
新疆南疆猪繁殖与呼吸综合征病毒毒株分型鉴定
2012-08-08 07:10崔浩然刘永宏焦海宏张志峰刘俊峰
塔里木大学学报 2012年3期
崔浩然 刘永宏 赵 丽 焦海宏 张志峰 刘俊峰
(塔里木大学动物科学学院/新疆生产建设兵团塔里木畜牧科技重点实验室,新疆阿拉尔市,843300)
猪繁殖与呼吸综合征(Porcine Reproductive and Respiratory Syndrome,PRRS)是由猪繁殖与呼吸综合征病毒(Porcine Reproductive and Respiratory Syndrome virus,PRRSV)引起猪的一种高度接触性传染病,不同年龄、品种和性别的猪均可感染,但以妊娠母猪和1月龄以内的仔猪最易感,该病以母猪的流产、死胎、弱胎、木乃伊胎以及仔猪的呼吸困难、败血症、高死亡率等为主要特征[1]。
目前,PRRS严重威胁着全球养猪业和世界公共卫生,给经济带来巨大的损失,仅美国因该病每年至少要花费5.6亿美元[2],权威组织评价每头生长猪出栏前因PRRS花费5.60美元~7.60美元。
2006年春夏之交,我国从南向北25个以上省、市、自治区暴发以高热、高发病率、高死亡率和低治愈率为特征的“猪高热病(porcine high fever disease,PHFD)”,至少200多万头猪发病,死亡40万头之多,2007年7月田克恭等及农业部[3]将其确诊为高致病性猪繁殖与呼吸综合征(Highly Pathogenicity Porcine Reproductive and Respiratory Syndrome,HP-PRRS),JXA1株为HP-PRRSV代表株,属于美洲型。并将HP-PRRS确定为中国一类动物疫病[3],普通毒力的PRRS确定为中国二类动物疫病。
PRRSV为单股正链RNA病毒,根据抗原性差异将各地PRRSV分离株分为两个型,即欧洲型(Ⅰ型)和美洲型(Ⅱ型),原型株分别为LV株和ATCC VR-2332株[4]。PRRSV基因组全长约15kb,至少编码9个相互重叠的开放阅读框架(Open reading frame,ORF),即ORF1a、ORF1b、ORF2a、ORF2b、ORF3、ORF4、ORF5、ORF6和ORF7[5]。ORF5、ORF6和ORF7编码三个主要的结构蛋白,分别为被膜蛋白GP5、膜蛋白M和核衣壳蛋白N[5]。和欧洲型原型株LV株相比,北美型N蛋白核苷酸同源性仅约63%,氨基酸同源性仅约59%的水平[6]。如此相对高的偏离原因为多个核苷酸置换、插入和缺失引起,使北美型N蛋白比欧洲型LV株少了5个氨基酸[7] 。
考虑新疆南疆周邻多国的地理位置、戈壁沙漠地域较广和干旱气候等特点,以及PRRSV高变异性[7]和抗体依赖性增强[8]等特点,新疆南疆毒株可能有其自身的、不同于国内外其他毒株的特点。本研究拟针对新疆南疆PRRSV ORF7部分因序列进行遗传演化分析,讨论所得毒株类型,为新疆南疆有效防制PRRS提供依据和技术储备。现结果报告如下:
1 材料与方法
1.1 材料
1.1.1 病料
采自新疆南疆阿克苏和库尔勒地区发病自然死亡猪只。阳性对照为上海海利/蓝耳病活疫苗/高致病性猪繁殖与呼吸综合征活疫苗(HuN4-F112株)。
1.1.2 主要试剂盒
Trizol invitrogenTM(Code No.:15596-026),购自Invitrogen生物工程有限公司;TaKaRa RNA PCR Kit(AMV)Ver.3.0(Code No.:DRR019A),Premix Taq Version2.0(Code No.:D331A),购自宝生物工程(大连)有限公司;TIANgel Midi Purification Kit(Code No.:DP209),TIANprep Mini Plasmid Kit(Code No.:DP103),pGM-T克隆试剂盒(Code No.:VT202),DNA markerⅡ(Code No.:MD102),DNA markerⅢ(Code No.:MD103)均购自天根生化科技(北京)有限公司;等。
1.1.3 PRRSV参考毒株
PRRSV参考毒株均下载于NCBI数据库,毒株名称、大小和GenBank登录号见表1。
1.1.4 引物
按照引物设计原则,利用Primer Premier5.0分子生物学软件,以GenBank数据库中保守的PRRSV基因序列为参照设计特异性引物,跨度300bp左右,引物由生工生物工程(上海)有限公司合成。具体序列如下:上游引物:5'-AATGGCCAGCCAGTCAATCA-3';下游引物:5'-GAATCAGGCGCACTGTATGA-3'。

表1 参考毒株
1.1.5 仪器
PCR仪(TC-5000,Bibby scientific Ltd),小型高速冷冻离心机(R134a,Hermetically sealed refigeration system),电泳仪(DYY-12,北京市六一仪器厂),紫外分析仪(JY02S,北京君意东方电泳仪设备有限公司),等。
1.2 方法
1.2.1 总RNA提取
从病死猪肺脏等混合组织中提取总RNA,按Trizol invitrogenTM试剂盒说明书操作。
1.2.2 RT-PCR反应
总RNA按照TaKaRa RNA PCR Kit(AMV)Ver.3.0试剂盒,利用自行设计的特异性引物,进行一步法扩增ORF7基因片段,退火温度54.1℃。设立阳性对照(弱毒疫苗)和阴性对照(蒸馏水)。
1.2.3 PCR产物纯化回收
取ORF7基因RT-PCR扩增产物电泳切胶后,按照TIANgel Midi Purification Kit试剂盒说明进行回收纯化。
1.2.4 纯化产物连接T载体
胶回收纯化目的产物与pGM-T载体连接,按照pGM-T克隆试剂盒说明操作。
1.2.5 转化DH5α感受态细胞、鉴定、摇菌及提取质粒送测序
将连接产物转化自制的感受态细胞DH5α,进行蓝白斑筛选。白色菌落利用Premix Taq?Version2.0试剂盒进行菌落PCR鉴定,挑取多个PCR阳性菌落摇菌,按照TIANprep Mini Plasmid Kit试剂盒提取质粒DNA,标记为XJNJ-4-1、XJNJ-4-3、XJNJ-5-2、XJNJ-5-3、XJNJ-9-1、XJNJ-9-2、XJNJ-13-1、XJNJ-13-2、XJNJ-14-1、XJNJ-14-3、XJNJ-18-1、XJNJ-18-2、XJNJ-19-2和XJNJ-19-3,送生工生物工程(上海)有限公司测序,测序引物为T7。
1.2.6 序列分析
用DNAMAN和DNAStar生物学软件进行序列比对和遗传演化分析。
2 结果与分析
2.1 ORF7基因RT-PCR扩增结果
利用ORF7基因特异性引物,对发病疑似PRRS猪组织病料总RNA进行RT-PCR扩增,将扩增产物进行1%琼脂糖凝胶电泳。结果显示,14例PRRSV阳性,分别命名为PRRSV XJNJ-4-1、XJNJ-4-3、XJNJ-5-2、XJNJ-5-3、XJNJ-9-1、XJNJ-9-2、XJNJ-13-1、XJNJ-13-2、XJNJ-14-1、XJNJ-14-3、XJNJ-18-1、XJNJ-18-2、XJNJ-19-2和XJNJ-19-3株,得到的目的基因片段与预期的大小一致,对照成立(图1)。
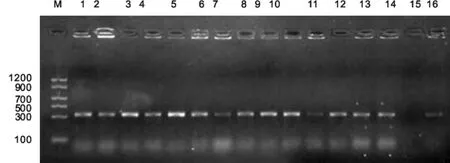
图1 ORF7基因RT-PCR扩增结果
2.2 ORF7基因核苷酸序列比较分析
RT-PCR阳性产物胶回收纯化,连接pGM-T载体,转化自制的感受态细胞DH5α,白色菌落进行菌落PCR鉴定,挑取PCR阳性菌落摇菌,提取质粒DNA,送生工生物工程(上海)有限公司测序,获得了14个ORF7基因部分片段序列。
本研究获得的14个新疆南疆PRRSV毒株与6个国内代表毒株(2006年中国暴发HP-PRRS代表株JXAl,中国另外一个HP-PRRSV毒株HuN,中国自然弱毒疫苗株R98,JXA1致弱株JXA1 P80和中国代表株CH-1a及其致弱株CH-1R)和3个国外代表株(进口美洲型疫苗株MLV,美洲型代表毒株VR2332和欧洲型代表毒株LV)相应的ORF7基因序列进行序列比对分析,结果显示本研究获得的14株、国内6株参考株以及进口美洲型疫苗株MLV和美洲型代表毒株VR2332都不存在9个碱基的缺失,只有欧洲型代表毒株LV该位置有9个碱基的缺失,结果与预期设计相符(图2)。

图2 ORF7片段中核苷酸的缺失
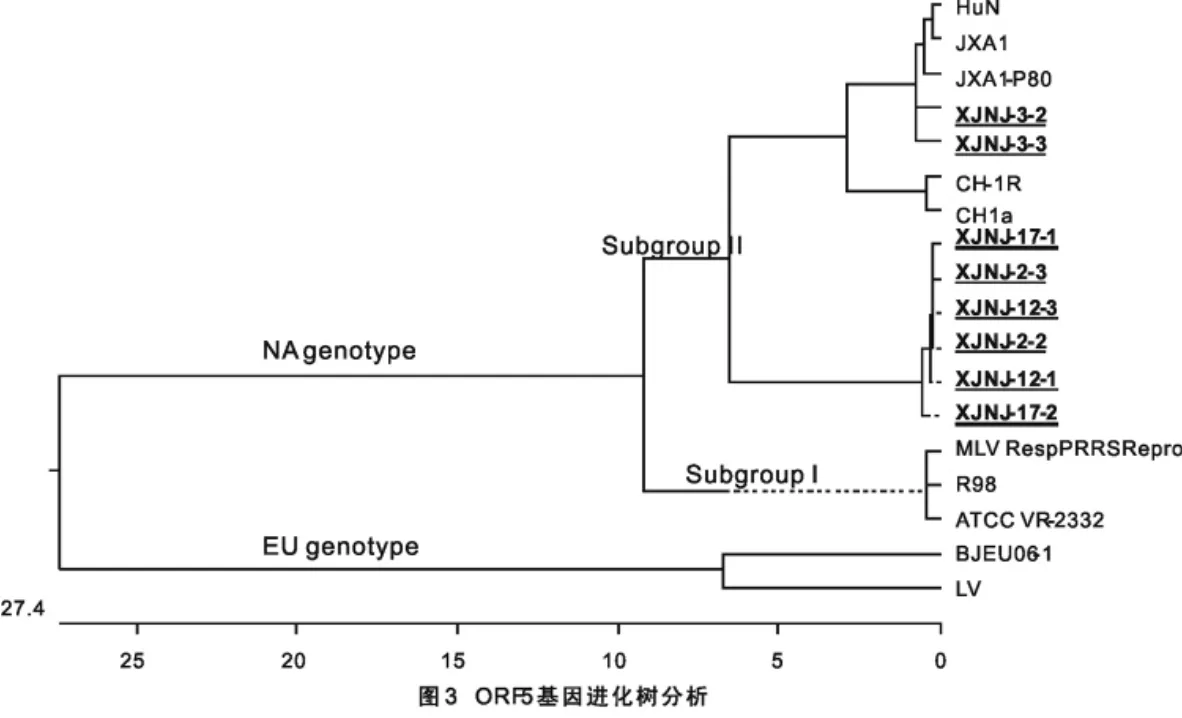
2.3 ORF7基因核苷酸序列遗传进化树
将本研究获得的14个PRRSV毒株ORF7基因部分片段,与参考的9株PRRSV截取相应位置长度的ORF7基因序列,进行核苷酸比对,绘制了23个毒株的发育进化树。如图3所示,本研究的14株均与中国代表株CH-1a及其致弱株CH-1R、2006年中国暴发HP-PRRS代表株JXAl及其致弱株JXAl P80、中国另外一个高致病性毒株HuN、中国自然弱毒疫苗株R98、进口美洲型疫苗株MLV和美洲型代表毒株VR2332在同一个大的分支,均不与欧洲型代表株LV在一个分支。另外,XJNJ-5-2和XJNJ-5-3株与2006年中国暴发HP-PRRS代表株JXAl及其致弱株JXAl P80和中国另外一个高致病性毒株HuN在同一个小的分支内。
3 讨论
1996年郭宝清等[9]首次从北京流产胎儿体内分离到PRRSV,从而证实PRRS已在我国存在。随后,该病在中国各省市相继均有报道。2006年,中国猪养殖场由南向北暴发一次毁灭性灾难-HPPRRS。中国新疆其他省市对PRRS研究较多,新疆对该病的研究属于探索阶段。新疆地处中国西北边陲,周边与8国接壤,尤其在俄罗斯流行PRRSV欧洲型毒株,与中国流行的美洲型毒株有明显差异。另外,中国台湾[10]、中国内地检疫口岸以及内地一些地区有欧洲型毒株的报道。之前的研究,两洲分离株属于两个不同的基因型,各洲野外分离株和各洲起源株有高度的亲缘关系,亚洲国家PRRSV遗传学关系显示为美洲型[11]。近年来,PRRSV的流行早已打破原来的地域限制,在亚洲和北美均有欧洲型PRRSV流行的报道,欧洲也出现了野生型PRRSV的美洲株。所以,有必要研究分析清楚新疆南疆PRRSV流行毒株的类型及特点,以便于更好的预防和控制新疆南疆PRRS。对于PRRSV的相关基因的变异性研究是十分必要的,在此基础上进行合理的种毒株选择,疫苗设计和制备,以便提出有效的防控措施和防制计划。
PRRSV的ORF7基因,是除ORF6基因以外保守性最高的编码结构蛋白的基因[12],美洲型毒株间氨基酸的同源性为96%~100%,欧洲型毒株间氨基酸同源性为94%~99%,但欧洲型与美洲型毒株之间N蛋白氨基酸的同源性仅为63%,氨基酸同源性仅为59%[6]。本研究针对相对保守的ORF7基因,设计了可以同时扩增欧洲型和美洲型PRRSV毒株ORF7基因部分片段的特异性引物,扩增片段包括欧洲型毒株9个核苷酸片段的缺失区,针对此用于区分欧洲型和美洲型毒株。同时,本研究病料均采自新疆南疆发病疑似PRRS猪只,且均来自未免疫PRRS弱毒疫苗猪场,所以此次阳性结果表明该猪场存在PRRSV感染。本研究通过RT-PCR有14例PRRSV阳性,说明该组织样品来源猪场存在PRRSV感染。RT-PCR阳性产物克隆测序获得的14个ORF7基因片段,均没有该位置9个核苷酸的缺失。同时结合参考的9株PRRSV代表株,截取相应位置长度的ORF7基因序列,进行核苷酸序列比对得到的进化树显示,本研究的14株均与中国代表株CH-1a及其致弱株CH-1R、2006年中国暴发HP-PRRS代表株JXAl及其致弱株JXAl P80、中国另外一个高致病性毒株HuN、中国自然弱毒疫苗株R98、进口美洲型疫苗株MLV和美洲型代表毒株VR2332在同一个大的分支,均不与欧洲型代表株LV在一个分支。以上结果综合表明,本研究采集样品猪场感染了PRRSV美洲型毒株,同时说明这些猪场没有感染欧洲型毒株。
另外,进化树显示XJNJ-5-2和XJNJ-5-3株与2006年中国暴发HP-PRRS代表株JXAl及其致弱株JXAl P80和中国另外一个高致病性毒株HuN在同一个小的分支内,因为本研究病料均采自未免疫PRRS弱毒疫苗猪场,所以XJNJ-5-2和XJNJ-5-3株属于高致病HP-PRRSV毒株
至于更准确的结论,以及新疆南疆更多毒株的获得及毒株特点分析,会在将来的工作中逐步进行,从采样点全面、毒株数量多等方面考虑,最终监测新疆南疆PRRSV毒株类型及特点,以便及早的发现新毒株和新特点,提出更加有效的防制对策,减少因PRRS造成新疆南疆养猪业损失。
4 结论
本实验未检测到欧洲型毒株,获得的14个新疆南疆地区PRRSV毒株均属于美洲型毒株,其中2株属于美洲型高致病性HP-PRRSV毒株。
[1] Cho J G,Dee SA.Porcine reproductive and respiratory syndrome virus[J] .Theriogenology,2006,66(3):655-662.
[2] Neumann EJ,Kliebenstein JB,Johnson CD,et al.Assessment of the economic impact of porcine reproductive and respiratory syndrome on swine production in the United States[J] .Am Vet Med Assoc,2005,227:385-392.
[3] Tian K,Yu X,Zhao T,et al.Emergence of Fatal PRRSV Variants:Unparalleled Outbreaks of Atypical PRRS in China and Molecular Dissection of the Unique Hallmark[OL] .PLoSOne.2007,2:e526.DOI:10.1371/journal.pone.0000526.
[4] Wensvoort G.Lelystad virus and porcine epidemic abortion and respiratory syndrome[J] .Vet Res,1993,24:117-124.
[5] Dea S,Gagnon C A,Mardassi H,et al.Current knowledge on the structural proteins of porcine reproductive and respiratory syndrome(PRRS)virus:comparison of the North American and European isolates[J] .Arch Virol,2000,145:659-688.
[6] Meng X J,Paul P S,Halbur P G,et al.PHylogenetic analyses of the putative M(ORF6)and N(ORF 7)genes of porcine reproductive and respiratory syndrome virus(PRRSV):Implication for the existence of two genotypes of PRRSV in the USA and Europe[J] .Arch Virol,1995,140:745-755.
[7] Mardassi H,Mounir S,Dea S.Identification of major differences in the nucleocapsid protein genes of a Quebec strain and European strains of porcine reproductive and respiratory syndrome virus.[J] Gen Virol,1994,75:681-685.
[8] Yoon K J,Wu L L,Zimmerman JJ,et al.Field isolates of porcine reproductive and respiratory syndrome virus(PRRSV)vary in their susceptibility to antibody dependent enhancement of infection.[J] Vet Microbiol,1997,55(1-4):277-287.
[9] 郭宝清,陈章水,刘文兴,等.从疑似PRRS流产胎儿分离PRRSV的研究[J] .中国预防兽医学报,1996(2):1-4.
[10] Chang C C,Chung W B,Lin M W,et al.Porcine reproductive and respiratory syndrome(PRRS)in Taiwan I.Viral isolation.JChin Soci Vet Sci,1993,19:268-276.
[11] Tong GZ,Qiu HJ,Guo BQ,et al.A PRRSV isolated from the first outbreak in China is genetically related with North American genotype.Proceedings of the International Symposium on PRRS and Aujeszky's Disease,Ploufragan,France,June 21-24,1999,53.
[12] Meulenberg J J,Petersen den Besten A,de Kluyver E,et al.Molecular characterization of Ldystad virus[J] .Vet Microbiol,1997,55(1-4):197-202.
猜你喜欢
科学大观园(2022年2期)2022-01-23
小哥白尼(野生动物)(2021年12期)2021-03-29
歌海(2021年6期)2021-02-01
艺术家(2020年6期)2020-08-10
娃娃乐园·综合智能(2019年10期)2019-12-07
中成药(2018年2期)2018-05-09
中国猪业(2017年11期)2017-12-11
中国卫生(2016年7期)2016-11-13
小火炬·阅读作文(2014年2期)2015-03-11
特产研究(2014年4期)2014-04-10
