长平均值肥披碱草最大,垂穗披碱草最小,分为6个显著差异性组合(P<0.05);第一颖芒长平均值老芒麦最大,黑紫披碱草最小,分为5个显著差异性组合(P<0.05);第一颖宽平均值披碱草最大,垂穗披碱草最小,分为3个显著差异性组合(P<0.05);旗叶与穗基部长度平均值麦薲草最大,老芒麦最小,分为5个显著差异性组合(P<0.05);主穗轴第一节间长平均值肥披碱草最大,圆柱披碱草最小,分为4个显著差异性组合(P<0.05)。
由此可知,8种披碱草属牧草中,肥披碱草的14项穗部形态学性状中有9个性状(穗长、穗宽、小穗长、小穗宽、小穗数、每穗轴小花数、第一外稃芒长、第一颖长、主穗轴第一节间长)为最大值;黑紫披碱草的穗宽、第一外稃长、第一颖芒长3个性状为最小值。
14项穗部性状中(表3),变异系数大小依次为第一颖芒长>旗叶与穗基部长度>第一颖长>主穗轴第一节间长>第一颖宽>第一外稃芒长>穗宽>小穗数>每穗轴小花数>小穗宽>穗长>小穗长>第一外稃长>第一内稃长。
2.2 披碱草属8种牧草遗传多样性分析
2.2.1披碱草属8种牧草野生居群总多样性分析披碱草属8种牧草居群总多样性指数为垂穗披碱草>老芒麦=披碱草>麦薲草>肥披碱草>短芒披碱草>圆柱披碱草>黑紫披碱草;总平均多样性指数为1.826(表4)。由此可见,垂穗披碱草、老芒麦、披碱草和麦薲草4种披碱草属牧草的居群总多样性较其他披碱草属牧草丰富。
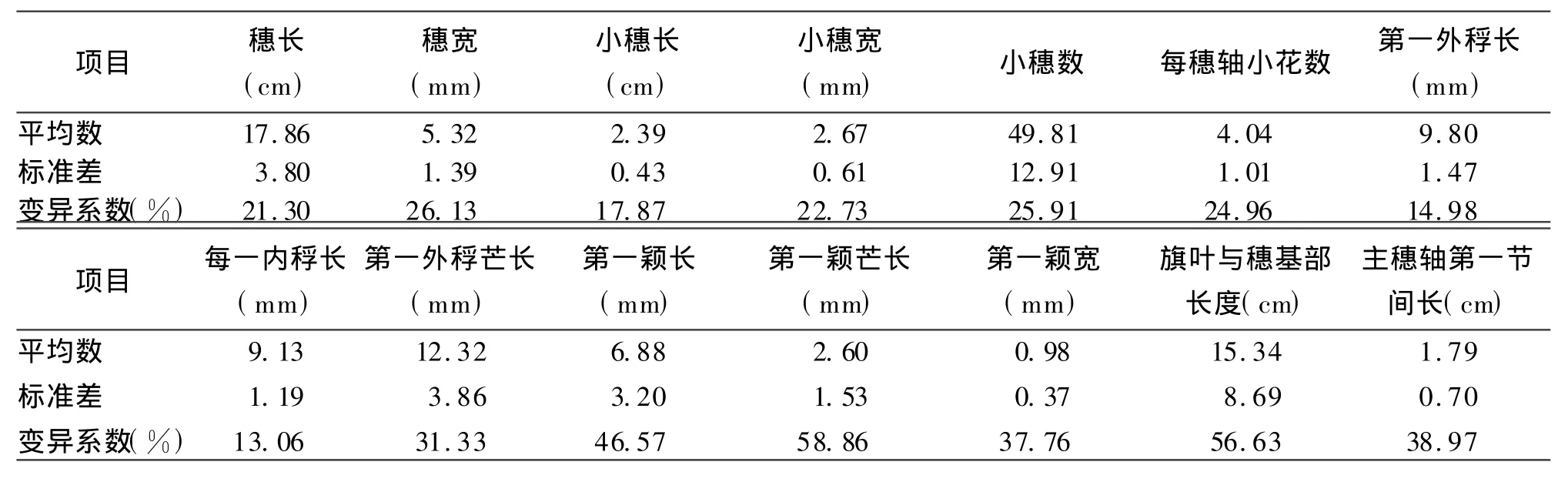
表3 14项穗部形态学性状

表4 居群总多样性指数
2.2.2披碱草属8种牧草野生居群平均多样性分析根据表1中各居群的多样性指数计算披碱草平均多样性指数。8种披碱草属牧草居群平均多样性指数为老芒麦>肥披碱草>垂穗披碱草>披碱草>黑紫披碱草>短芒披碱草>圆柱披碱草>麦薲草;所有居群的平均多样性指数为1.372(表5)。
由此可见,居群多样性相对较高的有老芒麦、肥披碱草、垂穗披碱草和披碱草,说明这4种披碱草属牧草居群存在较高的多样性。
2.2.3披碱草属8种牧草野生居群表型分化系数分析 8种披碱草属牧草居群表型分化系数为麦薲草>圆柱披碱草>垂穗披碱草>披碱草>老芒麦>肥披碱草>短芒披碱草>黑紫披碱草;平均表型分化系数为24.86%(表6)。由此可见,麦薲草、圆柱披碱草、垂穗披碱草、披碱草存在相对较高的表型分化,说明导致它们多样性不仅是由于居群多样性,而且居群间多样性也起着相对较大的作用。
综上可见,披碱草属8种牧草的多样性丰富是由居群和居群间多样性引起的,不同物种其多样性二者所占的比例各不相同。
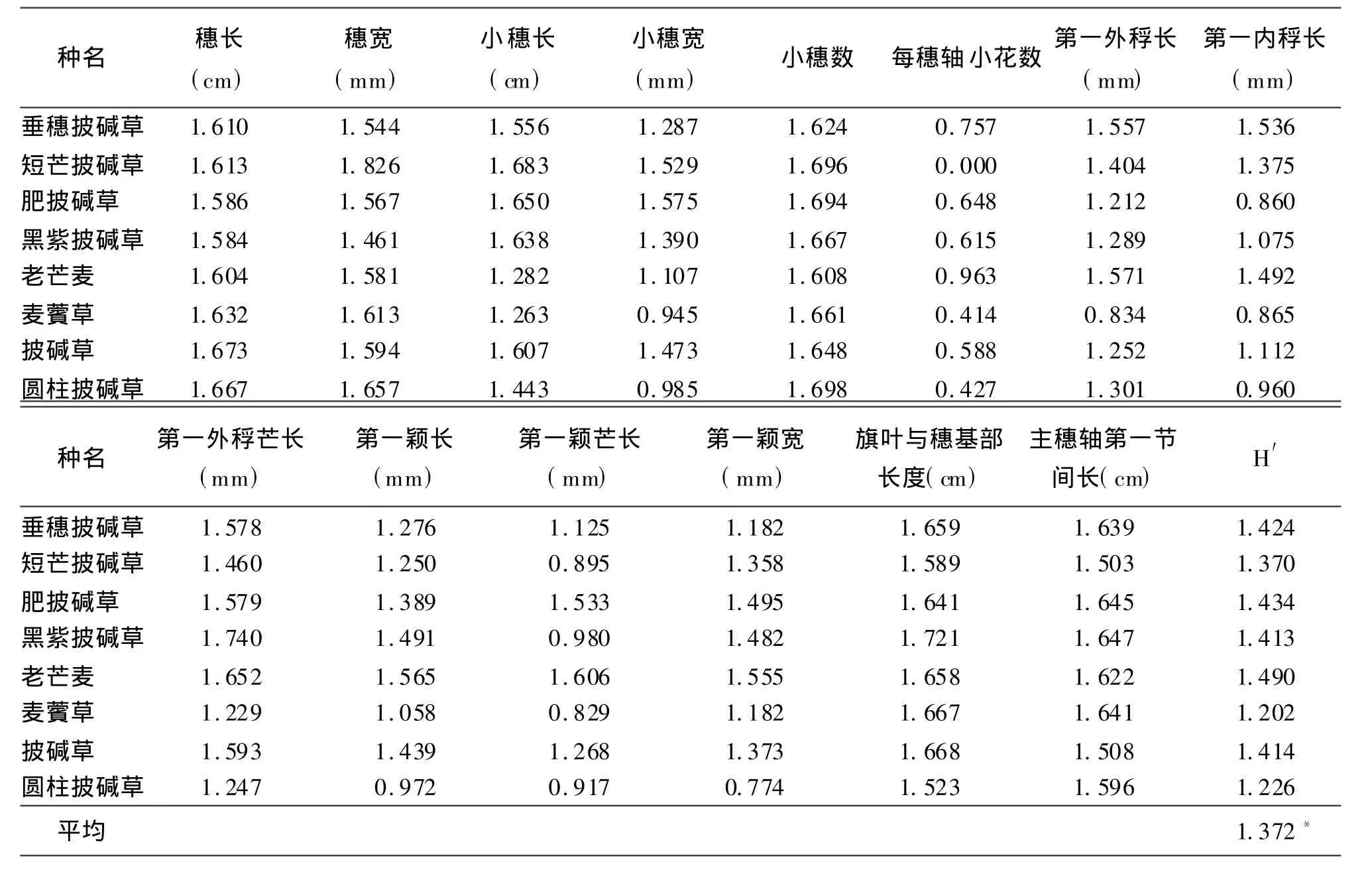
表5 居群平均多样性指数
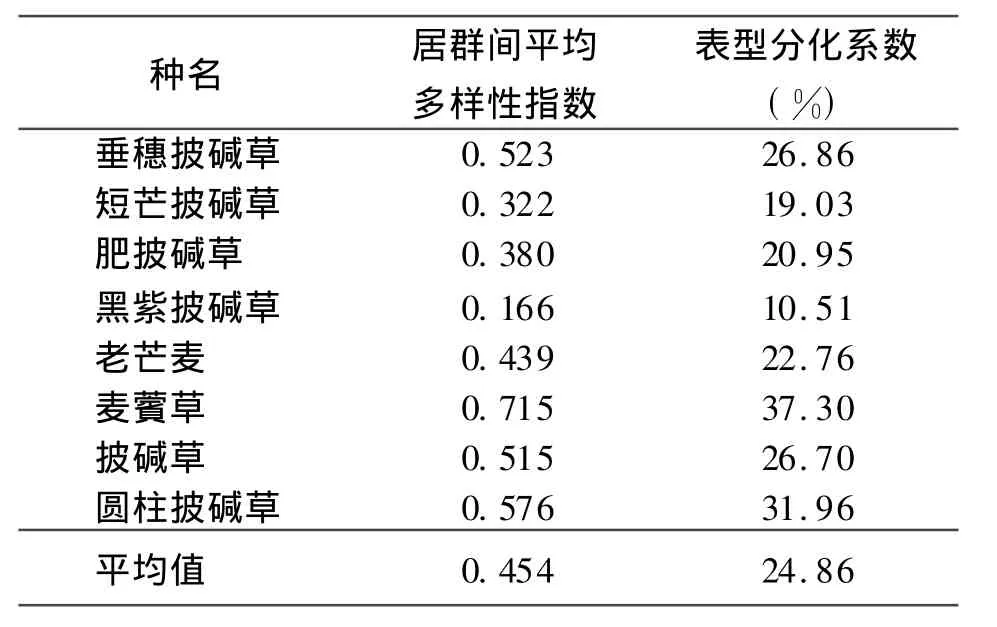
表6 居群间多样性与表型分化系数
3 讨论
形态学标记研究材料的外部形态,受到基因和所处生态环境的共同影响,将所有的材料种植在同一环境下,减少环境对研究材料的影响,对材料间受基因控制的遗传差异进行研究。对处于不同生态环境下的同种材料的形态学研究,可以了解环境对基因表达的影响[13]。我国披碱草属8种牧草101个野生居群的14项穗部性状,其平均数、标准差、变异系数均存在较大的变异,且变异程度各不相同,说明我国8种披碱草属牧草野生居群的各个穗部性状均存在着较丰富的遗传变异。14项穗部性状中,第一外稃芒长和第一颖长都可分为6个显著差异性的组合,说明这两个性状种间存在明显的差异,可作为种的分类依据。8种披碱草属牧草均在第一颖芒长和旗叶与穗基部长度这2个性状上表现出较大变异,第一外稃长和第一内稃长上表现出较小变异,说明这些性状具有相对的稳定性,用于物种的遗传变异分析是有效的。从8种披碱草属牧草居群的遗传多样性指数来看,披碱草属牧草各居群存在丰富的遗传多样性,均远远高于麦类作物(中国普通小麦(Triticum aestivum)的遗传多样性指数[14]。垂穗披碱草和老芒麦的居群总遗传多样性指数较高,黑紫披碱草的居群总遗传多样性指数最低,说明往往分布范围广的物种多样性指数较高,这可能是由于生境广,植物种类繁多,物种间竞争激烈,迫于环境的选择压力,使得该物种基因得以积累。披碱草属牧草这种较高的遗传变异及不同的表型分化系数主要有以下几个方面的原因,首先,Hamrick和Godt[15]曾对涉及165个属、449个种的不同类型植物的遗传变异水平和居群分化程度进行统计,结果发现自花授粉植物51%的遗传变异存在于居群间,异交风媒植物只有9.9%的遗传变异存在于居群间[16],所以从这个结论来看这8种披碱草属牧草并不属于严格意义上的自花授粉植物,可能进行天然杂交,致使遗传物质重新组合;可能的种间交流也是增加遗传多样性的途径之一(8种披碱草属牧草种植于同一生境)。其次,遗传变异程度与人工选育的选择压力有密切的关系,一般来讲,天然居群其遗传多样性要高于栽培品种,本研究所用材料均取自天然居群,人为选择程度小,所以仍保留了物种较高的基因多样性。另外,从生物进化论的观点来看,任何物种都不是一成不变的,随环境条件的变化而变化。我国北方地区常年干旱少雨,草原退化、草地荒漠化、沙化日益加剧,生态极其脆弱;土壤盐碱化程度高,风蚀水蚀相当严重,多年来一直是国内外重点生态建设、治理的焦点。植物在这种恶劣生境条件下,以适应生境压力,进化积累了较多的遗传变异[17-18]。但是,生境的恶劣程度和人工选育的力度超出物种适应的最大限度,那么必将导致该物种遗传多样性的丧失甚至灭绝。因此,对植物资源利用的同时更要注重其保护。
[1]严学兵,王堃,王成章,等.不同披碱草属植物的形态分化和分类功能的构建[J].草地学报,2009,17(3):275.
[2]严学兵,郭玉霞,周禾,等.披碱草属植物分类和遗传多样性的研究现状[J].草业科学,2005,22(12):1-7.
[3]马啸,周永红,于海清,等.野生垂穗披碱草种质的醇溶蛋白遗传多样性分析[J].遗传,2006,28(6):699-706.
[4]李永祥,李斯深,李立会,等.披碱草属 12个物种遗传多样性的ISSR和SSR比较分析[J].中国农业科学,2005,38(8):1522-1527.
[5]史冀伟,高爱农,刘津刚,等.小麦族中间鹅观草不同居群的形态多样性分析[J].植物遗传资源学报,2009,10(4):547-552.
[6]高飞,柴守诚,高爱农.基于形态学的中华鹅观草遗传多样性分析[J].麦类作物学报,2006,26(3):12-17.
[7]王赞,高洪文,韩建国,等.柠条锦鸡儿不同居群形态变异研究[J].西北植物学报,2005,25(1):0118-0123.
[8]袁庆华,张吉宇,张文淑,等.披碱草和老芒麦野生居群生物多样性研究[J].草业学报,2003,12(5):44-49.
[9]肖海峻,徐柱,翟利剑,等.鹅观草居群表型多样性研究[J].西北植物学报,2005,13(2):111-116.
[10]孙建萍,袁庆华.披碱草属种质资源研究进展[J].草业科学,2005,22(7):2-5.
[11]王照兰,赵来喜.老芒麦种质资源描述规范和数据标准[M].北京:中国农业出版社,2007.
[12]West N E.Biodiversity of rangelands[J].Journal of Rangland Management,1993,46:2-13.
[13]苗佳敏,钟金城,陈智华.披碱草属种质资源研究现状[J].草业与畜牧,2007,27(11):2222-2227.
[14]董玉琛,曹永生,张学勇,等.中国普通小麦初选核心种质的产生[J].植物遗传资源学报,2003,4(1):1-8.
[15]Hamrick J L,Godt M J W.Allozyme diversity in plant species[A].In:Brown A H D,Clegg M T,Kahler A L.Plant Population Genetics,Breeding and Genetic Resources[C].Sunderland:Sinauer,1990:43-63.
[16]鄢家俊,白史且,马啸,等.川西北高原野生老芒麦居群穗部形态多样性研究[J].草业学报,2007,16(6):99-106.
[17]李俊清.物种遗传多样性保护及其分子生物学研究方法[J].生态学杂志,1994,13(6):27-33.
[18]李景欣,云锦凤.科尔沁沙地冰草居群形态多样性研究[J].内蒙古民族大学学报(自然科学版),2005,20(1):67-70.
猜你喜欢
草业学报(2022年11期)2022-11-23
美文(2022年18期)2022-10-20
草地学报(2022年8期)2022-08-31
现代畜牧科技(2021年7期)2021-07-28
草业科学(2021年5期)2021-06-18
西藏农业科技(2021年4期)2021-04-18
种子(2019年8期)2019-09-11
生态学报(2019年4期)2019-04-08
中国畜禽种业(2019年7期)2019-01-06
草业学报(2018年7期)2018-07-30
