
杭州湾及钱塘江河口水鸟群落组成、季节动态及种间相关性分析
2011-12-25 06:39蒋科毅邵学新
Zoological Research 2011年6期
蒋科毅, 吴 明, 邵学新
(中国林科院亚热带林业研究所 国家林业局杭州湾湿地生态系统定位研究站, 浙江 富阳 311400)
杭州湾及钱塘江河口水鸟群落组成、季节动态及种间相关性分析
蒋科毅, 吴 明, 邵学新
(中国林科院亚热带林业研究所 国家林业局杭州湾湿地生态系统定位研究站,浙江 富阳311400)
2007年7月—2010年11月, 对杭州湾曹娥江口至甬江口间潮间带及其附近海涂水库的水鸟群落进行定期监测, 共记录9目18科128种。其中, 候鸟119种, 占总数的93%; 而国家重点保护鸟类11种。对13种鸻鹬类和9种鸭类进行种群数量随时间变化的种间相关性分析, 两个类群各有21对和23对显著相关。通过对水鸟季节动态及其迁徙模式分析, 将其迁徙过程分为6个阶段, 即:4月上旬—5月中旬、7月下旬—9月下旬(旅鸟迁徙高峰期)、2月下旬—3月下旬、10月上旬—12月中旬(冬候鸟迁徙高峰期)、12月下旬—次年2月中旬(越冬期)和5月下旬—7月中旬(繁殖期)。种间吸引作用可能是影响该区域迁徙季节鸻鹬类和越冬地鸭类群落组成的重要原因, 而该区域特殊地理位置和水鸟群落构成可能是造成鸻鹬类春秋季迁徙高峰数量巨大差异的主要因素。
迁徙水鸟; 种间相关性; 迁徙模式
杭州湾为我国东部沿海钱塘江口延伸的河口湾, 其正处于东亚—澳大利亚候鸟迁徙路线上的中间位置(Wilson & Barter, 1998), 是重要的迁徙鸟类中转停歇地和越冬地(Barter, 2002)。过去在杭州湾及钱塘江河口开展的水鸟研究多仅涉及北岸(Lu et al, 1988; Qian et al, 1985; Wang & Qian, 1988; Ge et al, 2006;而涉及杭州湾南岸的相关研究较少(Zhuge et al, 1986; Zhu & Yang, 1988; Zhu, 1989a; Zhu et al, 1989b)。
鸟类的迁徙活动受各种因素制约(Dänhardt & Lindström, 2001), 种间吸引作用对鸟类迁徙活动的影响已获得较多学者的关注(Thomson et al, 2003; Forsman & Thomson, 2008; Hromada et al, 2008; Forsman et al, 2009)。目前, 国内对水鸟迁徙动态的研究已有较多报道(Ma et al, 2002a, b; Zhang et al, 2004; Ge et al, 2006; Yang et al, 2009), 但将种间相互作用运用于水鸟迁徙动态分析的研究仍较少(Zhang et al, 2004; Yang et al, 2009)。
杭州湾地处长江三角洲经济发达的沪、杭、甬金三角地带, 其保护与利用的矛盾非常突出。在该区域开展水鸟资源调查, 以了解其群落组成和季节动态, 并研究迁徙水鸟的种间相关性, 将为该区域水鸟资源监测、保护和合理利用湿地资源提供重要的基础资料和理论依据。
1 研究区域概况
杭州湾位于我国东部沿海的浙江省东北部,为喇叭状河口湾, 西界与钱塘江相连, 东界为芦漕港闸与长跳咀边线。湿地总面积5.86×104hm2,其中浅海水域3.18×104hm2、潮间淤泥海滩2.05×104hm2、潮间盐水沼泽6 244 hm2, 海拔−5~5 m(Zhejiang Forestry Administration, 2002)。除钱塘江河口外, 湾内还包含曹娥江、甬江等重要河口,其南岸属淤涨型海岸, 三北浅滩是浙江最大的海涂,而北岸则属侵蚀型海岸。湿地类型以浅海水域和期间淤泥海滩为丰, 其它尚有潮间盐水沼泽和岩石性海岸。杭州湾为一喇叭口形状的河口海湾, 湾内海域水深都小于l0 m, 水下地形平坦, 中北部至口门为杭州湾水下浅滩; 湾内有大小岛屿69个, 岛屿附近发育有潮流深槽、冲刷深潭及潮流沙脊; 海岸线长258.49 km, 以人工海岸为主。
近年来持续的大规模围垦使得杭州湾滩涂湿地急剧减少, 其中2004—2008年间每年减少10%以上(National Marine Environmental Monitoring Center, 2009), 由于普遍采用了中、低滩围垦方式,现有自然滩涂上的植被状况较差, 以低滩盐藻光滩为主, 狭窄带状分布的植被覆盖区域较少。为了解决围垦区生活用水和工农业用水问题, 杭州湾南岸目前有中小型海涂水库10余个, 水深均在3 m以上,面积最大的四灶浦水库, 达5.12 km2, 水深5.7 m。
2 研究方法
2007年7月—2008年6月, 对杭州湾上虞曹娥江口至镇海甬江口之间潮间带及其附近海涂水库的水鸟群落进行逐月统计。在随后的2008年7月—2010年11月继续开展季度监测。我们将一年分为4期:12—次年2月(越冬期)、3—5月(北迁期)、6—8月(繁殖期)和9—11月(南迁期)。季度监测是指每年每期选择其中一个月进行调查, 为了更好地掌握研究区域内的鸟类资源组成和动态, 第二年的同期调查时间顺延至下一个月进行, 循环往复。
采用直接计数法, 直接记录水鸟种类和数量,计数方法以精确记数为主, 估算为辅, 并结合“集团统计法”, 推算水鸟的总数以及群落中各物种所占百分比(Howes & Endagama, 1995)。调查时用Kowa BD42-8GR双筒望远镜和Zeiss Diascope 85T单筒望远镜观察鸟类。鸟类分类系统参考《中国鸟类分类与分布名录》(Zheng, 2005), 中文鸟名主要参考《中国鸟类野外手册》(Mackinnon et al, 2000)。
调查时间, 潮间带鸟类调查根据潮汛情况, 选择在高潮点前后2 h内进行。为避免调查数据重复,在鸟类数量统计中采用最大值保留法(Howes & Bakewell, 1989), 并将种群数量超过10%的种群定为优势种。
对水鸟物种数的月动态分析采用了2007年7月—2010年11月的所有监测数据。利用2007年7月—2008年6月的逐月调查数据, 对水鸟数量的年动态进行分析, 并对记录的鸻鹬类和鸭类分别进行两两配对的种间数量随时间变化的Spearman相关性分析。为了提高分析的准确性, 在各迁徙阶段中种群数量低于1%的物种未纳入上述分析。数据处理采用SPSS10.0进行。
3 结 果
3.1 水鸟种类组成
杭州湾及钱塘江河口湿地共记录水鸟9目18科128种(表1)。候鸟119种, 占总数的93.0%〔其中, 冬候鸟64种(50.0%), 旅鸟34种(26.6%), 夏候鸟17种(13.3%)及迷鸟4种(3.1%)〕。繁殖鸟(夏候鸟和留鸟)26种, 占该地水鸟总数的20.3%。鸻形目、雁形目、鹳形目和鸥形目为记录物种最多的4个目,合计占该地水鸟总数的87.5%。本区域的优势种随迁徙阶段的变化而变动, 自然潮间带的优势种有黑腹滨鹬(Calidris alpina)、黑尾塍鹬(Limosa limosa)、红颈滨鹬(Calidris ruficollis)等12种, 海涂水库的优势种包括罗纹鸭(Anas falcata)、斑嘴鸭(Anas poecilorhyncha)等5种。
杭州湾及钱塘江河口湿地共记录列入国家重点保护野生动物名录的水鸟有11种, 包括国家I级重点保护鸟类东方白鹳(Ciconia boyciana)、中华秋沙鸭(Mergus squamatus)和遗鸥(Larus relictus), 列入IUCN中国鸟类受威胁状况名录的鸟类数量17种, 包括濒危EN等级的中华秋沙鸭、青头潜鸭(Aythya baeri)、东方白鹳和黑脸琵鹭(Platalea minor)。此外, 杭州湾及钱塘江河口共有88种水鸟列入中日候鸟保护名录, 42种列为中澳候鸟保护名录。
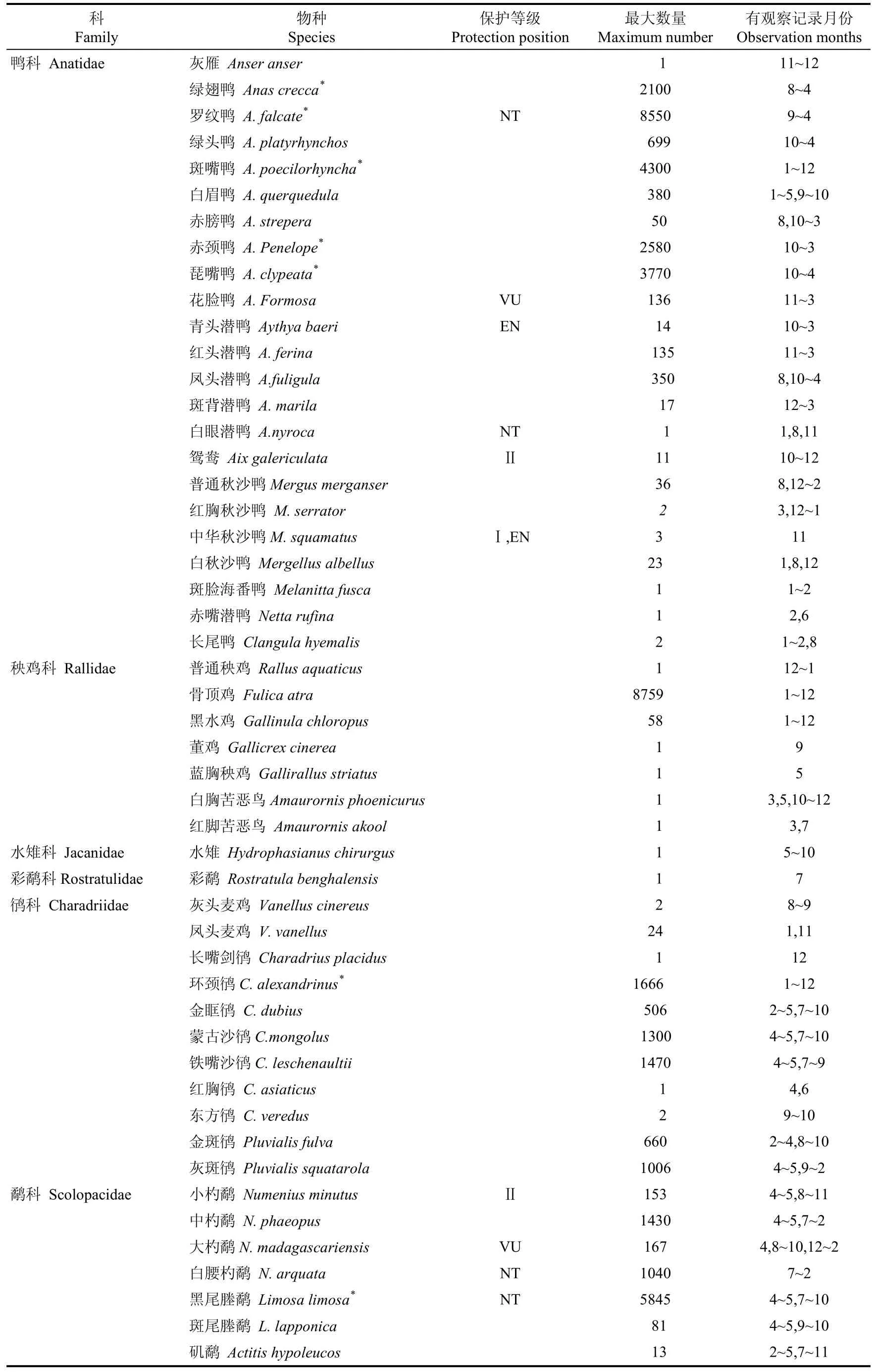
科Family物种Species保护等级Protection position最大数量Maximum number有观察记录月份Observation months鸭科 Anatidae 灰雁 Anser anser 1 11~12绿翅鸭 Anas crecca* 2100 8~4罗纹鸭 A. falcate* NT 8550 9~4绿头鸭 A. platyrhynchos 699 10~4斑嘴鸭 A. poecilorhyncha* 4300 1~12白眉鸭 A. querquedula 380 1~5,9~10赤膀鸭 A. strepera 50 8,10~3赤颈鸭 A. Penelope* 2580 10~3琵嘴鸭 A. clypeata* 3770 10~4花脸鸭 A. Formosa VU 136 11~3青头潜鸭 Aythya baeri EN 14 10~3红头潜鸭 A. ferina 135 11~3凤头潜鸭 A.fuligula 350 8,10~4斑背潜鸭 A. marila 17 12~3白眼潜鸭 A.nyroca NT 1 1,8,11鸳鸯 Aix galericulata Ⅱ 11 10~12普通秋沙鸭Mergus merganser 36 8,12~2红胸秋沙鸭 M. serrator 2 3,12~1中华秋沙鸭M. squamatus Ⅰ,EN 3 11白秋沙鸭 Mergellus albellus 23 1,8,12斑脸海番鸭 Melanitta fusca 1 1~2赤嘴潜鸭 Netta rufina 1 2,6长尾鸭 Clangula hyemalis 2 1~2,8秧鸡科 Rallidae 普通秧鸡 Rallus aquaticus 1 12~1骨顶鸡 Fulica atra 8759 1~12黑水鸡 Gallinula chloropus 58 1~12董鸡 Gallicrex cinerea 1 9蓝胸秧鸡 Gallirallus striatus 1 5白胸苦恶鸟Amaurornis phoenicurus 1 3,5,10~12红脚苦恶鸟 Amaurornis akool 1 3,7水雉科 Jacanidae 水雉 Hydrophasianus chirurgus 1 5~10彩鹬科Rostratulidae 彩鹬 Rostratula benghalensis 1 7鸻科 Charadriidae 灰头麦鸡 Vanellus cinereus 2 8~9凤头麦鸡 V. vanellus 24 1,11长嘴剑鸻 Charadrius placidus 1 12环颈鸻C. alexandrinus* 1666 1~12金眶鸻 C. dubius 506 2~5,7~10蒙古沙鸻C.mongolus 1300 4~5,7~10铁嘴沙鸻C. leschenaultii 1470 4~5,7~9红胸鸻 C. asiaticus 1 4,6东方鸻 C. veredus 2 9~10金斑鸻 Pluvialis fulva 660 2~4,8~10灰斑鸻 Pluvialis squatarola 1006 4~5,9~2鹬科 Scolopacidae 小杓鹬 Numenius minutus Ⅱ 153 4~5,8~11中杓鹬 N. phaeopus 1430 4~5,7~2大杓鹬N. madagascariensis VU 167 4,8~10,12~2白腰杓鹬 N. arquata NT 1040 7~2黑尾塍鹬 Limosa limosa* NT 5845 4~5,7~10斑尾塍鹬 L. lapponica 81 4~5,9~10矶鹬 Actitis hypoleucos 13 2~5,7~11
* 代表该物种是某个迁徙阶段的优势种;* Represents a dominant species in the waterbird community of one or more migration stages;I和Ⅱ:分别代表国家一和二级重点保护野生动物;I represents the national first-grade protection of wildlife in China and II represents the second-grade level of protection;CR、EN、VU、NT:世界自然保护联盟IUCN保护等级IUCN Red List Categories and Criteria, CR:极危(Critically Endangered);EN:濒危(Endangered);VU:易危(Vulnerable);NT:近危(Near Threatened)。
3.2 水鸟种类及数量动态
杭州湾及钱塘江河口水鸟物种数的年动态曲线总体上呈现两个峰值(图1), 分别是10月73种和4月70种, 而6月份水鸟物种数全年最低, 仅记录24种, 另一个低谷出现在3月(54种)。对鸻形目、雁形目、鹳形目和鸥形目这4种优势类群物种数动态分析结果显示, 鸻形目和雁形目物种数年动态波动较大, 而鹳形目和鸥形目物种数波动较小; 鸻形目鸟类物种数年动态呈现“双峰”曲线, 分别在4和9月份达到最高值36和39种, 其低谷值出现在3和6月, 分别为11和4种。雁形目鸟类物种数动态总体上表现为一单峰曲线, 峰值出现在12月, 记录25种, 5—7月为低谷期, 仅记录1~3种。鹳形目和鸥形目物种数的最小值分别出现1—3月和6—7月, 分别仅记录7和2种。
杭州湾及钱塘江河口水鸟个体数量的年动态曲线与其物种数动态曲线存在较大差异, 总体上呈现3个峰值(图2), 其中最低值出现在6月, 记录个体数量为4 316只, 最高值出现在3月初, 记录水鸟114 395只。鸻形目鸟类数量动态曲线与总曲线较为相似, 其最小值出现在6月(430只), 最高值在3月(101297只); 雁形目鸟类数量动态曲线呈“双峰”,峰值分别出现在11和3月, 分别记录10 622和
10 862只; 与前两者相比, 鹳形目和鸥形目个体数量总体偏少, 波动相对平缓; 鹳形目个体数量高峰期出现在繁殖期6和8月, 分别记录了3 561和11 367只, 其余月份记录数量均低于600只; 鸥形目数量动态曲线总体上呈现3个峰值, 分别为9、 11和3月, 分别记录1 628、4 892和1 348只。
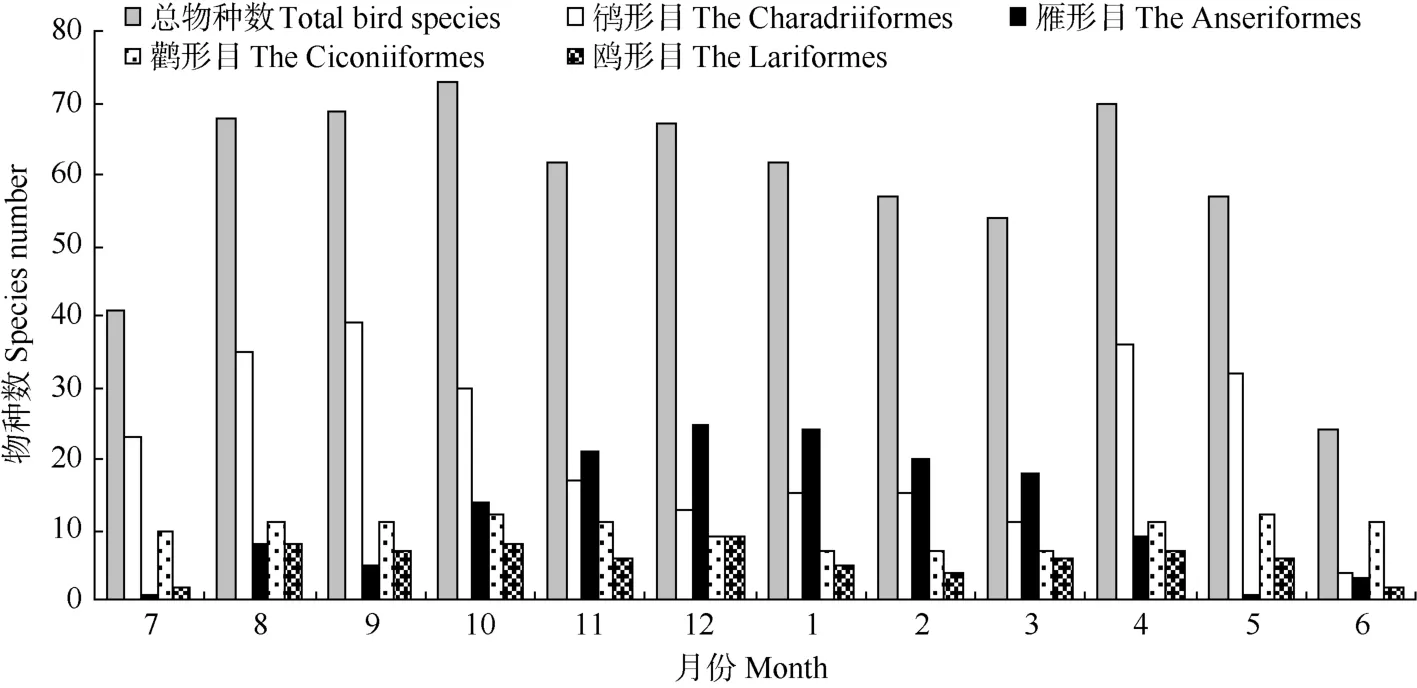
图1 杭州湾及钱塘江河口水鸟物种数的月动态Fig. 1 Monthly dynamics of waterbirds species in Qiantangjiang River estuary and Hangzhou Bay
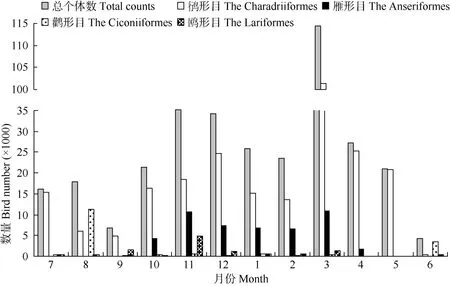
图2 杭州湾及钱塘江河口水鸟数量的月动态Fig. 2 Monthly dynamics of waterbirds number in Qiantangjiang River estuary and Hangzhou Bay
3.3 迁徙水鸟种间相关性分析
对杭州湾及钱塘江河口鸻鹬类和鸭类的物种数量变化进行了种间相关性分析结果显示(表2、表3), 进行两两配对相关性分析的13种鸻鹬类中, 有20对为显著正相关, 其中13对为极显著正相关, 有1对为显著负相关。蒙古沙鸻(Charadrius mongolus)、铁嘴沙鸻(C. leuschenaultii)、黑尾塍鹬、红颈滨鹬和尖尾滨鹬(Calidris acuminate)之间均极显著正相关。金眶鸻(C. dubius)与前4种鸻鹬类(不包括红颈滨鹬)显著正相关, 且与金斑鸻(Pluvialis dominica)为极显著正相关, 与青脚鹬为显著正相关。与金眶鸻类似, 青脚鹬与前5种极显著正相关鸻鹬旅鸟群中的4种(不包括铁嘴沙鸻)显著正相关, 其中与蒙古沙鸻、红颈滨鹬极显著相关, 还与金眶鸻显著相关。灰斑鸻与金斑鸻显著负相关。
9种鸭类进行了种间相关性分析, 有23对为显著正相关, 其中10对为显著正相关。除针尾鸭(Anasacuta)和琵嘴鸭(A. clypeata)之间无显著相关性外,绿翅鸭(A. crecca)、罗纹鸭、斑嘴鸭、赤颈鸭(A. penelope)、凤头潜鸭(Aythya fuligula)、针尾鸭与琵嘴鸭之间均两两显著正相关。绿头鸭(Anas platyrhynchos)仅与凤头潜鸭、针尾鸭和绿翅鸭之间显著正相关。而白眉鸭(A. querquedula)与其它任一种鸭类均无显著相关性。
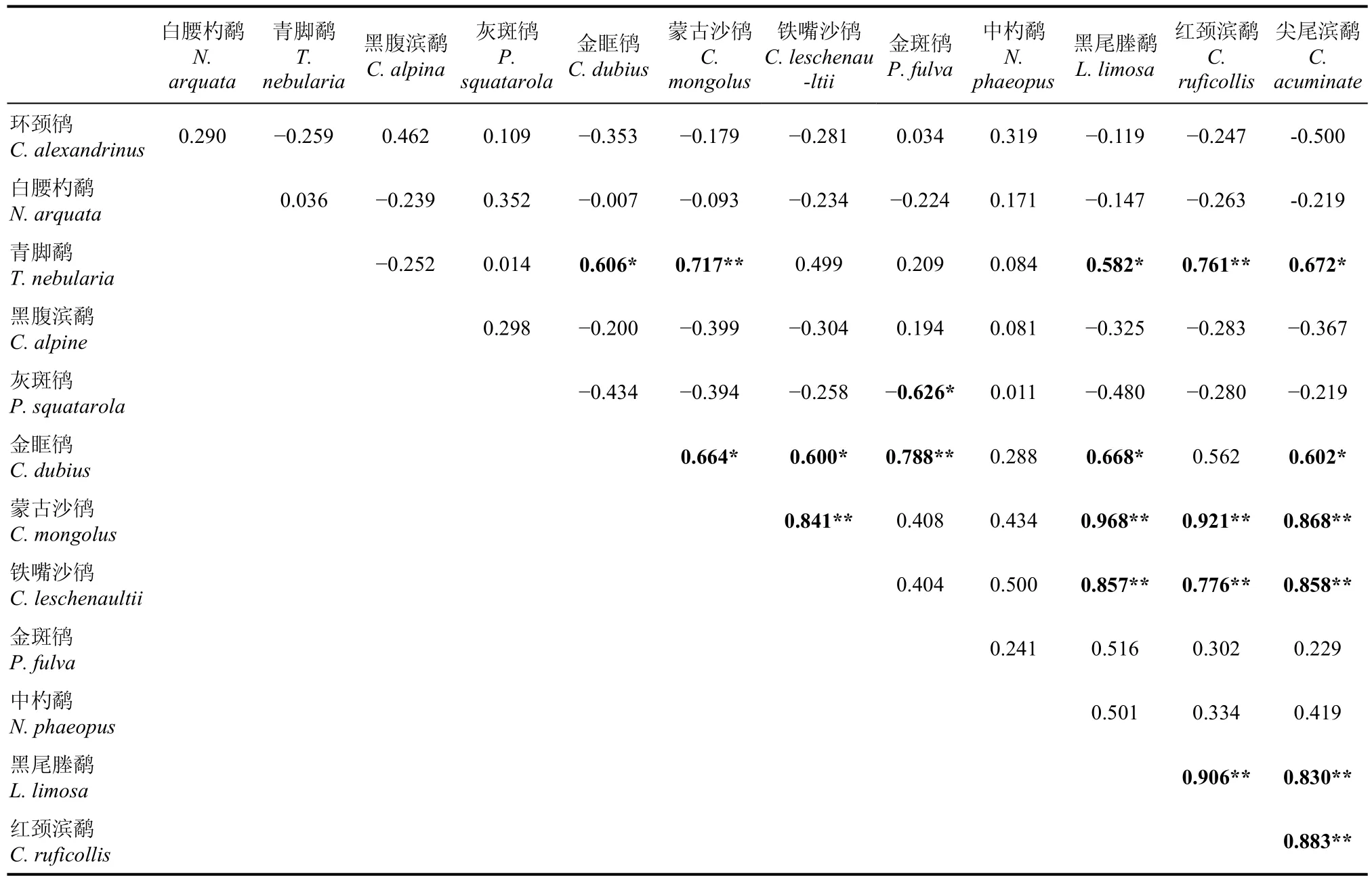
表2 13种鸻鹬类数量变化的相关系数及显著性矩阵Tab. 2 Correlation coefficient matrix of 13 species of shorebirds in Qiantangjiang River estuary and Hangzhou Bay
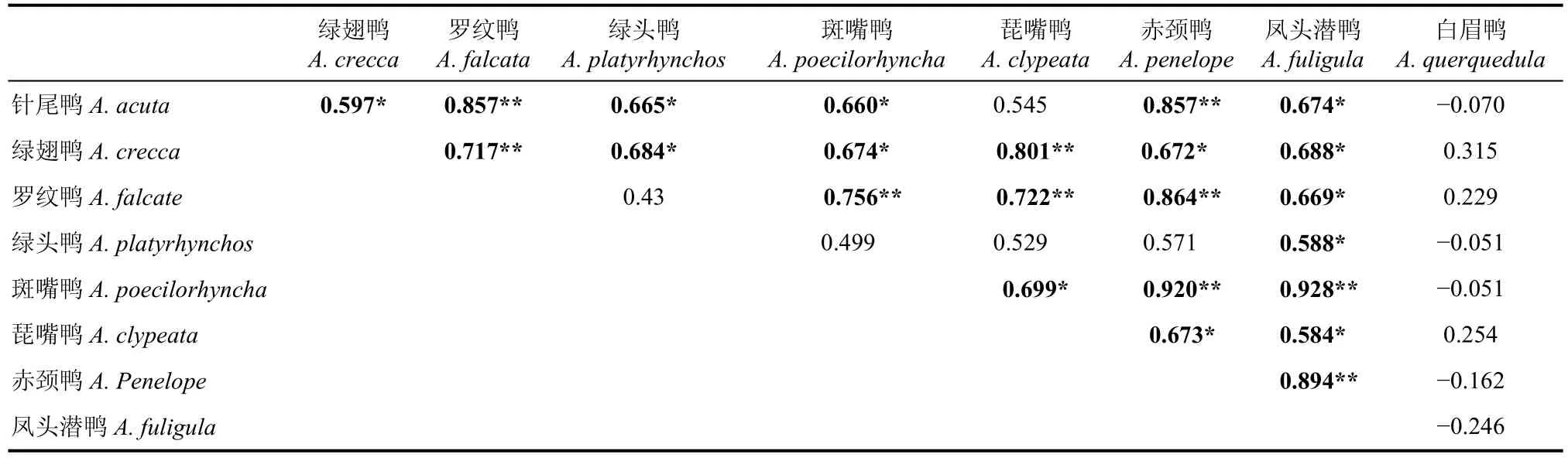
表3 8种鸭类数量变化的相关系数及显著性矩阵Tab. 3 Correlation coefficient matrix of eight species of duck in Qiantangjiang River estuary and Hangzhou Bay
4 讨 论
4.1 种间相关性与水鸟迁徙模式
研究结果显示, 在杭州湾及钱塘江河口出现的鸻鹬类中, 参与种间相关性分析的7种旅鸟中的5种(包括黑尾塍鹬、红颈滨鹬、尖尾滨鹬、蒙古沙鸻和铁嘴沙鸻)之间呈现两两显著正相关, 表明它们的迁徙模式是相同的, 而中杓鹬和金斑鸻与前5者之间及两者内部均没建立这种显著性联系。这可能表明旅鸟群中至少存在3种迁徙模式。以上述5种显著相关性鸻鹬类旅鸟为优势种构成的迁徙水鸟群, 其迁徙时间集中在7月下旬—9月下旬(秋季南迁时期)和3月下旬—5月中旬(春季北迁时期)。参与相关性分析的鸻鹬类冬候鸟种间均无显著相关性,表明黑腹滨鹬、白腰杓鹬、灰斑鸻、青脚鹬等冬候鸟之间的迁徙模式可能是不同的。金眶鸻、青脚鹬均与由5种旅鸟组成的极显著相关旅鸟群中的4种显著正相关, 表明其具有与后者类似的迁徙模式。生态位相近的灰斑鸻与金斑鸻显著负相关, 两者的迁徙模式可能截然相反。中杓鹬、白腰杓鹬、黑腹滨鹬和环颈鸻与参与分析的其他13种鸻鹬类之间均无显著相关性, 表明其具有相对独立的迁徙节律。
而在该区域越冬的鸭类群落中, 5种优势种包括绿翅鸭、罗纹鸭、斑嘴鸭、琵嘴鸭和赤颈鸭及凤头潜鸭之间显示了显著的正相关性, 针尾鸭与前5种鸭类中的4种(不包括琵嘴鸭)建立了种间显著正相关, 仅白眉鸭与其他鸭类无显著相关性(表3)。这表明鸭类的迁徙模式相对单一, 大多数鸭类都从10月上旬开始大规模的迁徙至杭州湾, 并于次年4月中旬全部迁离。
杭州湾是东亚—澳大利西亚水鸟迁徙通道中一个重要的中转停歇地(Barter, 2002), 而种间吸引作用可能是杭州湾迁徙水鸟群落形成的一个重要驱动因子。鸟类群落中的种间吸引作用已被实验所证实(Mikko & Jukka, 2002; Robert et al, 2003; Sebastián-González et al, 2010), 但Mikko et al(1999)认为, 种间吸引作用可能仅在种间竞争足够低时才处于优势。食物资源是影响鸟类中途停歇地选择的重要因素之一(Hicklin & Smith, 1979; Wilson, 1990)。但已有研究表明,食物资源可能并不是杭州湾鸻形目鸟类的迁徙规模和数量动态的限制因素。杭州湾潮间带大型底栖动物功能群的密度和绝大多数种类数分布在季节间差异均不显著(Li et al, 2007), 崇明东滩的研究结果与此类似(Jing, 2005)。我们的研究结果显示(图2), 杭州湾鸻鹬类水鸟群落的数量的季节变动最高值在10万只左右, 表明除该高峰期外, 杭州湾的食物资源相对于鸻鹬类来说可能是充裕的, 因而鸻鹬类因食物而出现的种内和种间的竞争将处于较低的水平。进入越冬期(12—次年2月)的鸭类群落数量均大幅低于春秋季迁徙季节(图2), 表明此时的鸭类群落数量处于相对稳定的状态, 其种间竞争水平可能较低。
出于共同的生境需求, 个体可能将不同种的存在状况作为评价栖息地斑块质量的一个指标(Mikko & Jukka, 2002; Jukka & Robert, 2008; Hromada et al, 2008)。这对于刚到达繁殖地的迁徙鸟类来说更为重要(Robert et al, 2003; Jukka & Robert, 2008; Hromada et al, 2008; Jukka et al, 2009)。对于是否停留它们必须快速地做出抉择, 而留鸟的出现与否及种群情况将是其重要的参考因素(Sebastián-González et al, 2010)。种间吸引假说同时认为, 多物种的群落中的个体能够降低其被捕食风险(Mikko et al, 1990)。本研究结果中, 7种参与分析的旅鸟中的6种与反映该区域栖息地质量的繁殖鸟金眶鸻或冬候鸟青脚鹬显著正相关。而5种旅鸟组成的极显著相关群中的任两种的最初接触可能也是基于一方能反映另一方迁徙路线上的中转停歇地的生境质量, 最终基于共同的生境需求(获取食物和降低捕食风险), 同时最小化种间排斥作用而有选择的集群迁徙。Recher(1966)认为迁徙鸻鹬类集群中所有的种间差异和瞬态的集群特征都明显为了最大限度的减少种间排斥作用。这种观点也体现在我们的研究结果中, 极显著相关群中的5种旅鸟的喙长是不同的, 而喙长与取食方式有关(Brown & Wilson, 1956), 不同的喙长意味着食物选择的分化(Wang & Qian, 1988)。
对于在杭州湾越冬的鸭类来说, 由于地处长江三角洲经济发达的沪、杭、甬金三角地带, 降低被捕食(主要为人类的捕食活动)风险将是影响其栖息地选择尤其是休息场所选择的重要因素之一。本研究结果中, 凤头潜鸭与除白眉鸭外的7种鸭类显著正相关, 而其也是最先来到杭州湾的鸭类之一, 但其食性以动物性食物为主, 因而其在水库的出现与否及种群数量大小可能成为后来者评价该水库生境质量(不包括食物资源)的重要指标。
4.2 杭州湾及钱塘江河口水鸟群落迁徙规律
根据杭州湾及钱塘江河口水鸟物种及数量迁徙动态(图1、图2)的研究结果, 本区域水鸟群落整个迁徙过程可分为6个阶段。
第一阶段从7月下旬开始—9月下旬, 迁徙高峰出现在8月中旬, 这一阶段的迁徙水鸟群落的主体是以黑尾塍鹬、红颈滨鹬及蒙古沙鸻为优势种群的鸻鹬类旅鸟。在该阶段, 鹳形目的鹭鸟正处于繁殖期, 8月中旬大量亚成鸟离巢, 鹭鸟数量达到最高峰, 9月下旬鹭科夏候鸟陆续南迁; 而鹳形目白琵鹭和黑脸琵鹭从7月下旬开始—8月陆续迁飞至此短暂停歇。以白额燕鸥为主体的鸥类繁殖鸟及鸥嘴噪鸥(Gelochelidon nilotica)开始南迁, 同时以西伯利亚银鸥(Larus vegae)和黑嘴鸥为主体的冬候鸟开始迁飞到达。
第二阶段从10月上旬—12月中旬, 迁徙水鸟群落以冬候鸟的鸻鹬类和雁鸭类为主。这一阶段的鸻鹬类以黑腹滨鹬为主要优势种。从10月上旬开始, 大规模鸭群陆续到来, 11月下旬鸭类群落数量达到一个小高峰, 并在12月中旬进入相对稳定期。从11上旬开始, 大型水鸟如鹳形目的黑脸琵鹭、白琵鹭、雁鸭类的小天鹅、豆雁(Anser fabalis)以及鹈形目的卷羽鹈鹕等陆续迁飞至此。11下旬, 以黑尾鸥(Larus crassirostris)为优势种的鸥类冬候鸟个体数量达到了全年的最高值。
第三阶段从12月下旬—次年2月中旬, 为越冬稳定期。这时水鸟群落以冬候鸟为主, 群落物种组成与数量相对比较稳定, 鸻鹬类以黑腹滨鹬为绝对优势种, 其种群数量占总数的80%以上。
第四阶段从2月下旬—3月下旬, 为水鸟迁徙期。该阶段以黑腹滨鹬为主的冬候鸟迁徙首先开始, 2月下旬—3月上旬, 黑腹滨鹬的种群数量骤升至前期的5倍以上, 之后其数量回落到越冬期相近的水平。3月中旬, 鸭类迁徙高峰到来, 其个体数量在3月下旬达到一个高峰之后迅速下降。3月下旬鸻鹬类旅鸟陆续迁来。小天鹅则在2月下旬就已经迁离。3月中旬鹈形目的卷羽鹈鹕迁飞至此, 短暂停留后即迁离。
第五阶段从4月上旬—5月中旬, 为以红颈滨鹬、尖尾滨鹬及黑尾塍鹬为优势种的旅鸟群的迁徙高峰期。但该阶段, 以黑腹滨鹬为优势种的冬候鸟群仍为水鸟群落的主体。鸭类在4月中旬基本迁离。3月下旬—4月下旬, 陆续有鹳形目黑脸琵鹭和白琵鹭迁飞至此, 作短暂停留后即迁离。
第六阶段从5月下旬—7月中旬, 为候鸟的繁殖季节。此阶段的水鸟群落主要由鹭鸟和以白额燕鸥为优势种的鸥类及普通燕鸻等夏候鸟构成。鹭鸟夏候鸟从2月开始就陆续迁来, 5月至7月进入繁殖高峰期。4月中下旬, 普通燕鸻就已经迁来, 而白额燕鸥也在此时出现在杭州湾, 但前者的个体数量在7月中旬即达到高峰, 而后者则在繁殖季节结束后的8月下旬—9月上旬达到高峰。
从杭州湾及钱塘江河口水鸟季节动态分析中,我们发现该区域春季鸻鹬类迁徙高峰在数量上远超过秋季迁徙高峰, 达到了后者的4倍以上, 类似的研究结果出现在崇明东滩(Qian et al, 1985; Ma et al, 2002b)。马志军等认为, 春季鸻鹬类迁徙高峰数量远高于秋季的主要原因在于崇明东滩在鸻鹬类迁徙路线中的特殊地理位置(Ma et al, 2002b)。我们的研究结果与此类似, 杭州湾及钱塘江河口水鸟秋季迁徙高峰数量上的巨大差异来自于该区域的特殊地理位置和水鸟群落构成。对春秋季迁徙季节的水鸟群落组成分析结果显示, 从10月上旬开始—次年3月上旬, 黑腹滨鹬的种群数量占鸻鹬类群落总数的比例均维持在80%以上, 并在3月上旬达到98%, 其后的4—5月, 虽然其种群数量开始下降,但仍维持在总数的60%以上。已有研究表明太平洋东海岸的黑腹滨鹬会采取一系列的短距离轻跳方式, 沿海岸北上到达繁殖地, 而南迁过程中, 则可能直接飞越阿拉斯加海湾到达较南的停歇地或越冬地(Warnock et al, 2004)。位于太平洋西海岸的东亚—澳大利西亚水鸟迁徙通道的黑腹滨鹬也可能采用类似的迁徙方式。崇明东滩的黑腹滨鹬在北迁时具有较高的能量储备, 可能是为迁徙飞行做准备(Cai, 2009)。由于黑腹滨鹬在中国东南部地区越冬(Barter et al, 2006), 杭州湾所在的区域有可能是其迁飞前的最后一个能量补给站, 因而该区域黑腹滨鹬的种群数量在其迁徙前夕急剧增加, 但在其2月下旬—3月上旬的首轮迁徙高峰结束后, 黑腹滨鹬的种群数量显著回落。
Barter M. 2002. Shorebirds of the Yellow Sea: Importance, threats and conservation status. Australia[M]//Wetlands International Global Series 9: International Wader Studies 12. Canberra, Australia: [s.n.].
Barter M, Lei G, Cao L. 2006. Waterbird Survey of the Middle and Lower Yangtze River Floodplain in February 2005[M]. Yang Q, Translation. Beijing: China Forestry Publishing House. [马克·巴特,雷刚,曹垒. 2006.长江中下游水鸟调查报告(2005年2月).杨琴,译.北京:中国林业出版社.]
Brown WL, Wilson EO. 1956. Character displacement[J]. Syst. Zool,5(2): 49-64.
Cai ZY. 2009. Ecology of Dunlin (Calidris alpine) in Chongming Dongtan[D]. MSc thesis, Fudan University. [蔡志扬. 2009.崇明东滩黑腹滨鹬的生态学研究.硕士学位论文,复旦大学.]
Dänhardt J, Lindström Å.2001. Optimal departure decision of songbirds from an experimental stopover site and the significance of weather[J]. Anim Behav.,62: 235-243.
Ge ZM, Wang TH, Zhou X, Shi WY. 2006. Seasonal change and habitat selection of shorebird community at the South Yangtze River Mouth and North Hangzhou Bay, China[J]. Acta Ecol Sin,26(1):40-47. [葛振鸣,王天厚,施文彧,周晓. 2006.长江口杭州湾鹆形目鸟类群落季节变化和生境选择.生态学报,26(1): 40-47. ]
Hicklin PW, Smith PC. 1979. The diets of five species of migrant shorebirds in the Bay of Fundy[J]. Proc NS Ins Sci,29: 483-488.
Howes M, Endagama P. 1995. Farmers, Forests and Fuel: Towards a New Biomass Energy Strategy for Sri Lanka[M]. London: Intermediate Technology Publications.
Howes J, Bakewell D. 1989. Shorebird Studies Manual[M]. Malaysia: AWB Publication, NO.55.
Hromada M, Antczak M, Valone TJ, Tryjanowski P. 2008. Settling decisions and heterospecific social information use in shrikes[J]. PLoS ONE,3(12): e3930.
IUCN. 2011. IUCN Red List of Threatened Species.Version 2011.1.[R/OL] http://www.iucnredlist.org.
Jing K. 2005. Stopover ecology of shorebirds in Chongming Dongtan, Shanghai[D]. PH.D. thesis, Fudan University, Shanghai. [敬凯. 2005.上海崇明东滩鸻鹬类中途停歇生态学研究.博士学位论文.复旦大学.]
Jukka TF, Robert LT. 2008. Evidence of information collection from heterospecifics in cavity-nesting birds[J]. Ibis,150: 409-412.
Jukka TF, Ma°rten BH, Lars G.2009. Experimental evidence for the use of density based interspecific social information in forest birds[J]. Ecography,32: 539-545.
Li HH, Bao YX, Hu ZY, Ge BM. 2007. Seasonal dynamics of macrobenthic functional groups and trophic levels in the bridge construction zone at the South Bank of Hangzhou Bay, China[J]. Acta Zool Sin,53(6): 1011-1023. [李欢欢,鲍毅新,胡知渊,葛宝明.2007.杭州湾南岸大桥建设区域潮间带大型底栖动物功能群及营养等级的季节动态.动物学报,53(6): 1011- 1023.]
Lu JJ, Shi M, Cui ZX. 1988. A preliminary study on the over-wintering wader (Charadriiformes) community in the North Coast of East China Sea[J]. Chn J Ecol,7(6): 19-22. [陆健健,施铭,崔志兴. 1988.东海北部沿海越冬鸻鹬群落的初步研究.生态学杂志,7(6): 19-22.]
Mackinnon J, Philipps K, He FQ. 2000. A Field Guide to the Birds of China [M]. Lu HF, Translation. Changsha: Hunan Education Press. [马敬能,菲利普斯,何芬奇. 2000.中国鸟类野外手册.卢和芬,译.长沙:湖南教育出版社.]
Ma ZJ, Jing K, Tang SM, Chen JK. 2002a. Shorebirds in the Eastern intertidal areas of Chongming Island during the 2001 northward migration [J]. Stilt,41: 6-10.
Ma ZJ, Tang SM, Lu F, Chen JK.2002b. Chongming Island: a less important shorebirds stopover site during southward migration? [J]. Stilt,41: 35-37.
Mikko M, Pekka H, Gerald JN, Kent M. 1990. Numerical and behavioural responses of migrant passerines to experimental manipulation of resident tits (Parus spp.): heterospecific attraction in northern breeding bird communities? [J] Oecologia,85: 218-255.
Mikko M, Roger H, Jukka TF, Juha T. 1999 Evolution of heterospecific attraction: Using other species as cues in habitat selection[J]. Evol. Ecol,13: 91-104.
Mikko M, Jukka TF. 2002. Heterospecific attraction among forest birds: a review[J]. Ornithol Sci,1: 41-51.
National Marine Environmental Monitoring Center. 2009. The Marine Environment Quality Bulletin of China 2009 [EB/OL]. http://www. mem.gov.cn/ indexshow.html [国家海洋环境监测中心. 2009. 2009年中国海洋环境质量公报. http://www.mem.gov.cn/ indexshow.html.]
Qian GZ, Cui ZX, Wang TH. 1985. Waders (charadiiformes) community of the Chang-jiang river estuary and the Hang-zhou bay[J]. Acta Zool Sin,31(1): 96-97. [钱国桢,崔志兴,王天厚. 1985.长江口杭州湾北部的鸻形目鸟类群落.动物学报,31(1): 96-97.]
Recher HF. 1966. Some aspects of the ecology of migrant shorebirds[J]. Ecology,47(3): 393-407.
Robert LT, Jukka TF, Mikko M. 2003. Positive interactions between migrant and resident birds: testing the heterospecific attraction hypothesis[J]. Oecologia,134: 431-438.
Sebastián-González E, Sánchez-Zapata JA, Botella F, Ovaskainen O. 2010. Testing the heterospecific attraction hypothesis with time-series data on species co-occurrence[J]. Proc R Soc,277: 2983-2990.
Wang TH, Qian GZ. 1988. Waders (Charadiiformes) Community of the Chang-jiang River Estuary and the Hang-zhou Bay[M]. Shanghai: East China Normal University Press. [王天厚,钱国桢. 1988.长江口杭州湾鸻形目鸟类.上海:华东师范大学出版社.]
Warnock ND, Takekawa JY, Bishop MA. 2004. Migration and stopover strategies of individual Dunlin along the Pacific coast of North America[J]. Can J Zool,82: 1687-1697.
Wilson WH Jr. 1990. Relationship between prey abundance and foraging site selection by semipalmated sandpipers on a Bay of Fundy mudflat[J]. J Field Ornithol,61: 9-19.
Wilson JR, Barter MA. 1998. Identification of potentially important staging area for “long jump” migrant waders in the East Asian-Australian Flyway during northward migration[J]. Stilt,32: 16-26.
Yang CC, Cai Y, Liang W, Zhang GG, Shi HT. 2009. Waterbirds diversity, seasonal dynamics, and interspecific correlation in the wetlands of Beili and Houshui Bays, Hainan[J]. J Biodivesr Sci,17(3): 226-232. [杨灿朝,蔡燕,梁伟,张国钢,史海涛. 2009.海南北黎湾和后水湾湿地水鸟的季节动态、物种丰富度和种间相关性.生物多样性,17(3): 226-232.]
Zhang SP, Zhang ZW, Xu JL, Sun QH, Liu DP. 2004. Seasonal changes and interspecific correlation among the migratory waterbird communities in Tianjin[J]. Acta Ecol Sin,42(4): 666-673. [张淑萍,张正旺,徐基良,孙全辉,刘东平. 2004.天津地区迁徙水鸟群落的季节动态及种间相关性分析.生态学报,42(4): 666-673.]
Zhejiang Forestry Administration. 2002. Zhejiang Forestry Natural Resources-Wetland [M]. Beijing: Chinese Agriculture Science and Technology Publishing House. [浙江省林业局. 2002.浙江林业自然资源(湿地卷).北京:中国农业科学技术出版社.]
Zheng GM. 2005. A Checklist on the Classification and Distribution of the Birds of China[M]. Beijing: Science Press. [郑光美. 2005.中国鸟类分类与分布名录.北京:科学出版社.]
Zhuge Y, Jiang SR, Zheng ZW, Fang GW. 1986. Preliminary studies on geographical ecology of birds and mammals on some islands of Zhejiang Province[J]. Acta Zool Sin,32(1): 74-85. [诸葛阳,姜仕仁,郑忠伟,方国伟. 1986.浙江海岛鸟兽地理生态学的初步研究.动物学报,32(1): 74-85.]
Zhu X, Yang CJ. 1988. A survey of birds in Zhejiang Province[J].J Zhejiang For Coll,5(3): 243-258. [朱曦,杨春江. 1988.浙江鸟类研究.浙江林学院学报,5(3): 243-258.]
Zhu X. 1989a. Preliminary studies on the ecological types and geographical distribution of birds in Zhejiang[J]. J Zhejiang For Coll,6(3): 283-289. [朱曦.浙江省鸟类的生态地理初步研究. 1989.浙江林学院学报,6(3): 283-289. ]
Zhu X, Xuan ZC, Chen LQ, Gao YJ, Ma XX. 1989b. Preliminary investigation on spring bird in East Coast of Zhejiang Province[J].J Zhejiang For Sci Technol,9(1): 35-40. [朱曦,宣志灿,陈李群,高银江,麻秀新. 1989.浙江东部沿海春季鸟类初步调查.浙江林业科技,9(1): 35-40.]
Community composition, seasonal dynamics and interspecific correlation of waterbirds in the Qiantangjiang River estuary and Hangzhou Bay
JIANG Ke-Yi, WU Ming, SHAO Xue-Xin
(Research Institute of Subtropical Forestry, CAF; Wetland Ecosystem Research Station of Hangzhou Bay, State Forestry Administration, Fuyang 311400 , China)
Waterbird surveys were conducted regularly in the Qiantangjiang River estuary and Hangzhou Bay from July 2007 to November 2011. A total of 128 species (nine orders and 18 families) were recorded, including 119 migrants which accounted for 93% of the total species; eleven species were listed as National Protected Species. Inter-specific correlation analysis for 13 shorebird populations and nine duck populations recorded over time found that 21 pairs of shorebirds and 23 pairs of ducks were correlated. By looking at seasonal dynamics and migration patterns we were able to divide the migration process into six stages: (1) late July to late September was the migration peak of shorebirds, which were dominated byLimosa limosa,Calidris ruficollisandCharadrius mongolus. (2) Early October to mid-December was the migration peak of wintering migrants of shorebirds and ducks, which were the first two large groups in our study areas. (3) Late December to mid-February was the wintering period of migration waterbirds. (4) Late February to late March was the peak migration of ducks and the winter migrants of shorebirds dominated byCalidris alpina.(5) Early April to mid-May was the migration peak of passage migrants such as,Calidris ruficollis, Calidris acuminate and Limosa limosabut the population size of shorebird winter migrants dominated byCalidris alpinewas still larger than the former. (6) Late May to mid-July was the breeding season of all egrets, summer migrants of gulls and several species of shorebirds. Our surveys show that interaction among species is possibly an important determinant of community composition of shorebirds and wintering ducks during the migration season. It may be the geographical position and community composition of migrant shorebirds across Hangzhou Bay that mean during the northward migration there are far more shorebirds than during the southward migration.
book=32,ebook=110
Migratory waterbirds; Interspecific correlation; Migration pattern
Q959.7;Q958.13
A
0254-5853-(2011)06-0631-10
10.3724/SP.J.1141.2011.06631
2011-06-02;接受日期:2011-10-09
浙江省基金项目“杭州湾湿地景观格局变化对水鸟多样性的影响”(Y5080130);国家“十一五”科技攻关专题项目“典型湿地可持续综合利用技术试验示范”(2006BAD03A1904);中国林科院亚热带林业研究所专项资金项目“杭州湾湿地生物多样性监测及数据库构建”(RISF6146)
E-mail: jiangky2005@126.com
book=32,ebook=118
猜你喜欢
海洋通报(2022年4期)2022-10-10
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
幼儿教育·父母孩子版(2022年3期)2022-03-14
铁道通信信号(2020年8期)2020-02-06
作文小学中年级(2019年10期)2019-11-04
蔬菜(2018年5期)2018-05-17
林业与生态(2017年2期)2017-03-01
骏马(2016年6期)2017-01-09
杂草学报(2015年2期)2016-01-04
天津诗人(2014年4期)2014-11-14

- Zoological Research的其它文章
- A phylogeny of the Tylototriton asperrimus group (Caudata: Salamandridae) based on a mitochondrial study: suggestions for a taxonomic revision
- 体外培养猕猴穆勒细胞经诱导可表现出视网膜前体细胞特征
- 体细胞核移植生产绵羊转hALR基因囊胚
- 上海市南汇东滩围垦后海岸带湿地冬春季水鸟生境选择
- 云南新平哀牢山西黑冠长臂猿分布与群体数量
- Histological and immunocytochemical study of deferens ducts in the Chinese rat snake (Zaocys dhumnades)