
干扰素-γ增强RAW264.7巨噬细胞对阿萨希毛孢子菌杀伤作用的机制研究
2012-10-20 05:53聂建巍杨蓉娅王文岭夏志宽李海涛
实用皮肤病学杂志 2012年1期
聂建巍,杨蓉娅,丛 林,王文岭,夏志宽,李海涛
近年来,由于免疫抑制剂以及广谱抗生素的广泛应用,器官移植、肿瘤的放疗、化疗以及侵入性治疗的增加,播散性毛孢子菌病的报道越来越多[1-3]。随着唑类和两性霉素B等抗真菌药物的大量应用,又使真菌耐药的问题日趋严重,给抗真菌治疗带来新的挑战[4]。中性粒细胞、单核巨噬细胞以及T淋巴细胞介导的细胞免疫反应在抵御病原体的入侵中起着重要的防御作用。其中,单核-巨噬细胞被认为是机体抗感染的主要防线。深部真菌感染与机体免疫防御和免疫状态之间有着密切的关系,而细胞因子在深部真菌感染发展过程中,起着重要的免疫调节作用,因此,免疫调节治疗近年来也逐渐成为深部真菌感染治疗的热点问题。干扰素-γ (IFN-γ)是辅助性T细胞(Th1细胞)分泌的一种细胞因子,本实验从细胞及分子水平探讨IFN-γ对小鼠巨噬细胞(Macrophage,Mφ)体外抗阿萨希毛孢子菌(Trichosporon asahii,T.asahii)活性的影响,以此来阐述IFN-γ在抗T.asahii感染中的作用机制。
1 材料与方法
1.1 细胞与菌株
小鼠单核/巨噬细胞系RAW264.7细胞株由中国协和医科大学基础医学细胞中心提供。T. asahii株为北京军区总医院全军皮肤病诊治中心-80℃保存的T. asahii临床分离株(编号:BZP07005),经API 20CAUX系统生化鉴定及DNA序列分析鉴定证实(AS2.2174)[5]。
1.2 主要试剂
小鼠重组 IFN-γ(PeproTech,1×107U/mg),快速瑞氏-吉姆萨染液(南京建成科技有限公司),胎牛血清(FBS)(杭州四季青生物工程材料有限公司),胰酶细胞消化液(0.25%Trypsin-EDTA Solution)(碧云天生物技术有限公司),Trixon-100(sigma公司),细胞培养基采用DMEM高糖培养基(Hyclone公司),NO检测试剂盒,超氧化物检测试剂盒(碧云天生物技术有限公司)。
1.3 方法
1.3.1 T.asahii传代、培养 将-80℃保存的T. asahii临床分离株转种于固体沙氏培养基,传代3次纯培养,挑取一接种环的T.asahii临床株接种于液体YPD培养基中,37℃、150r/min摇床培养8h,6000rpm离心,5min收集孢子,PBS(pH7.0)洗涤3次,然后用DMEM高糖培养基进行重悬,用血球计数板调整到1×106个 /ml。
1.3.2 Mφ的分离、培养 RAW264.7巨噬细胞用DMEM高糖培养基和10%FBS培养3天后,弃培养液,以胰酶-EDTA消化,接种于细胞培养瓶,37℃、5% CO2培养箱培养,传代2次后取对数生长期的RAW264.7细胞,1000r/min离心5min后,弃上清,用PBS洗2遍,用含10%FBS的DMEM培养基悬浮细胞,快速瑞氏-吉姆萨染液染色证实为巨噬细胞,台盼蓝染色计活细胞数>95%,计数、调整细胞浓度至1×105个/ml。
1.3.3 待测上清液的制备 96孔板,分为实验组及巨噬细胞对照组。每孔加巨噬细胞悬液100μl,3h后用预热的PBS洗2遍,继续向实验组每孔中加入终浓度分别为10、100、1000U/ml的IFN-γ,置37℃、5%CO2培养箱培养18h,弃旧液,用培养基洗2遍,每孔加T.asahii细胞悬液100μl,培养4h,取上清100μl,转移到另一96孔板,然后置于-80℃冰箱待检。
1.3.4 不同浓度INF-γ作用后巨噬细胞抗T.asahii活性测定 将上述96孔板剩余的上清液体系中每孔加入2%的TritonX-100 100μl,10min后混匀,从每孔吸出10μl悬液,稀释100倍后取200μl加到15ml、50℃左右的沙氏培养基中,摇匀,待凝后倒置培养48h,观察每个平皿的菌落数,计算每毫升CFU及T.asahii生长抑制率。空白对照组不加Mφ及IFN-γ,其余同上[6]。
每毫升T.asahii菌落形成单位(CFU/ml)=菌落数×稀释倍数(100)×5;T.asahii生长抑制率=(空白对照组平均CFU/ml-实验组平均CFU/ml)/空白对照组平均CFU/ml×100%
1.3.5 上清液中亚硝酸根离子(NO2-)的检测 本实验参照Ding[7]方法,-80℃冰箱取出待测上清液,恢复至室温,用含10%FBS的RPMI1640 稀释标准品1mol/L NaNO2溶液,使其浓度分别为0、1、2、5、10、20、40、60、100μmol/L,每孔依次加 GriessⅠ试剂50μl,GriessⅡ试剂50μl,室温放置10min,在全自动定量绘图酶标仪上550nm波长处测各孔吸收度值。以浓度值(C)为横坐标,吸光度值(A)为纵坐标,绘制标准曲线。结果:A=0.052+0.016C,r2=0.999。根据标准曲线, 由A值求得相应的C值,结果以μMNO2-/106MΦ表示。
1.3.6 上清液中超氧阴离子(O2-)的检测 上清液中每孔依次加入终浓度为(10、100、1000U/ml)的IFN-γ,共同孵育18h,吸弃培养液,用PBS洗涤一次。每孔依次加入超氧化物检测缓冲液200μl、WST-1溶液10μl,将产生的超氧化物还原成橙色的有色物,Catalase溶液2μl清除过氧化氢等过氧化物对WST-1显色的干扰,37℃孵育3min,再加入内毒素(lipopolysaccharide,LPS)、菌液共同在37℃、5% CO2培养箱中孵育90min后检测超氧化物阴离子O2-。
1.3.7 统计学分析 实验结果以X ± S表示,采用SPSS 16.0软件作整体F 检验提示方差齐,用单因素方差分析(ANOVA)中的Bonferroni法进行各组均数间的两两比较。
2 结果
2.1 不同浓度IFN-γ对RAW264.7细胞抗T.asahii活性的影响(表1)
各实验组与Mφ对照组比较P<0.01,结果表明:随着lFN-γ浓度的增加,T.asahii生长抑制率增加,其效应为剂量依赖性。
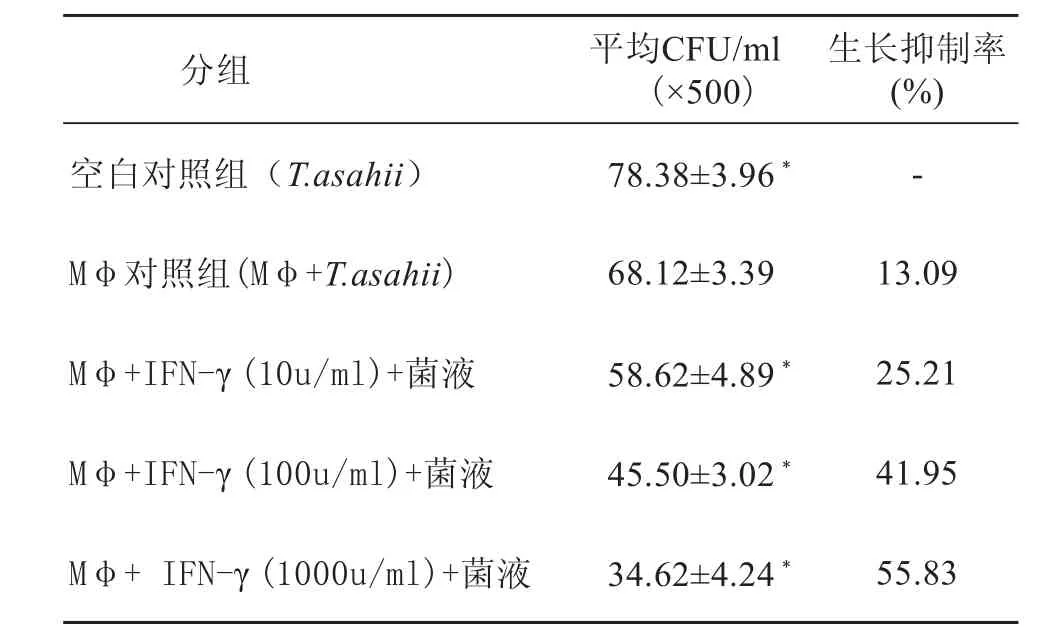
表1 Mφ在不同浓度IFN-γ中抗T.asahii的活性
2.2 上清液中亚硝酸根离子(NO2-)检测结果(表2)
lFN-γ的浓度分别为 10、100、1000U/ml时,测得上清液中NO2-的浓度,分别为89.16±3.32、108.07±3.81、126.44±3.52,与空白对照组(74.47±1.48)相比组间具有显著性差异(P<0.01)。
2.3 上清液中超氧阴离子(O2-)的检测结果(表2)
lFN-γ的浓度分别为10、100、1000U/ml时,测得上清液中的O2-分别为0.317±0.006、0. 424±0.006、0.518±0.006,与空白对照组(0. 222±0.008)相比均具有显著性差异 (P<0.01)。

表2 不同浓度IFN-γ对巨噬细胞抗T.asahii产生NO及O2-的影响
3 讨论
巨噬细胞作为重要的免疫细胞在抵抗感染中起着关键作用,它通过调理素(特异性抗体、补体、甘露聚糖结合蛋白)和非调理素(甘露聚糖受体、葡聚糖受体等)依赖的机制吞噬病原体,并产生多种效应分子杀伤病原体。若巨噬细胞不能利用这些机制将病原体吞噬、杀伤,就会使病原体逃逸,造成慢性感染[8]。
IFN-γ生物学活性广泛,在免疫应答和免疫调节中发挥着重要的作用,临床上用来抗病毒、抗肿瘤、治疗过敏性皮炎(AD)、治疗类风湿性关节炎(RA)[9]、抗肝纤维化[10]等。有研究表明宿主免疫细胞在病原体感染过程中被激活,IFN-γ通过激活Mφ,活化的巨噬细胞在NADPH(烟酰胺腺嘌呤二核苷酸磷酸)氧化酶的催化作用下将分子氧还原成O2-,在铁离子的催化下,形成羟自由基,后者与卤化物反应形成活性氧介质(reactive oxygen intermediate,ROI)进一步氧化病原体胞膜上的氨基酸,从而杀伤病原体[11]。还有研究表明,Mφ内的诱生型一氧化氮合酶(inducible NOS,iNOS)可利用精氨酸、分子氧及NADPH作为底物催化产生一氧化氮(NO),NO的存在形式不稳定,其合成后很快被氧化成NO2-,通常以亚硝酸根的形式存在于细胞外液。NO及其衍生物统称为活性氮介质(reactive nitrogen intermediates,RNI),对多种病原体具有杀伤作用[12]。总之,IFN-γ能激活巨噬细胞并促进其胞内杀菌酶活性增强,诱导产生O2-和NO等强烈杀菌物质,从而增强机体对病原体的抵抗力[13-15]。
以往的研究提示IFN-γ可增强对烟曲霉、白念珠菌等病原真菌的杀伤作用[16],Lyman等[17]的研究显示,IFN-γ预孵育可明显增强单核细胞杀伤T.asahii的活性,我们的结果证实了IFN-γ可明显增强巨噬细胞抵抗T.asahii感染的活性。本实验参照Ding[7]方法,用Griess反映测定细胞培养液中的NO2-以反映NO的浓度变化。IFN-γ在( 1~ 1000U/ml)不同浓度组产生NO 的浓度随着IFN-γ浓度的升高而升高,两者呈剂量依赖的关系。T.asahii与1000U/ml的IFN-γ共孵育后,细胞培养上清液中NO的产生明显增多,Mφ表现出高效的杀伤T.asahii的活性。本实验研究提示:随着IFN-γ浓度的升高,T.asahii的生长抑制率增加。IFN-γ增强Mφ杀伤T.asahii的作用机制与NO和O2-产生增多有关。
[1]杨蓉娅, 敖俊红, 王文岭, 等. 阿萨希丝孢酵母引起播散性毛孢子菌病国内首例报告 [J].中华皮肤科杂志, 2001,34(5):329-332.
[2]尹秀云, 陈建魁, 高峰, 等. 一例极重度骨髓型急性放射病人合并阿萨丝孢酵母菌感染的实验鉴定 [J]. 中华放射医学与防护杂志, 2007, 27(1):32-33.
[3]唐芹芳, 文怡, 梅亚宁, 等. 侵袭性阿萨希毛孢子菌和克柔假丝酵母菌双重感染1例 [J]. 临床检验杂志, 2008, 26(3):206.4.
[4]Thibeault R, Champagne M, Repentigny Lde, et al. Fatal disseminated Trichosporon asahii infection in a child with acute lymphoblastic leukemia [J]. Can J Infect Dis Med Microbiol,2008, 19(2):203-205.
[5]杨蓉娅, 李天伟, 赵建花, 等.国内首例播散性毛孢子菌病病原菌阿萨希丝孢酵母菌的DNA序列分析 [J]. 中华医学杂志,2001, 81(8):472-475.
[6]Redmond HP, Shou J, Gallagher HJ, etal. Macrophage-dependent candidacidal mechanisms in the murine system.Comparison of murine kupffer cell and peritoneal macrophage candidacidal mechanism [J]. J Immunol, 1993, 150(8Pt 1):3427-3433.
[7]Ding AH, N athan CF, Stueher DJ. Release of reactive nitrogen intermediates and react ive oxygen intermediates from mouse peritoneal macrophages [J]. J Immunol, 1988, 141(7):2407-2412.
[8]Roilides E, Katsiardani AA, Georgiadou AD, et al. Suppressive effects of interleukin 10 on human mononuclear phagocyte functi on against Candida albicans and Staphylococcus aureus [J].J Infect Dis, 1998, 178(6):1734-1742.
[9]Stevens SR, Hanifin JM, Hamilton T, et al. Long-term effectiveness and safety of recombinant human interferon gamma therapy for atopic dermatitis despite unchanged serum IgE levels [J]. Arch Dermatol, 1998, 134 (7):799-804.
[10]Baroni GS, D'Ambrosio L, Curto P, et al. Interferon gamma decreases hepatic stellate cell activation and extracellular matrix deposition in rat liver fibrosis [J]. Hepatology, 1996, 23(5):1189-99.
[11]Vazquez G J, Robledo I E, Arroyo A, et al. Acomparison of the antimicrobial resistance patterns of gram-positive cocci isolated from community -private and university-affiliated hospitals from Puerto Rico [J]. PR Health Sci J, 2003, 22(2):131-136.
[12]A ggard E. Nitricoxide: mediator, murderer and medicine [J].Lancet, 1994, 343(8907):1199-1206.
[13]Yoshimasa Yamamoto, Thomas M Klein, Mitsugu Tomioka, et al. Differential effects of Granulocyte/Macrophage Colony-Stimulating factor (GM-CSF) in enhancing macrophage resistance to legionella pneumophila vs Candida albicans [J].Cell Immunol, 1997, 176(1):75-81.
[14]H Paul Rimond, Jian Shou, Hubert J Gallagher, et al.Macrophage-dependent candidacidal mechanisms in the murine system.Compaerison of murine Kupffer cell and peritoneal macrophage candidacidal mechanisms [J]. J Immunol, 1993,150(8Pt 1):3427-3433.
[15]Marodi L, Schreiber S, Anderson DC, et al. Enhancement of macrophage candidacidal activity by interferon-gamma.Increased phagocytosis-killing,and calcium sigal mediated by a decreased number of mannose receptors [J]. J Clin Invest, 1993,91(6):2596-2601.
[16]Gaviria JM, van Burik JA, Dale DC, et al. Comparison of interferon-gamma, granulocyte colony-stimulating factor, and granulocyte-macrophage colony-stimulating factor for priming leukocyte-mediated hyphal damage of opportunistic fungal pathogens [J]. J Infect Dis, 1999, 179(4):1038-1041.
[17]Lyman CA, Garrett KF, Pizzo PA, et al. Response of human polymorphonuclear leukocytes and monocytes to Trichosporon beigelii: host defense against an emerging opportunistic pathogen [J]. J Infect Dis, 1994, 170(6):1557-1565.
猜你喜欢
科学(2020年3期)2020-11-26
科学(2020年3期)2020-11-26
现代园艺(2017年13期)2018-01-19
中学生数理化·八年级物理人教版(2017年6期)2017-11-09
渔业致富指南(2016年12期)2016-11-11
科学大众(中学)(2015年9期)2015-10-12
少儿科学周刊·少年版(2015年3期)2015-07-07
生物进化(2014年2期)2014-04-16
安徽医专学报(2014年6期)2014-03-20
现代检验医学杂志(2014年1期)2014-03-03
