
云南特有种馨香木兰种群格局的分形特征1)
2013-03-05 09:58祁荣频孟广涛邵金平李贵祥
东北林业大学学报 2013年5期
柴 勇 祁荣频 孟广涛 邵金平 李贵祥
(云南省林业科学院,昆明,650201)
分形(Fractal)理论是法国数学家曼德尔勃罗特(B. Mandelbrot)提出来的[1],用以定量研究那些不规则但其局部与整体又有某种自相似性、其维数不必为整数的几何体或形态。分形理论具有较多的特征,如用传统几何语言无法描述的不规则性、任意小的比例尺都可以包含整体的精细结构、局部到整体的不同尺度的比例变换等,但其最主要的特征是自相似性和分形维数。分形的自相似性,是指重复放大分形的局部可看到本身相似结构的再度出现,并且这种出现过程具有随机性,只有大小的区别,而没有形状的不同。这些具有自相似性结构的几何体或形态,在几何上又都可以用一个有效的空间维数——分形维数来表示。分形维数为一个连续变化的数,是用来定量描述分形所具有的自相似性的参数[2-4]。在自然界,每一个植物种都是由许多个体组成,而且都具有自己独立的特征、结构和功能。在小范围内,各种植物个体在空间上和时间上的不同配置可组合成不同的植物群落;在大范围内,许多植物群落组合在一起则构成一个整体——植被。因此,植被可视为分形的一种。目前将分形理论应用于植被格局的研究已较为广泛,从植株分枝、植冠、种群、群落,到景观各个层次的空间格局均已有不同程度的研究[5-12]。
馨香木兰(Magnolia odoratissima)是木兰科(Magnoliaceae)乔木树种,国家二级重点保护野生植物,按IUCN 地方濒危等级标准评价属“极危种(CR)”,仅分布于云南省的广南、西畴等县,为云南特有种[13]。有关馨香木兰的研究在分类学[14-17]、细胞学[18]、生态生物学[19]方面均有报道。本文在前期研究[20-22]的基础上,应用分形理论对馨香木兰种群格局的分形特征进行了研究,以进一步掌握其分布规律和生态适应性,为其保护和管理提供理论依据。
1 研究区概况
调查点位于云南省广南县黑支果乡、西畴县新马街乡及麻栗坡县大坪镇。由于地处滇东岩溶区的南端,该区域多为石灰岩山地,海拔1 300 ~1 700 m,地势沿山脊起伏变化不大,相对高差较小。气候属亚热带季风气候,年均气温15. 9 ℃,年均降水量1 294 mm,年均相对湿度为82%,年日照时间1 500 ~1 600 h,年均无霜期350 d。土壤以发育在砂岩、砂页岩上的红壤或红褐壤为主。由于水热条件较好,植被生长繁茂,地带性植被为南亚热带常绿阔叶林。馨香玉兰所在群落可分为圆果化香树(Platycarya longipes)林、青冈(Cyclobalanopsis glauca)林、西畴润楠(Machilus sichourensis)林等几种类型,各类型群落结构特点可参阅文献[20]、[21]。
2 研究方法
2.1 样地设置
在研究区内馨香木兰分布的典型地段设置6 块20 m×20 m 的样地。其中:1、2 号样地(Q1、Q2)为圆果化香树林;3、4 号样地(Q3、Q4)为西畴润楠林;5、6 号样地(Q5、Q6)为青冈林。对每块样地记录其海拔、坡向、坡度、土壤、植被状况等生境特征,并以样地一角为原点,测定样地内所有乔木个体的坐标,同时记录树种、树高、胸径等指标。各调查样地生境状况见表1。

表1 各调查样地的生境状况
2.2 数据处理
2.2.1 计盒维数的计算
以馨香木兰个体在各样地的分布坐标为基础,用不同边长的网格逐次对其进行栅格化,将每次栅格化所得的非空格子数N(ε)与对应的网格边长ε在双对数坐标下进行直线拟合,所得直线斜率的绝对值即为它们的计盒维数,计算公式为[10]:
Db=lim(lnN(ε)/lnε)。
式中:Db为计盒维数;ε 为划分尺度(即格子的边长);N(ε)为对应于划分尺度ε 的非空格子数。
本研究在确定ε 值时,将样地的边长(20 m)由2等分划分到20 等分,划分19 次,得到相对应的19 个ε 值:10.00、6.67、5.00、4.00、…、1.05、1.00 m。
2.2.2 关联维数的计算
关联维数的计算公式为[8]:Dc= lim(lgC(ε)/lgε)。
研究中仍以馨香木兰个体在各样地的分布坐标为基础,先将样地内所有馨香木兰个体两两间的欧氏距离计算出来,然后给定一个距离值,查找小于该距离的欧氏距离个数(Ni);变换距离值,可以得到一系列Ni;通过每一距离个数与距离总个数(N)的比值得到C(ε)。将C(ε)与对应的距离值在双对数下进行(分段)直线拟合,所得拟合直线斜率的绝对值即为其关联维数(Dc)。在本研究中,根据样地内所有个体两两间欧氏距离的计算结果,确定给定距离ε 值的范围为0 ~22 m,变换步长为0.5 m。
3 结果与分析
3.1 馨香木兰种群格局的计盒维数
馨香木兰种群格局的计盒维数计算结果见图1。图1中,6 块样地均出现两个线性区域,拟合出两条直线,也就是说出现了拐点,其拐点尺度(计盒维数发生激烈变化时的尺度)分别为2.5、2.5、4.0、3.3、3.3、4.0 m。在拐点前尺度范围内,计盒维数介于0.028 4 ~0.202 6 之间,它们的直线拟合效果欠佳,相关系数均在0.5 以下,且样地Q1、Q4、Q6 尚未达到显著水平(P >0.05)。在拐点后尺度范围内,计盒维数介于0. 535 1 ~0. 986 8 之间,其中样地Q1、Q2、Q3、Q5 的直线拟合效果较好,相关系数均在0.9 以上并达到显著水平(0.01 <P <0.05)或极显著水平(P <0.01)。样地Q4、Q6 直线拟合的效果则较差,未达到显著水平。
3.2 馨香木兰种群格局的关联维数
馨香木兰种群格局的关联维数结算结果见图2。图2中可看出,各样地馨香木兰种群格局关联维数直线拟合效果均达到了极显著水平(P <0.01)。其中:样地Q2、Q5、Q6 仅出现一个直线区域,关联维数分别为0.564 9、1.332 2、1.654 4。样地Q1、Q3、Q4 则分别出现两个直线区域,意即也出现了拐点,其拐点尺度分别为6、3、5 m,它们的关联维数在拐点前分别为0.798 0、0.429 7、0.169 6,在拐点后分别为1.307 5、1.161 8、1.511 0。
4 结论与讨论
计盒维数可以揭示种群占据和利用生态空间的能力[10,23]。在较小的尺度范围内(本研究中为0 ~2.5 m),计盒维数会遵循一定的规律(对应的直线斜率不变),但在较大的尺度范围内(本研究中为2.5 ~10.0 m)计盒维数可能会有不同(对应的直线斜率发生变化),意即种群占据和利用生态空间的能力在不同尺度范围内会有差异。本研究中,6 块样地计盒维数的拟合直线斜率均不同,反映了馨香木兰种群占据生态空间的能力在拐点尺度前后的剧烈变化。有研究认为[24-25],小于拐点尺度的计盒维数反映的是位点(植株个体)对生态空间的占据,而大于拐点尺度的计盒维数反映的是种群完整个体对生态空间的占据。本研究中,馨香木兰种群格局拐点尺度前的计盒维数均远低于种群占据全部生态空间时(计盒维数为2)的水平,在一定程度上表明馨香木兰在个体水平上对生态空间的占据能力较差。而拐点尺度前较差的拟合效果则意味着在该拐点前的尺度范围内仍可能有其它拐点存在,反映了馨香木兰个体对生态空间的占据能力在小范围内仍存在尺度上的差异。拐点尺度后的计盒维数稍有增加,但仍全部低于1 而远离种群占据全部生态空间时的水平。这表明馨香木兰在种群水平上对生态空间的占据能力也较差。同样,样地Q4 和Q6 的拟合效果较差,说明这两样地馨香木兰在种群水平上对生态空间的占据能力也存在尺度上的差异。
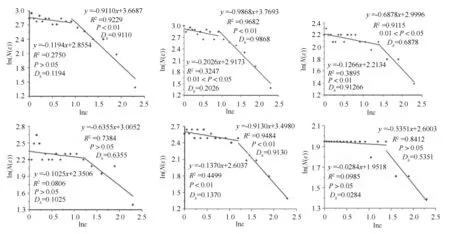
图1 不同样地馨香木兰种群格局的计盒维数

图2 不同样地馨香木兰种群格局的关联维数
理论上,种群占据全部生态空间时计盒维数应为2,余下的维数为生态间隙维。计盒维数反映的是种群现实占据生态空间的能力,而生态间隙维则反映的是种群潜在占据生态空间的能力[25]。据此分析,馨香木兰种群格局的生态间隙维在拐点前、后分别介于1.797 4 ~1.971 6 和1.013 2 ~1.464 9 之间,意味其潜在可占据的生态空间所占比例分别高达89.87% ~98.58%和50.26% ~73.25%,这在理论上似乎表明馨香木兰种群在群落中尚有较大的生存发展空间。然而,实事表明,目前在自然界很少有馨香木兰的纯林,其植株个体多呈散生状态分布在以圆果化香树、青冈、西畴润楠等为主要优势种的常绿阔叶林中,且多数都处于林木下层,它们在种群数量、个体形态、空间占据等方面与其它树种相比均无优势,因而其潜在的生存空间实际上极其有限。
计盒维数的拐点尺度是种群格局斑块大小的反映,能够在一定程度上看出种群空间占据能力在尺度上的差异。在计盒维数的计算过程中,拐点尺度大小及拐点数量的确定往往带有一定的主观性。在直线拟合效果不佳的情况下,理论上一般可以通过增加拐点来增强拟合效果,拐点及拟合直线的增加更多地反映了种群空间占据能力在尺度上的剧烈变化过程。本研究在计算计盒维数时只用了一个拐点,部分样地的拟合效果未到达显著水平,这说明馨香木兰种群对空间的占据能力在研究尺度范围内可能存在更多的拐点,需要进一步结合植株形态、冠幅、种内种间关系等加以分析。
种群格局的关联维数可揭示种群个体空间相关的尺度变化规律,反映种群个体的空间相关程度[8]。在关联维数的直线拟合过程中可能出现1 ~2 个直线区域:如果只存在1 个直线区域,则说明种群个体空间分布规律相同,呈非聚集分布;若存在2个直线区域,表明个体空间分布规律不一致,表现为聚集分布。关联维数的大小则可以反映种群个体空间相关程度和个体间的竞争强度:关联维数大(接近于2),表明种群个体的空间相关程度高,种群整体对空间的占据程度也较高,个体竞争强烈;反之,种群个体的空间相关程度低,整体对空间的占据程度较弱[26]。本研究中,样地Q2、Q5、Q6 仅出现一个直性区域,表明在这3 个样地中馨香木兰的个体分布在研究尺度范围内具有一定的规律性(每个个体周围的同种个体数随距离增加而有规律地增加),种群表现为非聚集的分布格局。其中:样地Q5、Q6的关联维数相对较高,表明样地中馨香木兰个体的空间相关程度较高,种群对空间的占据程度较高,个体间竞争剧烈;样地Q2 的关联维数处于较低的水平,说明样地中馨香木兰个体间的空间相关程度较低,个体间竞争较弱;样地Q1、Q3、Q4 则分别出现了两个直线区域,表明在这3 个样地中馨香木兰的个体分布在不同尺度上存在较大差异,种群格局表现出聚集分布的特点。种群个体的空间关联性在拐点尺度前后发生激烈变化,其中在拐点后的较大尺度范围内具有更强的空间关联性,相互间竞争更剧烈。总的来说,馨香木兰种群格局的关联维数都处于较低的水平,说明其种群个体间空间关联程度较低,种群整体对空间的占据程度也较低,相互间竞争较弱。这一分析结果在一定程度上反映出馨香木兰渐临濒危的生存状态。
[1] Mandelbrot B B. The fractal geometry of nature[M]. New York:W H Freeman and Co.,1982.
[2] 赵茂程,高素萍,潘一凡,等.分形理论及其在林业中的应用与研究进展[J].世界林业研究,2002,15(2):28 -34.
[3] 高峻,张劲松,孟平.分形理论及其在林业科学中的应用[J].世界林业研究,2004,17(6):11 -16.
[4] 王倩,汪权方,梅新.分形理论在遥感图像分类处理中的应用评析[J].安徽农业科学,2009,37(7):3325 -3326.
[5] 邓红兵,周永斌,王庆礼.三峡库区次生柏木林中物种分布格局的分形特征[J].应用生态学报,1999,10(5):518 -520.
[6] 梁士楚.云贵鹅耳枥种群分布格局的分形特征[J]. 武汉植物学研究,2001,19(4):263 -268.
[7] 倪红伟,马克明,赵伏臣.小叶樟种群分布格局的分形特征Ⅰ计盒维数[J].植物研究,2000,20(2):229 -234.
[8] 马克明,祖元刚,倪红伟. 兴安落叶松种群格局的分形特征——关联维数[J].生态学报,1999,19(3):353 -358.
[9] 马克明,祖元刚. 植被格局的分形特征[J]. 植物生态学报,2000,24(1):111 -117.
[10] 张文辉,祖元刚,马克明.裂叶沙参与泡沙参种群分布格局分形特征的分析[J].植物生态学报,1999,23(1):31 -39.
[11] 梁士楚,王伯荪.红树植物木榄种群植冠层结构的分形特征[J].海洋通报,2002,21(5):26 -31.
[12] 马克明,祖元刚. 兴安落叶松分枝格局的分形特征[J]. 植物研究,2000,20(2):235 -241.
[13] 李玉媛.云南国家重点保护野生植物[M].昆明:云南科学技术出版社,2005:18 -26,211 -212.
[14] 刘玉壶. 中国木兰[M]. 北京:科学技术出版社,2004:82 -83.
[15] 司马永康,余鸿.木兰属一新命名[J]. 云南林业科技,2003,32(1):39.
[16] 李捷.中国木兰科植物修订[J].云南植物研究,1997,19(2):131 -138.
[17] 夏念和,邓云飞.木兰科植物订正[J].热带亚热带植物学报,2002,10(2):128 -132.
[18] 王亚玲,张寿洲,李勇,等.木兰科13 个分类群和12 个杂交组合的染色体数目[J].植物分类学报,2005,43(6):545 -551.
[19] 张茂钦.云南珍稀濒危树种生态生物学研究[M]. 昆明:云南大学出版社,1998:253 -264.
[20] 柴勇,孟广涛,武力,等. 馨香玉兰所在群落主要树种的种间联结性[J].西北林学院学报,2009,24(5):31 -35.
[21] 柴勇,孟广涛,武力,等. 馨香木兰所处植物群落的物种多样性及其种群结构特征研究[J].西部林业科学,2008,37(3):46 -50.
[22] 祁荣频,柴勇,武力,等. 馨香木兰集中分布地生境的土壤理化性状研究[J].西部林业科学,2010,39(4):95 -98.
[23] 郭华,王孝安,肖娅萍.秦岭太白红杉种群空间分布格局动态及分形特征研究[J].应用生态学报,2005,16(2):227 -232.
[24] 贺强,崔保山,胡乔木,等. 水深环境梯度下柽柳种群分布格局的分形分析[J].水土保持通报,2008,28(5):70 -73.
[25] 马克明,祖元刚.兴安落叶松种群格局的分形特征:计盒维数[J].植物研究,2000,20(1):104 -111.
[26] 吴承祯,洪滔,宋萍,等. 长苞铁杉种群空间格局的分析特征[J].福建林学院学报,2008,28(1):1 -5.
猜你喜欢
教学月刊(中学版)(2022年12期)2022-12-27
数学物理学报(2022年4期)2022-08-22
工会博览(2022年17期)2022-07-15
数学物理学报(2020年3期)2020-07-27
青年生活(2020年6期)2020-03-28
幽默大师(2019年5期)2019-05-14
东坡赤壁诗词(2018年2期)2018-05-10
鹿鸣(2018年1期)2018-01-30
歌剧(2017年4期)2017-05-17
营销界(2015年25期)2015-08-21
