
新疆扁桃F1群体抗寒性研究
2012-08-08 07:10杨晓静徐崇志
塔里木大学学报 2012年3期
杨晓静 徐崇志
(塔里木大学植物科学学院,新疆阿拉尔843300)
扁桃(Amygdalus communis L.),维吾尔语巴旦木姆,又名巴旦木,属于蔷薇科(Rosaceae),李亚科(Prunoideae),扁桃属(Amygdalus),也为有人将其归为为李属(Prunus)植物,落叶乔木或小乔木[1]。扁桃是一种优良的木本油料和干果树种,也是新疆名、特、优干果之一,其栽培面积、产量和贸易量居世界干果首位[2-4]。扁桃仁香甜可口,富含蛋白质、脂肪、矿物质元素、维生素及酶类,是高级的营养食品;扁桃仁可用于治疗糖尿病、高血压、癌症、冠心病、气管炎、神经衰弱、小儿佝偻病等病症,具有极高的药用价值[5]。扁桃容易贮藏,便于运输,国内外市场需求量大,售价较高,具有广阔的市场前景。扁桃适应性强,耐旱、耐贫瘠、耐热、耐盐碱,特别适合在新疆发展,是南疆干旱荒漠及绿洲地区生态建设的优良树种[6]。扁桃原产于中亚细亚和非洲北部山区,我国新疆也是扁桃的次生原产地之一。扁桃在我国具有悠久的栽培历史,目前我国扁桃栽培主要集中在新疆的南部的疏勒、疏附、英吉沙、叶城、泽普、和田等地。但长期以来南疆的扁桃未得到应有的重视和发展,多处于半野生、半栽培状态,一些优良的类型或株系面临自生自灭的危机,相关方面的研究更少。对于目前扁桃的主栽区普遍面临的问题是产量始终处于低水平,究其原因,主要是新疆扁桃花期正值气温变化剧烈的季节,扁桃的花芽、花朵、幼果常因异常低温、寒流或大风降温天气而产生冻害,造成低产[7]。
本研究拟通过对大田扁桃枝条冻害情况调查及实验室内抗寒性相关生理生化指标(电解质渗出率、MDA含量、Pro含量、POD活性、CAT活性、SOD活性)的测定,并对测定结果进行主成份分析法进行综合分析,来探究4种新疆扁桃的自然杂交后代F1群体的抗寒性,从而从中筛选出抗寒性最强的单株或杂交组合,为探讨扁桃的抗寒性机理提供理论参考,旨在为解决新疆扁桃越冬过程中树体受冻和春季抽干的问题提供理论支持,同时也为扁桃种质资源的引进和大面积推广提供科学理论依据,从而为新疆扁桃产业化发展解决关键问题提供帮助。
1 材料与方法
1.1 材料
试验所需的枝条分别于2011年12月6号、12月22号、2012年1月6号、1月22号、2月7号从阿拉尔市塔里木大学园艺试验站扁桃种质资源圃内剪取,试验中涉及4个品种分别为莎车-1、莎车-2、莎车-9、莎车-13,后面涉及到其分别用S-1、S-2、S-9、S-13表示。在每个品种F1后代中健壮的植株上选择生长健壮、发育正常、充分成熟的1年生休眠枝,尽量在选定植株的不同方位不同层次选取粗细均匀的枝条,每个品种剪取12~16根,剪取的枝条长30~40 cm,而后分别贴好标签,装入聚乙烯袋内带回实验室。在实验室内用自来水冲洗,用蒸馏水冲洗3次后,用干净干燥的纱布擦干水分,分装于塑料袋中,置于冰箱(0℃~4℃)中保存备用。
1.2 方法
1.2.1 田间扁桃春季枝条抽干调查的方法
每个品种F1群体随机选取30株,于不同方向(东西南北)选取上下部、内外的枝条进行测算,调查其抽条情况;用观察法对扁桃抽条情况进行分级,具体分级标准为:0<受冻枝量/总枝量<25为Ⅰ级,25<受冻枝量/总枝量<50为Ⅱ级,50<受冻枝量/总枝量<75为Ⅲ级,75<受冻枝量/总枝量为Ⅳ级。参照王有科等[8]在引种扁桃抗寒力及冻害成因分析中的调查方法来对供试的新疆扁桃F1群体的抗寒性进行分级评价。具体方法为将调查数据按下式转化为越冬抽条指数:

据此对抗寒性进行分级评价:高抗抽条0~12.5%;中抗抽条12.6%~50%;易抽条50%~75%;极易抽条75%以上。
1.2.2 内抗寒性相关生理生化指标的测定方法
用电导率仪法[9]测枝条电解质渗出率;丙二醛(MDA)的含量用硫代巴比妥酸(TBA)显色法[10]测定;脯氨酸(Pro)的含量用磺基水杨酸提取茚三酮比色法[10]测定;过氧化物酶(POD)活性用愈创木酚法[11]测定;过氧化氢酶(CAT)活性用高锰酸钾(KMno4)滴定法测定;超氧化物歧化酶(SOD)活性用氮蓝四唑(NBT)还原法[12]。
1.2.3 数据统计与分析
数据统计及图表制作用Mcrosoft Excel,结果分析用DPS软件进行。
2 结果与分析
2.1 采样地采样期间气温情况
自2011年入冬后,阿拉尔垦区从12月中旬开始,旬平均气温持续偏低,旬平均气温偏低幅度分别为1.1℃、2.3℃、2.9℃、1.8℃、6.3℃,1月下旬旬平均气温偏低幅度与往年相比较大,低温长达50 d。极端最低气温达到-24.0℃,居近30年来首位。同时日最低气温达到了-21.6℃以下的日数持续了6 d,-24.0℃以下为3 d。具体变化情况如图1所示:
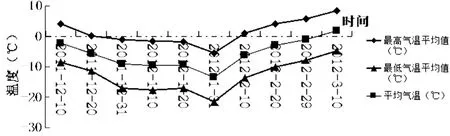
图1 采样期间采样地气温变化
2.2 大田扁桃枝条抽条情况调查结果
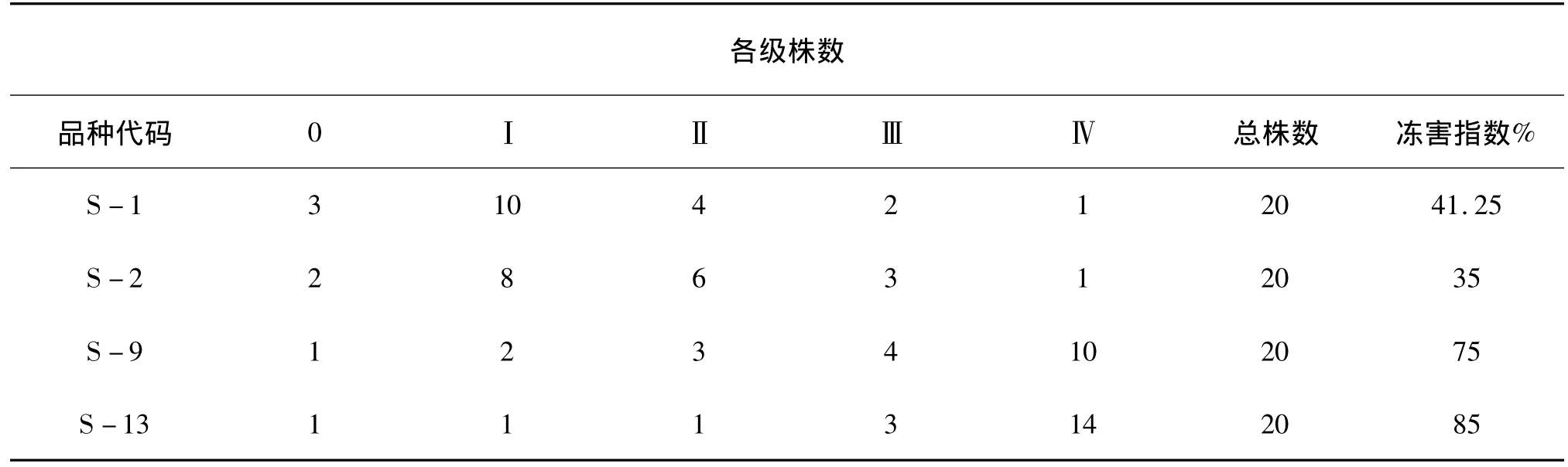
表1 大田扁桃枝条抽条情况调查结果
由表1春季抽干的调查可知,S-1的F1群体的抗抽条能力最强,S-2的其次,比前者稍弱些,抽条指数略大,这两者均属中抗抽条;S-9与S-13的抗抽条能力较差,均属极易抽条类型。
2.3 抗寒性相关指标的测定结果
2.3.1 新疆扁桃F1群体枝条电解质渗出率变化
目前在果树抗寒性研究中外渗电导率法是一种较为常用的方法。抗寒性强的品种,其电解质渗出率较低。由图2可知,在自然降温过程中,随气温的变化4个新疆扁桃自然杂交后代休眠枝的电解质渗出率变化趋势较一致,均为前期较缓慢增加而后期较快增加,在达到高峰后又迅速下降,形成一个山峰状,与该期的气温变化情况相反。但在整个过程中S-1与S-2的F1群体的电解质渗出率较为接近,而前者稍大于后者,且均小于S-9与S-13的。在整个过程中,S-13的电解质渗出率均为最大,S-9其次,且两者在1月中下旬气温快速下降过程中电解质渗出率快速上升,表明这两者抗寒性较弱。从图2可推知测试的几个新疆扁桃自然杂交F1后代的抗寒性表现为S-2>S-1>S-9>S-13,这一结果与大田扁桃抽条情况的调查结果相符。
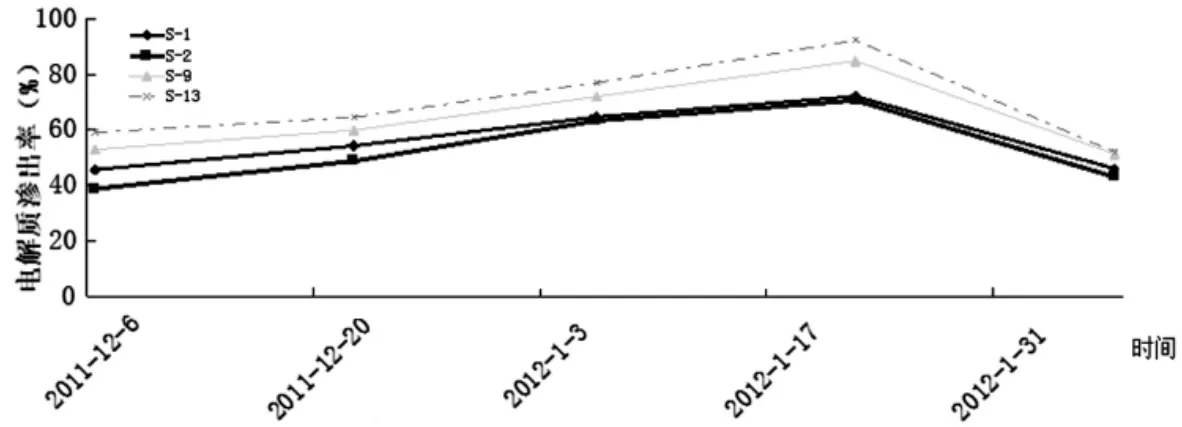
图2 自然降温过程中电解质渗出率变化
2.3.2 新疆扁桃F1群体枝条MDA含量变化
从图3可知,自然降温过程中供试的4个扁桃品种自然杂交后代枝条的丙二醛(MDA)含量的变化趋势与枝条的电解质渗出率的变化趋势基本相似,差别不大,均为先增大后减小。进一步比较发现在整个过程中MDA含量存在S-13>S-9>S-1>S-2这一现象,与电解质渗出率的情况一样。但在整个过程中S-2的MDA含量变化最为缓和,S-1的稍大些,S-13的表现的最为激烈,且S-2的MDA含量最小为1.765,温度最低时的最大值也仅为4.065,增幅为130.3%,而S-13的MDA含量则由4.048增大到14.98,增大了2.7倍。丙二醛是脂质过氧化作用的主要产物,其在组织中含量的多少反映了脂质过氧化作用的强弱,因此测定MDA的积累可以了解植物细胞受伤害的程度。MDA含量高说明植物的抗寒性弱,反之,则抗寒性强。故而由图3可推知整个过程中供试扁桃的抗寒性随气温的下降而减弱而后气温回升又增强,且抗寒性在整个过程中始终存在S-2>S-1>S-9>S-13,这一结果与电解质渗出率推出的结果相同,同时也印证了田间调查结果。
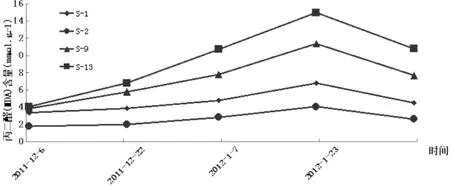
图3 自然降温过程中新疆扁桃F1群体枝条MDA含量变化
2.3.3 新疆扁桃F1群体枝条Pro含量变化
游离脯氨酸是渗透胁迫下易积累的一种氨基酸,也是一种重要的渗透调节物质,具有稳定细胞蛋白质结构、保护细胞内大生物分子和保持氮含量的作用。脯氨酸在植物抗冻中具有重要作用,低温逆境往往伴随着脯氨酸含量的增加,其含量高低与植物抗寒性密切相关,越是抗寒性强的品种增加的倍数越高。一般认为,植物组织中游离脯氨酸含量随温度下降而增加[14]。由图4可知,在自然降温过程中,4个供试的扁桃品种的自然杂交后代的脯氨酸含量的变化情况也大同小异,均表现为随着气温降升而增减,从1月中旬到2月上旬数值较大,而12月上旬到一月上旬数值相对较小。供试材料最初的Pro含量相当,相差不大,而其后的变化却不尽相同,表现为而后S-2和S-1的以较大的速率增加,然后增长势头减弱,而后增长速率又有所增大,达到最大值后S-2的含量仍有所增大,S-1的则减小;与S-1和S-2相比,S-9和S-13在其后的变化表现的较为一致。其中S-1与S-2的增幅显著,分别从最初的52.418μg/(g·Fw)和57.698μg/(g·Fw)增至最大值198.349μg/(g·Fw)和226.232 μg/(g·Fw),上升了2.76倍和3.01倍;而S-13的增幅较小,从35.864μg/(g·Fw)增大到75.949 μg/(g·Fw)仅增加了1.12倍。
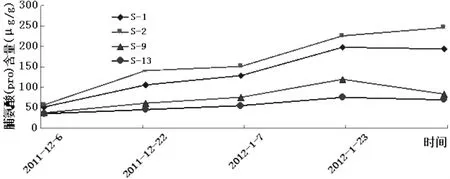
图4 自然降温过程中新疆扁桃F1群体枝条pro含量变化
2.3.4 新疆扁桃F1群体枝条保护酶活性变化
低温使膜上的酶系统受到破坏,使酶活性下降,利用酶活力变化可作为植物抗寒性鉴定指标。SOD是一种含金属的酶,是植物体中最重要的清除自由基的酶之一,主要功能清除超氧阴离子自由基使其发生歧化反应。CAT在植物体中是一种功能较专一的酶,普遍存在于植物的所有组织中,其活性与植物的代谢强度及抗寒、抗病能力有一定的联系[13],它的主要功能是分解H2O2。POD是一种功能较复杂、种类较多的酶,一般认为它的功能与木质素的合成有关,并与细胞的抗病相联系,POD的主要功能是催化H2O2分解,氧化其底物所产生的过氧化物。POD与CAT协同清除H2O2。陈新华等[14]试验表明,4个甜樱桃品种的SOD、POD酶活性随温度的降低而升高,随气温的回升而降低,酶活性高的品种表现出较强的抗寒性,反之,抗寒性较弱。
由图5可看出,4个新疆扁桃品种自然杂交后代枝条的POD活性在越冬过程中随气温“先下降后回升”的变化趋势也呈现出相同的“先减弱后增强”的变化趋势。在最低温出现的1月22号左右,4个品种的枝条POD活性也降为最低值,分别是S-1为82.417 U/(g·min)、S-2为100 U/(g·min)、S-9为50.667 U/(g·min)、S-13为23.5 U/(g·min)。与最初的POD活性相比S-13的降幅最显著从最初的127.75 U/(g·min)降到23.5 U/(g·min),减小了104.25 U/(g·min),降低了8成多。整个过程中S-2的POD活性始终为最高,S-1其次,且两者的变化趋势几近同步,S-9和S-13虽有交叉,但总体上为S-9的大于S-13的。由此可知,单从POD活性这与方面来说,S-2的杂交后代抗寒性为最强,其次为S-1,S-13的抗寒性差。
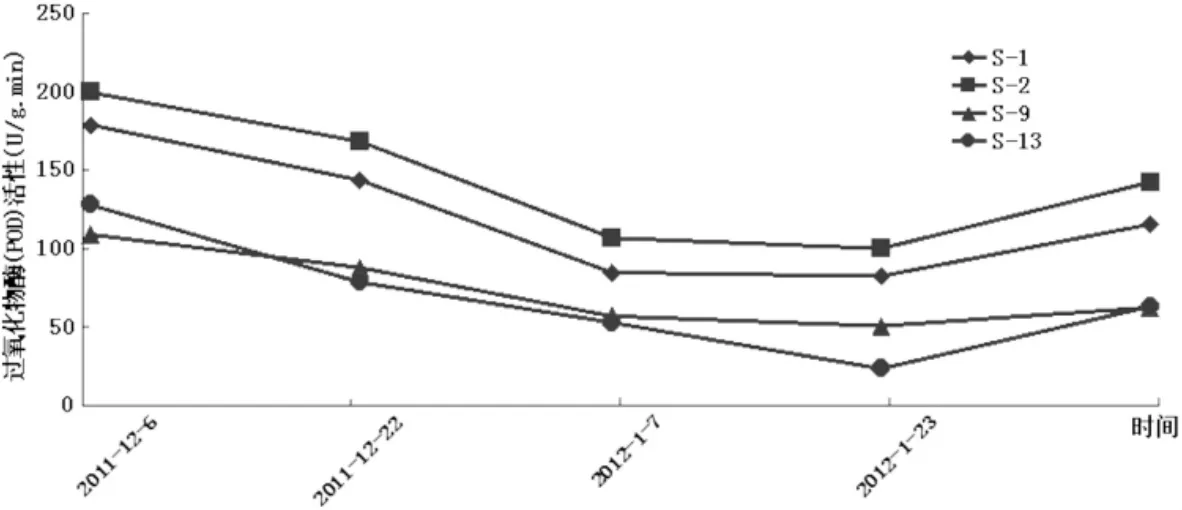
图5 自然降温过程中新疆扁桃F1群体枝条过氧化物酶(POD)活性变化
由图6可知,在自然降温过程中S-1、S-2、S-13的自然杂交后代群体枝条的过氧化氢酶(CAT)活性整体表现为先增后减的态势,其中S-13在12月下旬CAT活性有所下降,而S-9在达到峰值后并未下降反而有小幅增加。就总体而言,4个品种扁桃的自然杂交后代的过氧化氢酶活性均在气温最低的1月下旬达到较大值,而后随气温回升其活性有不同程度的下降。在整个过程的不同阶段中S-1与S-2过氧化氢酶活性的变化幅度较其它两者要剧烈些,尤其是在气温下降相对较剧烈的1月份,表明S-1与S-2的杂交后代应对低温变化的能力较S-9与S-13要强些。同时由图6可较直观得看出,越冬过程中4个供试品种的F1群体的CAT活性关系为S-2>S-1>S-9>S-13,故而可推知它们的抗寒性也存在相同的关系。
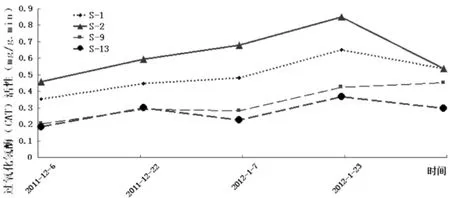
图6 自然降温过程中新疆扁桃F1群体枝条过氧化氢酶(CAT)活性变化
由图7可知,自然降温过程中,供试的4个品种的新疆扁桃F1群体的超氧化物歧化酶(SOD)活性整体呈现为“上升—下降—上升—下降”的变化,呈现为“M”型,这一结果与郑强卿[16]等在自然降温对杨树枝条抗寒性生理生化指标变化的影响中的实验结果相似。而且S-1与S-2在越冬过程不同时期的变化较为一致,S-9与S-13的变化也较一致,但总体来讲,整个过程中S-2的SOD活性始终最大,S-1的其次,S-9的稍大于S-13。
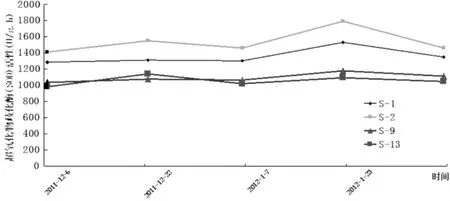
图7 新疆扁桃F1群体枝条过氧化物歧酶(SOD)活性变化
2.4 各个采样时期供试4个品种新疆扁桃F1群体抗寒性综合评价
依据试验对每个采样期所测的抗寒性指标,电解质渗出率、MDA含量、Pro含量、过氧化物酶活性、过氧化氢酶活性、超氧化物歧化酶活性等6个指标值进行主成分分析,计算出特征值和贡献率,根据累计贡献率选取主成分,数据处理在DPS软件上运行,主成分分析结果如表2和表3所示。
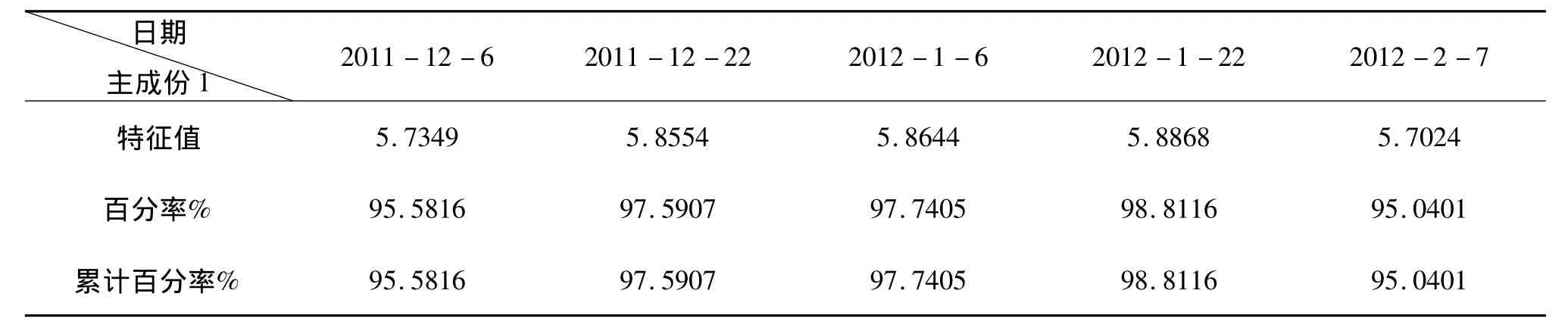
表2 不同采样期主成份1的特征值及累计百分率

表3 不同采样期各个指标在第一主成份上的权重
结合表2和表3可知,对从2011年12月6日到2012年2月7日的5次采样而言,不同采样时间供试的4个品种新疆扁桃F1群体抗寒性主成分分析综合评价结果较相似,均表明在6个抗寒指标中电解质渗出率与MDA含量表现出极显著的相关性,而与其它四种指标之间表现出显著或极显著的负相关性,其它四种指标间存在显著或极显著正相关,如在先前的4次采样期CAT活性均与POD活性、SOD活性、Pro含量表现出极显著正相关,仅是最后一次表现出显著的正相关性;且均是第一个主成份因子的累计贡献值就超过85%,且均在90%以上;又据主成份因子的载荷矩阵可推知第一主成分所包括的在抗寒指标均为Pro含量、SOD活性、CAT活性、POD活性,且四种指标在第一主成分中所占的权重相当,每个指标所占权重岁采样时间不同变化不大。这就说明在对不同采样时间采取的供试材料抗寒性进行综合评价时均可只对第一主成分所包含的4个指标进行分析、比较。据此可推出如下结果,对于不同的采样时间,供试的4个品种新疆扁桃F1群体抗寒性均存在S-2>S-1>S-9>S-13排序,这样的结果和田间枝条抽干结果相符,也与上述单指标评价结果相吻合,如表4所示:

表4 不同采样时间4个品种新疆扁桃F1群体抗寒性综合评价结果
3 讨论与小结
在本研究结果显示供试的4个品种扁桃的杂交后代的电解质渗出率均随气温的下降而上升,说明在气温下降过程中植株的生物膜发生了相变,膜通透性增大,造成细胞内容物外渗。然而不同品种的上升情况并不相同,比较而言上升速率为S-13>S-9>S-1>S-2,且S-1与S-2的增加速率较接近,均较缓和,单从这一方面来看,S-2的F1群体耐低温能力最强,其次为S-1,再为S-9,S-13最弱。MDA可以作为研究植物抗寒性的一个重要指标。本实验结果显示,与电解质渗出率相似在越冬过程中4个新疆扁桃品种的自然杂交后代群体的MDA含量随气温下降而上升,气温回升时有所下降,且变化幅度为S-13最为剧烈,尤其是在气温快速下降的一月下旬,而S-1与S-2在同期的变化却相对较缓和。这表明S-13在此阶段所受低温的伤害程度较大,S-1与S-2相对较小,而S-9介于两者之间。通过本次研究可发现,供试的4个品种的新疆扁桃F1群体的脯氨酸(Pro)含量也为随越冬过程中气温的降升而增减,呈山峰状,与电解质渗出率和丙二醛的变化趋势相同,而在数值上与前两项的结果恰相反。且在整个过程中S-2与S-1的脯氨酸含量变化对气温的变化较为敏感,S-13表现的最为迟钝,这表明S-1与S-2的杂交后代体内应对低温的机制较灵活,而S-13恰恰相反。
本次的研究结果显示,供试材料的POD活性均大致表现为与越冬过程中气温的变化相似的变化,为先减弱在气温最低时达谷值而后随气温回升也增大。这一结果与杨晓宇等[16]扁桃枝叶生理指标与抗寒性相关性研究中结果相同,但与郑强卿[16]等自然降温对杨树枝条抗寒性是生理指标变化的影响的研究结果却有差异,这可能使因为树种不同的原因,这也在一定程度上说明仅仅应用某一指标来衡量复杂的抗寒性生理过程是不科学、不全面的。
本实验结果表明,供试材料的CAT活性表现为随温度下降而增强随温度回升而下降。这一结果与郑强卿[16]等自然降温对杨树枝条抗寒性是生理指标变化的影响的研究结果相同,与一般相关理论相符。
此次实验结果表明,在自然降温过程中4个供试新疆扁桃品种的杂交后代的SOD活性变化均呈现为“M”型,且整个过程中其变化幅度均不大,变化均较缓和。越冬过程中S-2杂交后代的SOD活性始终处于较高的水平,S-1的也保持较高的水平,而S-13与S-9却始终处于较低水平。单从这一方面可推知S-2抗寒性最强,S-1其次,S-13最弱,S-9弱于前两者强于后者。
本实验通过对不同采样期测试指标进行主成分分析得出可用来综合评价供试材料抗寒性的第一主成分不论采样时间如何均是有Pro含量、SOD活性、CAT活性、POD活性4个指标组成,且它们之间均存在显著或极显著的正相关性;四者在第一主成分因子所占的权重也相当,每个指标随采样时间变化在主成分因子权重变化也不大。据此法评比出的抗寒性强弱顺序与田间调查结果相符也与单指标简单评价结果相吻合,均为S-2>S-1>S-9>S-13。
[1] 龚鹏,李秀霞,李增萍,等.低温胁迫对扁桃枝条细胞系统和渗透调节物质的影响[J] .新疆农业科学,2007,44(5):561-566.
[2] 武彦霞,王占和,何勇,等.我国扁桃的生产现状及发展前景[J] .山西农业科学,2005,33(2):20-22.
[3] 王有科,席万鹏,郁松林,等.引种扁桃抗寒力及冻害成因分析[J] .干旱地区农业研究,2006,24(5):126-129.
[4] 帕塔木,布早拉木,肖开提,等.巴旦木栽培技术[J] .新疆农业科学,2007(2):21.
[5] 王彩云.扁桃的栽培利用及其发展前景[J] .河北林果研究,2008,23(1).
[6] 谢军,耿文娟,何峰江,等.以电导法配合Logistic方程测定6种扁桃枝条的抗寒性[J] .新疆农业大学学报,2011,34(4):32-35.
[7] 徐崇志.扁桃自交不亲和强度差异的机理初探[D] .西安,西北农林科技大学,2007.
[8] 王有科,席万鹏,郁松林,等.引种扁桃抗寒力及冻害成因分析[J] .干旱地区农业研究,2006,24(5):126-129.
[9] 李合生.植物生理生化实验原理和技术[M] .北京:高等教育出版社,2000.
[10] 王学奎.植物生理生化实验原理和技术[M] .北京:高等教育出版社,2000.
[11] 孙群.植物生理学实验指导[M] .北京:西北农林科技大学教材,2002.
[12] 高俊风.植物生理学实验技术[M] .世界图书出版公司,2000.
[13] 李宁华,李志辉.桉树耐寒性与超氧化物歧化酶关系研究[J] .中南林学学报.2000,20(3):63-66.
[14] 陈新华,郭宝林,赵静,等.甜樱桃休眠枝条SOD酶、POD酶活性变化[J] .安徽农业科学,2009,37(11):4958-4959.
[15] 杨晓宇,田建宝.扁桃枝叶生理指标与抗寒性相关性研究[D] .2007.
[16] 郑强卿,李铭,李鹏程,等.自然降温对杨树枝条抗寒性生理指标变化的影响[J] .学术园地.2011,12.
猜你喜欢
热带作物学报(2022年5期)2022-06-01
黑龙江粮食(2021年10期)2021-12-13
中国果业信息(2021年7期)2021-12-01
童话世界(2019年31期)2019-11-25
小学生必读(低年级版)(2019年5期)2019-08-30
中成药(2018年6期)2018-07-11
诗林(2016年5期)2016-10-25
中国果菜(2016年9期)2016-03-01
中国马铃薯(2015年5期)2016-01-09
园艺与种苗(2015年10期)2015-02-27
