
长白落叶松家系碳含量遗传变异与高固碳家系选择1)
2013-03-05 09:58贾庆彬张含国王美玲朱航勇
东北林业大学学报 2013年5期
贾庆彬 张含国 王美玲 朱航勇 刘 灵
(林木遗传育种国家重点实验室(东北林业大学),哈尔滨,150040)
森林生态系统是陆地生态系统最大的碳库,在全球碳循环中起到重要作用[1]。近年来,随着大气中二氧化碳含量的增加,全球气候变化所造成的危害不断加剧,人们对森林生态系统在控制碳平衡中的重要作用有了更深的认识[2]。目前,国内外关于森林碳汇的研究已取得一定成果。马钦彦等[3]对华北地区主要森林类型的8 个乔木建群种含碳率进行研究时发现,华北落叶松树干部分平均含碳率为49%左右,针叶树种各器官的含碳率普遍比阔叶树种高出1.6% ~3.4%,相应的针叶林的碳储量也高于阔叶林。林分生物量对于森林碳储量的贡献极显著。Poudel 等[4]在对瑞典中北部地区森林生物量与碳平衡的研究中发现,树干部分生物量达到林木总生物量的50%以上,且该比例随树龄的增长而增加。巨文珍等[5]在长白落叶松林龄序列上的生物量及碳储量分配规律的研究中发现,除去较小的幼龄林分外,其他林分的林木,其树干生物量都是地上生物量的主体(所占比例平均为56.12%),而地上部分生物量又对总生物量起到决定作用。碳储量的分配与生物量相同,地上部分为林分碳储量的主要部分,这与罗云建等[6]对华北落叶松人工林生物量及其分配模式的研究、尤文忠等[7]对辽东山区落叶松人工林和蒙古栎天然次生林固碳功能的研究中所得结果相同。本文以24年生长白落叶松(Larix olgensis)半同胞家系试验林为研究对象,测定植株碳含量,并结合生长、材性性状进行综合分析,选择出具备高固碳能力的家系,为长白落叶松的杂交育种、营造高固碳人工林等提供依据。
1 材料与方法
1.1 试验林概况
试验林设于黑龙江林口县青山林场,种子来源于青山长白落叶松种子园。1987年春播种育苗,1988年春用1年生苗造林。试验采用完全随机区组设计,处理数35 个,重复5 次,小区8 株,共33 个家系。设两个对照,其中生产对照一个,种子园混杂种子对照一个。试验地面积0.67 hm2,株行距离2.0 m×1.5 m。
1.2 取样与试验
对试验林35 个处理4 次重复(Ⅰ、Ⅲ、Ⅳ、Ⅴ次重复)进行木材取样,每个处理每重复随机选取2株树作为样本,在胸高处同一方向用直径5 mm 的生长锥取得由树皮至髓心的完整无疵木芯,共取样品280 个。
木材密度利用饱和含水量法进行测定,综纤维素与木质素含量分别参照国家标准GB/T 2667.10—95 和GB/T 2667.8—94 进行测定。
含碳率利用德国耶拿分析仪器股份公司的碳元素分析仪multi EA 4000 进行分析。温度设定1 000℃,进样量50 mg。
干材生物量及碳储量计算方法:

式中:V 为材积,立木材积按平均试验形数法计算;f∍为形数,长白落叶松平均试验形数为0.41;h 为树高;g1.3为胸高处横断面积;ρ 为木材密度;C 为碳储量;B 为生物量;Cc为含碳率。
计算家系遗传力与遗传增益。
遗传力:h2=1 -1/F;
式中:h 为家系遗传力的平方根;i 为选择强度;σA为选择性状的标准差;为家系数值的平均数。
1.3 数据统计分析
数据分析采用PASW Statistics18 统计软件进行处理,主要包括方差分析、相关分析、Duncan 法检验等。
2 结果与分析
2.1 长白落叶松家系生长性状遗传变异
对长白落叶松33 个家系生长性状进行遗传变异分析,列出各性状变异系数较小的5 个家系和变异系数较大的5 个家系,见表1。结果显示:胸径性状变异相对较小,树高、材积存在较大变异,三者变异系数平均值分别为9. 95%、15. 08%、24. 87%。材积变异系数较大的前5 个家系分别为73 -12、73-14、73 -16、73 -21、73 -49,5 个家系平均变异系数为43.63%,高出家系总平均值75.43%,高出对照CK 与种长混75.50%和92.63%。其中:变异系数最大的家系73 -12 分别高于总平均85.32%、对照组CK 85.40%、种长混103.49%。

表1 长白落叶松家系生长性状均值与变异系数
2.2 长白落叶松家系材性性状遗传变异
对长白落叶松33 个家系材性性状进行遗传变异分析,列出了各性状变异系数较小的5 个家系和变异系数较大的5 个家系,见表2。
他带领五粮液进行二次创业;他进一步改革集团治理模式推进五粮液发展新战略,大力实施“引进来走出去”战略,强化国际国内顶级酒业的战略性并购;他依托财务公司和资本优势,强化资本运作,创新金融服务,特别是在产业链金融方面,加大对上游供应商和下游经销商的融资服务,为五粮液销售额再上新台阶不懈努力。

表2 长白落叶松家系材性性状均值与变异系数
结果显示:木材密度、综纤维素质量分数、木质素质量分数变异均较小,三者变异系数平均值分别为8.65%、5.70%、5.40%。木材密度变异系数较大的前5 个家系分别为73 -12、73 -18、73 -28、73-3、73 -49,5 个家系平均变异系数为12.37%,高出家系总平均值43. 01%,高出对照种长混33.30%,低于CK,为16.98%。其中,变异系数最大的家系73 - 28 高于总平均79. 97%,高出对照组CK7.59%、种长混67.72%。纤维素质量分数变异系数较大的前5 个家系分别为73 -11、73 -29、73 -34、73 -45、73 -49,5 个家系平均变异系数为11.45%,高出家系总平均值100.86%。木质素质量分数变异系数较大的前5 个家系分别为73 -28、73 -35、73 -46、73 -5、73 -7,5 个家系平均变异系数为11.10%,高出家系总平均值106.31%。
2.3 长白落叶松家系含碳率、树干生物量、碳储量遗传变异
对长白落叶松33 个家系含碳率、树干生物量、碳储量进行遗传变异分析,得出各性状变异系数较小的5 个家系和变异系数较大的5 个家系,见表3。

表3 长白落叶松家系含碳率、树干生物量、碳储量均值与变异系数
结果显示:树干生物量与碳储量存在较大变异,含碳率变异较小,三者变异系数分别为27. 84%、27.68%、2.58%。含碳率变异系数较大的前5 个家系为73 -12、73 -32、73 -42、73 -50、73 -7,5 个家系平均值为4.65%,高出总平均值80.62%,高出对照CK40.66%、种长混110.01%。在碳储量方面,变异系数较大的前5 个家系为73 -49、73 -14、73 -12、73-16、73-47,5 个家系平均值为50.02%,分别高出总平均值80.74%、高出对照CK56.20%、种长混69.55%。其中,变异系数最大的家系73 -49 分别高于总平均94.23%、对照组CK67.85%、种长混82.21%,高出变异系数最小的73-48 号家系7 倍。
2.4 长白落叶松各性状家系间差异性与高固碳家系选择
对长白落叶松各性状测定值进行方差分析,结果见表4。

表4 长白落叶松家系各性状方差结果
家系间生长性状、主干生物量、碳储量存在显著差异。对这3 个性状分别利用Duncan 法进行多重比较,发现73 -46、73 -49、73 -11、73 -14 这4 个家系与其它家系间均存在明显差异。在碳储量方面,4 个家系平均值为29.06 kg,分别高出家系总平均值57.05%、对照CK130.46%、种长混58.33%,高出碳储量最小的73 -45 家系190.86%。其中,碳储量最大的73-14 家系分别高出家系总平均值70.88%、对照CK150.76%、种长混72.28%,高出碳储量最小的73 -45 家系216.48%。选择以上4 个家系作为优良家系,则碳储量家系遗传力为0.634,选择强度i=2.023,遗传增益为0.568。材积与生物量的家系遗传力分别为0.671、0.639。
2.5 相关关系
林木某一性状的优良表现有时是多个性状相互作用的结果。通过相关分析了解各性状间的关系,探寻性状间的内部联系,通过对某一方面的改良使其它性状得到促进或抑制。本试验采用Pearson 法对33 个家系不同性状的测定值进行相关分析的结果显示(表5):含碳率与材积、木质素质量分数、碳储量呈正相关,与胸径、木材密度、综纤维素质量分数、生物量呈负相关。但无论正负,相关性均未达到显著水平,相关关系表现微弱。碳储量方面,除仅与木质素质量分数呈现微弱负相关之外,与其它各性状均呈正相关关系。其中,生长性状、生物量与碳储量呈极显著正相关。
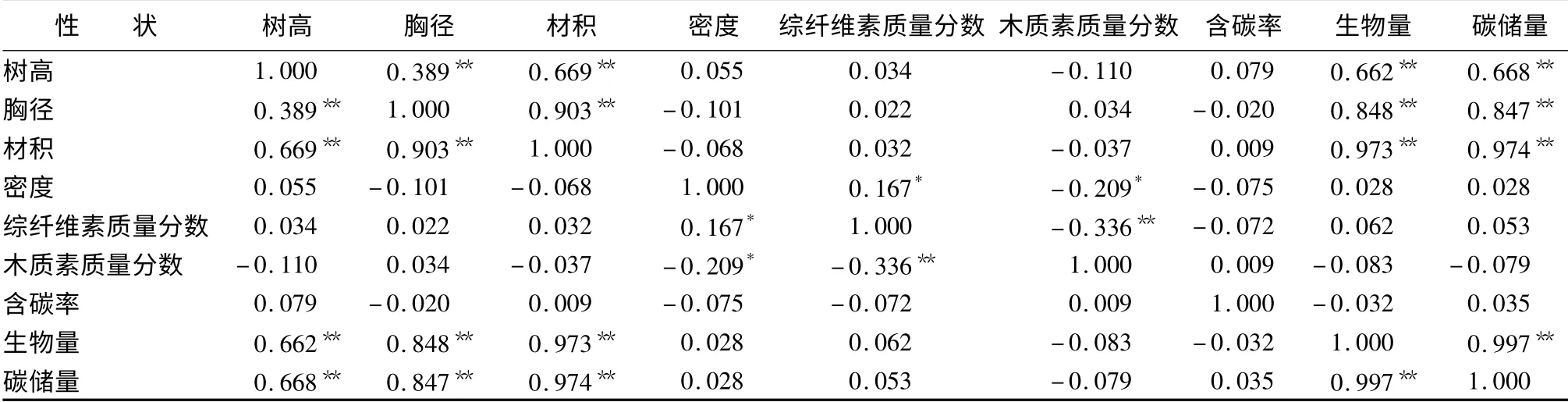
表5 长白落叶松家系各性状相关性
3 结论与讨论
长白落叶松33 个家系各性状中,材性性状与含碳率变异相对较小,变异系数分别为:木材密度8.65%、综纤维素质量分数5.70%、木质素质量分数5.40%、含碳率2.58%。材积、生物量、碳储量存在较丰富变异,三者变异系数分别为24.87%、27.84%、27.68%,家系遗传力分别为0.671、0.639、0.634,受中等强度的遗传控制,这为今后长白落叶松家系固碳能力的改良提供可能。
相关分析结果显示:木材密度与材积呈负相关关系,相关系数为-0.068,说明随着材积的增大,木材密度可能会有所下降。这与以往孙晓梅等[11]在日本落叶松纸浆材优良家系多性状联合选择、邓继峰[12]在17年生杂种落叶松遗传变异及优良家系选择等研究中所得结果相近。尽管木材密度与材积呈现负相关关系,但相关不显著,且表现微弱,表明并不是所有生长较快的植株的木材密度均会下降,因此,通过合理选择,可以筛选出木材密度较高且生长性状优良的家系。这一现象的存在很可能是由于木材密度与生长性状二者独立遗传,受不同的遗传机制控制,值得今后进行更深入地研究。
含碳率与材积、木质素质量分数、碳储量呈正相关,与胸径、木材密度、综纤维素质量分数、生物量呈负相关。而无论正负,相关都未能达到显著水平。木材中的碳主要以纤维素和木质素形式存在,本试验结果显示,含碳率与木质素呈正相关关系,这对今后通过其它途径对木质素质量分数进行遗传改良,进而提高林木含碳率具有一定的参考作用。以往对于森林固碳能力的研究多集中于天然林,为了使试验得到简化,通常采取控制含碳率为某一数值的方法估计某个区域内森林的总碳储量。但由于树种不同,含碳率范围一般在47% ~59%之间,目前被广泛接受且最常使用的数值为50%[13]。这一方法为从宏观上估算森林碳储量提供了方便,但是,本试验所得结果显示,样地内林木总体平均含碳率为45.65%,低于以往普遍的含碳率(50%)。分析原因,本试验所选样地为人工林,由于人为经营与管理,人工林内植株生长较快,生物量普遍高于同龄的天然林林木,而且生物量与含碳率存在微弱的负相关关系,这很可能是产生含碳率降低的原因之一。另外,本文是以林木的树干部分为研究对象,而以往的研究显示,林木不同器官含碳率存在差异,且针叶树种的枝、叶、皮含碳率普遍高于树干[14],这也是本文所得结果低于平均含碳率的原因。很多研究结论已表明,生物量对森林碳储量的影响起到决定作用,尽管含碳率有所下降,但综合生物量因素,单位面积人工林分的碳储量依然具有优势。黄从德[15]在四川森林生态系统碳储量及其空间分异特征的研究中也发现,人工林碳储量年均增长率明显大于天然林,随着人工林的进一步发展,人工林的碳汇功能将更加突出。
在碳储量方面,除仅与木质素质量分数呈现微弱负相关之外,与其它各性状均呈正相关关系。其中,生长性状、生物量与碳储量呈极显著正相关。此结果表明,生长性状与生物量的优秀表现对林木碳储量的增强具有积极的促进作用,这为今后通过对林木生长性状进行改良进而提高森林碳储量提供了可能,可作为营建高固碳人工林的生产实践参考。
综合研究结果,通过方差分析与Duncan 法多重比较,最终选择出73 -46、73 -49、73 -11、73 -14这4 个家系为高固碳能力家系。4 个家系碳储量平均值为29.06 kg,分别高出家系总平均值57.05%、对照CK130.46%、种长混58.33%,高出碳储量最小的73 -45 家系190.86%。其中,碳储量最大的73 -14 家系分别高出对照CK150.76%、种长混72.28%。以现有试验林为标准,按相同密度定植,若选择73 -14 家系,则每公顷固碳为65 728.32 kg,比目前试验林每公顷固碳量高70.88%,比利用碳储量最小的73 -45 家系造林每公顷固碳量高出2.16倍。由试验结果可见,通过合理选择,利用高固碳能力的家系造林,森林单位面积的碳储量将得到明显提高,对增加森林碳汇,发展绿色经济具有重要意义。
[1] Kimble J M,Birdsey R,Heath L S. The potential of U.S. forest soils to sequester carbon and mitigate the greenhouse effect[M].Boca Raton,FL:CRC Press/Lewis Publishers,2002:429.
[2] Vogel J G,Bond-lamberty B P,Schuur E A G,et al. Carbon allocation in boreal black spruce forests across regions varying in soil temperature and precipitation[J]. Global Change Biology,2008,14(7):1503 -1516.
[3] 马钦彦,陈遐林,王娟,等.华北主要森林类型建群种的含碳率分析[J]. 北京林业大学学报,2002,28(5/6):96 -100.
[4] Poudel B C,Sathre R,Bergh J,et al . Potential effects of intensive forestry on biomass production and total carbon balance in north-central Sweden[J]. Environmental Science & Policy,2012,15(1):106 -124.
[5] 巨文珍,王新杰,孙玉军,等.长白落叶松林龄序列上的生物量及碳储量分配规律[J].生态学报,2011,31(4):1139 -1148.
[6] 罗云建,张小全,王效科,等.华北落叶松人工林生物量及其分配模式[J].北京林业大学学报,2009,31(1):13 -18.
[7] 尤文忠,魏文俊,邢兆凯,等.辽东山区落叶松人工林和蒙古栎天然次生林的固碳功能[J]. 东北林业大学学报,2011,39(10):21 -24.
[8] 孟宪宇.测树学[M].北京:中国林业出版社,2004:56 -67.
[9] 张超,张含国,朱航勇,等.日本落叶松碳储量家系遗传变异及优良家系选择研究[J]. 中南林业科技大学学报,2011,31(10):53 -62.
[10] 李自敬,李雪峰,张含国,等. 长白落叶松优良家系选择的研究[J].林业科技,2008,33(4):1 -4.
[11] 孙晓梅,张守攻,李时元,等.日本落叶松纸浆材优良家系多性状联合选择[J].林业科学,2005,41(4):48 -54.
[12] 邓继峰,张含国,张磊,等.17年生杂种落叶松遗传变异及优良家系选择[J].东北林业大学学报,2010,38(1):8 -11.
[13] Bert D,Danjon F. Carbon concentration variations in the roots,stem and crown of mature Pinus pinaster (Ait.)[J]. Forest Ecology and Management,2006,222(1/3):279 -295.
[14] 田勇燕,秦飞,言华,等.我国常见木本植物的含碳率[J]. 安徽农业科学,2011,39(26):16166 -16169.
[15] 黄从德.四川森林生态系统碳储量及其空间分异特征[D].雅安:四川农业大学,2008.
猜你喜欢
现代交际(2022年10期)2022-11-01
山东林业科技(2019年2期)2019-06-03
森林工程(2018年4期)2018-08-04
黄河之声(2017年6期)2017-07-01
辽宁林业科技(2017年4期)2017-06-22
黄河之声(2016年24期)2016-02-03
小主人报(2015年5期)2015-02-28
中南林业科技大学学报(2014年6期)2014-12-28
安徽工业大学学报(自然科学版)(2014年4期)2014-07-11
中国火炬(2012年9期)2012-07-25
