
大豆萌发对大豆酸奶品质的影响
2013-05-02 14:46刘爱洁冯立科
食品与机械 2013年2期
刘爱洁 冯立科 李 理
LIU Ai-jie1 FENG Li-ke2 LI Li1
(1.华南理工大学轻工与食品学院,广东 广州 510640;2.广东燕塘乳业股份有限公司,广东 广州 510507)
(1.College of Light Industrial and Food Science,South China University of Technology,Guangzhou,Guangdong 510640,China;2.Guangdong Yantang Dairy Corporation,Guangzhou,Guangdong 510507,China)
大豆酸奶作为一种新兴的谷物酸奶,能充分利用中国大豆蛋白资源,同时可以优势互补的降低牛奶酸奶产品的脂肪、胆固醇含量,对于缓解中国乳源紧张局面、增强大豆蛋白质的深加工利用以及提高国民的身体健康具有重要的意义。但因大豆球蛋白的三、四级结构致密、硬度较大,使得大豆蛋白质消化率和生物效价不高。用豆浆酶解液制得的酸豆奶不仅营养价值大大提高、发酵时间缩短、乳酸菌的产酸能力提高,同时其感官品质也得以改善[1]。目前豆浆酶解多采用Alcalase蛋白酶、木瓜蛋白酶、菠萝蛋白酶、风味酶等工业用酶制剂,这些蛋白酶活性强,水解不易控制,易过度水解使豆浆变黄、产生咸味、碱味、苦味等[2]。大豆萌发不但可以增加活性异黄酮、维生素和膳食纤维等营养物质含量[3],且种子内部产生丰富的内源蛋白酶,能对大豆蛋白亚基进行一定程度的降解。本课题组的前期研究[4]表明大豆在浸泡12 h以后就显示出较高的中性和酸性蛋白酶活性,其中中性蛋白酶活力为29.20 μg/(d·mg),酸性蛋白酶活力为 8.57 μg/(d·mg)。大豆种子萌发过程中,利用大豆种子的内源酶分解大豆本身的蛋白质、淀粉和脂肪等大分子营养物质,可以改善大豆酸奶的综合品质[4,5]。本试验以不同萌发程度的大豆为原料,从内源蛋白酶的降解作用出发,分析蛋白降解对豆浆和大豆发酵乳的品质的影响。
1 材料与方法
1.1 材料与仪器
大豆:当季成熟的东北大豆;
脱脂乳粉:蛋白含量约32.7%,新西兰Fonterra有限公司;
其他试剂:均为国产分析纯。
瑞士乳杆菌(Lactobacillus helveticus LH-B02,LH-B02):丹麦Chr.Hansen公司样品;
嗜热链球菌(Streptococcus thermophilus IFFI 6038,S.T 6038)、保加利亚乳杆菌(Lactobacillus bulgaricus AS1.1482,L.B 1482):广东省微生物研究所菌种保藏中心。
生化培养箱:PXY-190S-A型,广东韶关科力仪器有限公司;
搅拌机:BE351B型,佛山顺德美的集团有限公司;
三恒电泳仪:ECP3000型,北京六一仪器厂;
双板夹芯式垂直槽:DYCZ30型,广州市丛源仪器有限公司;
手提式高压灭菌锅:YXQ-SG46-280S型,合肥华泰医疗设备有限公司;
电热恒温水浴锅:HSG-IB-2型,常州奥华仪器有限公司;
电子天平:JJ500型,浙江余姚铭称重校验设备有限公司;
分光光度计:S22PC型,上海凌光技术有限公司;
流变仪:RS-600型,德国HAAKE公司;
pH计:S20型,瑞士梅特勒—托利多仪器有限公司;
超净工作台:SZX型,吴江净化设备总厂;
激光共聚焦显微镜:TCSSP5型,德国莱卡仪器公司。
1.2 试验方法
1.2.1 萌发大豆酸奶的制备
(1)大豆酸奶制备工艺流程[6]:
大豆→挑选→消毒杀菌→浸泡→萌发→去皮→85℃热水打浆→过滤豆浆→加糖、复原乳→灭菌→42℃发酵4 h→4℃冷藏24 h
(2)大豆籽粒萌发[6]:挑选均一、饱满、无虫蛀、表皮无破损霉变的完整大豆,冲洗去杂后用75%酒精灭菌1min,无菌水清洗6次,室温无菌水中浸泡14 h,在培养盆中铺布均匀后以4层纱布遮盖,分别萌发至胚芽长度为0,1,3,5 cm。
(3)发酵剂和发酵底物的制备:配置12%的牛乳,经115℃灭菌10min,按5%接种量(V/V)分别接入瑞士乳杆菌LH-B02、保加利亚乳杆菌L.B 1482和嗜热链球菌S.T 6038,37℃静止培养8~12 h。发酵剂经数次活化后,参照文献[7]的方法测定菌种活力,活力达到0.6时即可使用。
将不同芽长的大豆,去皮,用85℃热水打浆(豆水比为1∶8,干豆),180目筛过滤后制得豆浆。配置12%复原乳,豆浆和复原乳的比例为7∶3,添加8%的蔗糖,100℃灭菌15min,冷却至室温。
(4)接种:按5%的总接种量,在发酵底物中分别添加2%的瑞士干酪乳杆菌,1.5%的保加利亚乳杆菌和嗜热链球菌,混匀分装后42℃静止发酵4 h,4℃后熟24 h。
1.2.2 萌发大豆豆浆的蛋白亚基及抗营养因子变化
(1)萌发大豆豆浆中蛋白亚基组分含量变化:以不同芽长的大豆所制豆浆为待测样品,于沸水中加热5 min后,10 000 r/min离心10 min。参照Laemm li的电泳方法[8],稍加改动。上样量为15μL,分离胶、浓缩胶浓度分别为14%和4%,浓缩胶、分离胶电流分别为40,80mA。电泳完毕后以考马斯亮蓝R250将凝胶染色40min,脱色至透明后拍照。
(2)萌发大豆豆浆中胰蛋白酶抑制因子含量变化:将不同芽长的大豆所制豆浆取样冻干,并在100℃下加热不同时间,参照Kakade的方法[9]测定豆浆中胰蛋白酶抑制因子的活性,并依据GB/T 21498——2008(ISO 14902:2001,IDT)做稍许改动。
1.2.3 萌发大豆酸奶各项指标变化
(1)萌发大豆酸奶pH、酸度及持水力的测定:用pH计测定样品的pH值。按GB/T 5009.46——2010《乳与乳制品卫生标准的分析方法》滴定酸度,以涅尔度°T为酸度指标。
取30 g接种后的大豆酸奶,在离心管(32 mm×115 mm)中42℃发酵4 h,4℃冷藏24 h后将样品在20℃条件下480×g离心10min,去除乳清后称重。

式中:
Q——持水力,%;
W1——离心前样品的质量,g;
W2——离心后样品的质量,g。
(2)大豆酸奶流变特性的测定:将大豆酸奶顺时针、逆时针各搅拌10次之后,用哈克流变仪测定酸奶流变性质。选用40mm直径的平板探头,温度控制在(25.0±0.5)℃,分析模式:频率扫描,应变为0.5%,频率从0.1~10 Hz;剪切扫描,速率从0逐渐增加至500/s,然后再从500/s减小至0,每个步骤时间为180 s。每个测定重复2次[10]。
(3)萌发大豆酸奶的微观结构变化:大豆酸奶的微观结构应用TCSSP5倒置显微镜(物镜20×HC PL APO/0.70NA)观察[11]。接种后的牛奶豆浆混合乳先用0.1%的Nile Blue再用0.01%的Nile Red染色。将接种染色后的混合乳样品置于带凹槽的载玻片中心,小心盖上盖玻片,避免产生气泡。在盖玻片边缘涂覆硅油以防止水分蒸发。制片完成后用锡纸包裹与酸奶样品一起42℃发酵4 h。后熟24 h后将载玻片倒置在载物台上,用100倍物镜观察。激发波长为488,633 nm,接收波长在稍大于相应激发波长的范围。为防止荧光淬灭,应在尽短时间内采集图像。
1.2.4 数据处理与分析 采用SPSS 17.0统计分析软件进行一维方差分析,置信区间为95%。
2 结果与讨论
2.1 萌发大豆内源酶对豆浆的影响
2.1.1 内源酶对大豆蛋白的降解作用 大豆球蛋白(11S)和β-伴球蛋白(7S)是大豆中的主要储存蛋白,占大豆总蛋白含量的70%以上。大豆在萌发过程中内源酶对蛋白亚基的降解作用通过SDS-PAGE表现为图1。豆浆中的脂肪氧化酶LOX的条带在芽长3 cm的泳道开始变浅。LOX的降解有助于减缓豆腥味物质的形成,帮助减少发酵豆乳的豆腥味。7S球蛋白的 α’,α 亚基及 11S 酸性亚基(A1、A2,A3,A4)在大豆萌发至3 cm时,电泳条带开始变淡并产生新的或加深在其附近小分子条带。除本课题组[4]前期测得的较高的中性和酸性蛋白酶活性外,也有相关报道发现,在大豆萌发过程中,蛋白酶C1、一种新的丝氨酸蛋白酶和半胱氨酸蛋白酶D3[12]等蛋白酶均能够有效地降解β-伴球蛋白的α’及α亚基[10,11]。酸奶是典型的酸致蛋白凝胶,大豆蛋白的适度降解对大豆酸奶的质构、风味和理化特性改善有重要作用。
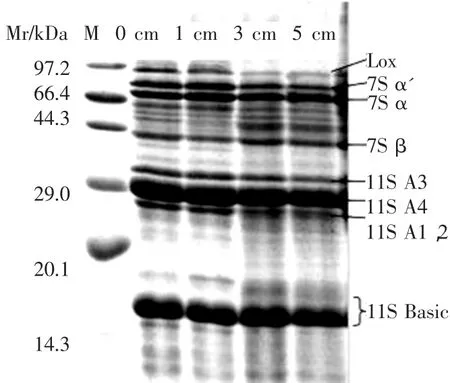
图1 萌发大豆豆浆中蛋白亚基组成变化图谱Figure 1 Degradation of soybean protein in the germinated soybeanmilk
2.1.2 内源酶降解对豆浆中胰蛋白酶抑制剂的影响 大豆胰蛋白酶抑制因子(soybean trypsin inhibitor,STI)是大豆主要抗营养因子之一,能引起胰腺增生和抑制动物生长,生大豆40%的抗营养作用是STI引起的[12]。由图2可知,内源酶的降解作用可以显著降低豆浆中胰蛋白酶抑制因子的活性并提高其热敏感性。当芽长为3 cm时,生豆浆中TIA降低了32%,而豆浆灭菌即100℃加热15min后,芽长1 cm和3 cm的大豆豆浆TIA分别降低了48%和40%,而未经萌发的大豆豆浆TIA只降低了30%。
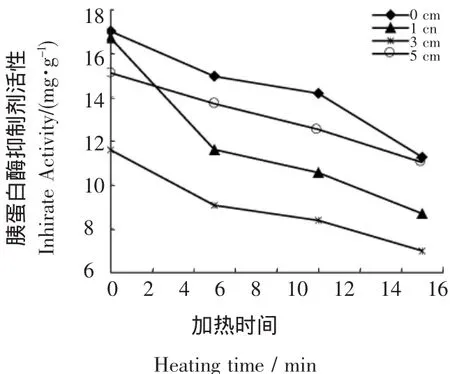
图2 萌发大豆豆浆中胰蛋白酶抑制剂活性变化Figure 2 The trypsin inhibition activity(TIA)of the germinated soybeanmilk
2.2 萌发大豆内源酶对大豆酸奶的影响
2.2.1 内源酶降解对大豆酸奶pH、酸度和持水力的影响
内源酶作用对大豆酸乳pH、酸度及持水力的影响见表1。萌发大豆酸奶的酸度随芽长增长即内源酶作用时间延长而显著性增大,pH也随芽长增长呈现降低的趋势,这是因为大豆萌发过程中,其内源蛋白酶限制性降解部分蛋白质,释放出大豆多肽和游离氨基酸[4],同时产生还原糖和矿物质等有利于缩短发酵剂的适应期,加快乳酸菌的增殖产酸[13,14],导致酸度上升、pH下降。凝固型酸奶的持水力是评价酸奶凝胶网络体系稳定性的重要指标之一。如表1所示,后熟24 h后,芽长为0,1,3 cm的大豆所制酸奶持水力较高,均在90%以上,其中1 cm最大,芽长5 cm的大豆所制酸奶持水力最低(<90%),3 cm次之。低持水力主要是由于发酵酸乳不稳定的三维凝胶网络结构而导致的[15],当大豆萌发至5 cm时,大豆蛋白亚基相对较多的被降解为肽链、多肽或氨基酸,导致蛋白凝胶中胶粒连接减弱,不能很好地将水分包裹在它的三维网络结构内,所以5 cm样品酸奶凝胶显示出较低的持水力。
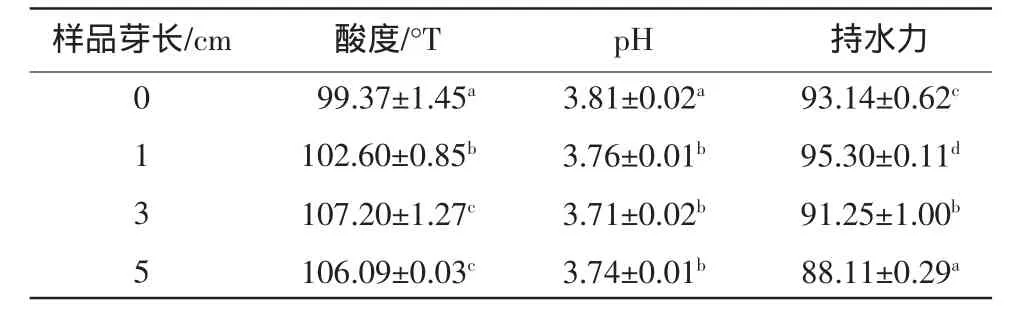
表1 萌发大豆酸奶的pH、酸度和持水力覮Table1 pH,acidity and water-holding capacity of germinated soy yogurt(n=3)
2.2.2 内源酶降解对大豆酸奶流变性质的影响
(1)频率扫描:应用线性黏弹性理论,研究萌发的即经内源酶作用的大豆酸奶在小变形范围内的黏弹性质及变化规律。不同芽长的大豆酸奶样品的频率扫描结果见图3。G'是在振动测试中每一个变形周期内的储能模量,它可以表征酸奶网络结构的强度,代表酸奶的弹性;G''是酸奶在变形过程中所损失的能量,代表酸奶的黏性。在0.1至10 Hz的线性黏弹性范围内,大豆酸奶样品的G'和G''显示出较强的频率敏感性,数值均随扫描频率增加而增大,且G'值均高于G''值,表现出类固体的流变特性[16]。大豆经过萌发后,大豆酸奶的G'及G"值均显著下降,且随着样品芽长增加,下降趋势更明显。有研究[17]表明,G'数值的降低,凝胶结构表现得越为疏松。由此可见,内源酶的作用可显著降低大豆酸奶的黏弹性,即是说大豆酸奶凝胶的抗压抗震能力减弱且黏度降低。这应该是萌发过程中,大豆内源酶的酶解作用使大分子蛋白部分降解,同时将结构致密的大豆球蛋白结构变得疏松,进而使萌发大豆制成的大豆酸奶凝胶三维网络结构变得更加疏松柔软,胶束之间的缠黏能力减弱导致的。
(2)剪切扫描:采用Herschel-Bulkley模型对萌发大豆酸奶的剪切速率与剪切应力曲线进行拟合分析(表2)。该模型可由式(2)来表示。

式中:
τ——剪切应力,Pa;
τ0——屈服应力,Pa;
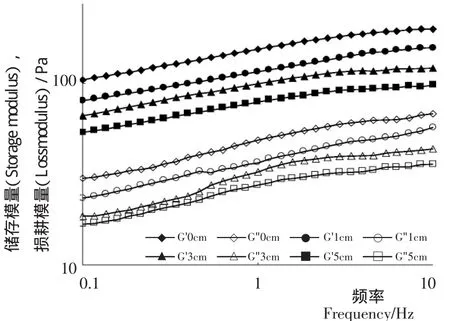
图3 萌发大豆酸奶的频率扫描曲线Figure 3 Frequency sweeps of germinated soy yogurt
γ——剪切速率,1/s;
к——稠度系数,Pa·sn;
n——流动行为指数。
如表2所示,所有样品的R2均高于0.95,表明Herschel-Bulkley模型适合用于大豆酸奶剪切扫描曲线的拟合分析;n值均小于1表明4种发酵酸乳均为非牛顿流体—假塑性流体[18]。大豆萌发后,发酵豆乳的屈服应力显著下降,分别从0 cm样品中的8.08 Pa下降至5 cm样品中的2.90 Pa。Harte等[19]研究表明,酸奶的屈服应力数值与感官品评中的“硬度”有密切关系,即与未萌发大豆酸奶相比萌发大豆酸奶的口感表现更为柔软。大豆经过萌发后,大豆酸奶的流动行为指数n值增大,稠度系数κ表观黏度降低,这与频率扫描样品黏弹性降低的结果一致。触变性是流体受到剪切时,稠度变小,停止剪切时,稠度又增加,即一触即变的性质。触变性流体的机理可以理解为随着剪切应力的增加,粒子间结合的结构受到破坏,黏度降低;当作用力停止时粒子间结合的构造逐渐恢复原样,但需要一段时间[20]。因此,剪切速率减少时的曲线与增加时的曲线不重叠,形成了与流动时间有关的滞后曲线(流变环)。随芽长增长,0,1,3 cm的酸奶样品触变性依次显著性增大,表明在同样的剪切作用下芽长越长的酸奶样品质构破坏的越彻底,所以萌发可使大豆酸奶口感变得柔软爽滑。至于5 cm的酸奶样品触变性最小应该是大豆内源蛋白酶把太多的大豆蛋白降解为较小分子[21],使酸奶的凝胶结构变得非常脆弱,在剪切初期结构就被破坏且蛋白胶束间交联比较困难,结构很难恢复的原因。
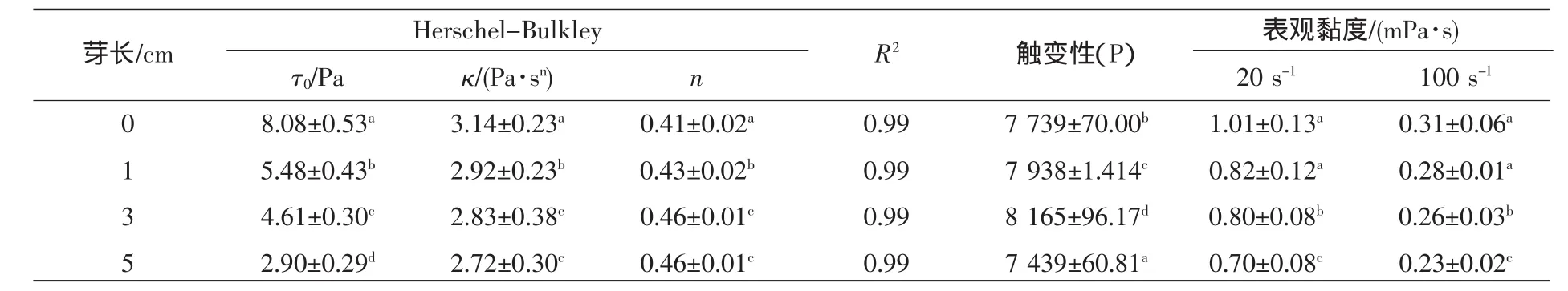
表2 萌发大豆酸奶的流变参数覮Table2 Rheological parameters of germinated soy yogurt(n=2)

图4 内源酶降解对大豆酸奶微观结构的影响Figure 4 Effectof Endogenous enzyme enzymolysis on themicrostructure of sogurt
(3)内源酶降解对大豆酸奶微观结构的影响:激光共聚焦显微镜(CLSM)经常用于乳液及凝胶的微观结构研究。图4为不同芽长的萌发大豆酸奶样品的微观结构图,本组图片中蛋白显现红色,油脂显现绿色。从图4中可以看到明显的絮状凝胶网络结构。酸奶凝胶的微观结构在很大程度上决定了其持水力、质构、流变等特性。随着萌发程度的增加,大豆酸奶三维凝胶网络结构的间隙显著增大,结构变得更为疏松。0,1 cm酸奶样品网络结构致密,这是由于大豆蛋白的分子结构较致密,分子刚性较强,因此所形成的蛋白凝胶结构非常紧密而刚硬,因此显示出很高的硬度、黏附性及黏弹性流变特性[22],这与流变数据吻合;它们较高的持水力也可以用致密的凝胶结构来解释。大豆芽长越长,其内源酶降解作用越强。内肽酶能将大豆蛋白降解为长短不一的多肽链,使大豆蛋白分子结构变得疏松,从而改善大豆酸奶中的蛋白胶束交叉连接情况,使交联网络结构中的羟基、羧基等亲水基团更多地暴露出来,亲和更多水分子进入大豆蛋白凝胶的网络体系[23],使凝胶的网络结构变得更为疏松柔软,这与3,5 cm酸奶样品具有较低的屈服应力τ0也相对应。此外,当芽长为5 cm时,大豆内蛋白被过度降解,蛋白分枝减少,生成的蛋白链末端亲水基团较多,使大豆酸奶中大豆蛋白的交联作用变弱,产生大的间隙孔洞,凝胶体系变得脆弱易破坏,结构不易恢复,使凝胶的持水力降低。
3 结论
(1)SDS-PAGE电泳分析不同芽长的大豆豆浆中蛋白质亚基组成的变化情况表明:萌发过程中,在内源蛋白酶作用下,大分子量蛋白随芽长增长而逐渐被降解产生小分子的蛋白。大豆在芽长为3 cm时,β-伴球蛋白(7S)的α'、α亚基及大豆球蛋白(11S)酸性亚基开始被降解;对大豆胰蛋白酶抑制因子活性的分析结果表明:内源酶的降解降低了豆浆中胰蛋白酶抑制剂的活性,提高了其热敏感性,提高了豆浆或大豆酸奶的安全性和营养性。
(2)大豆萌发能显著提高大豆酸奶的酸度、降低pH,并影响酸奶的持水力;CLSM微观图像和流变特性分析表明大豆萌发可使大豆酸奶凝胶网络结构变得细腻,显著降低大豆酸奶的屈服应力和表观黏度,对大豆酸奶的质构有很好的改善作用。
(3)试验表明芽长为3 cm时,大豆酸奶的胰蛋白酶抑制剂活性最低,理化特性、流变特性最好,微观结构细腻,综合品质最好。
1 余保宁,毛军,李理,等.3种酶解方法对大豆酸奶品质的影响[J].食品与发酵工业,2007,33(5):137~142.
2 王汝毅,陈福生,赵山,等.酶解豆浆制酸豆奶的研究 [J].食品科学,2005,26(10):267~269.
3 Feng S,Saw C L,Lee Y K,et al.Novel process of fermenting black soybean[Glycinemax(L.)Merrill]yogurtwith dramatically reduced flatulence-causing oligosaccharides but enriched soy phytoalexins[J].Journal of Agricultural and Food Chemistry,2008,56(21):10 078~10 084.
4 Yang M,Fu J,Li L.Rheological characteristics and microstructure of probiotic soy yogurt prepared from germinated soybeans[J].Food Technology and Biotechnology,2012,50(1):73~80.
5 Yang M,Li L.Physicochemical,textural and sensory characteristics of probiotic soy yogurt prepared from germinated soybean[J].Food Technology and Biotechnology,2010,48(4):490~496.
6 杨媚,李理.应用萌发大豆生产益生菌发酵豆乳的研究[J].食品工业科技,2010,31(11):206~209.
7 刘占祥,王琪.酸牛奶的加工技术[J].宁夏农林科技,2000(5):62~63.
8 Laemm li U K.Cleavage of structural proteins during the assembly of the head of bacteriophage T4[J].Nature,1970,227(5 259):680~685.
9 Kakade M,Rackis J,McGhee J,et al.Determination of trypsin inhibitor activity of soy products:a collaborative analysis of an im-proved procedure[J].Cereal Chemistry,1974,51(3):376~382.
10 杨媚,李理.萌发大豆制备益生菌发酵豆乳流变特性的研究[J].食品工业,2011(5):8~11.
11 Tang CH,Liu F.Cold,Gel-like soy protein emulsions by microfluidization:Emulsion characteristics,rheological and microstructural properties,and Gellingmechanism[J].Food Hydrocolloids,2013(30):61~72.
12 谷春梅,韩玲玲,曲洪生,等.大豆胰蛋白酶抑制因子的研究进展[J].大豆科学,2012,31(1):149~151.
13 王莘,王艳梅,闵卫红,等.大豆萌发期功能性营养成分测定与分析[J].中国粮油学报,2004,18(4):30~32.
14 王莘,王艳梅,董浩.豆类萌发期矿物质元素和糖类含量变化的研究[J].扬州大学学报:农业与生命科学版,2003,24(2):72~74.
15 Bau H M,Villaume C,Nicolas JP,et al.Effect of germination on chemical composition,biochemical constituents and antinutritional factors of soya bean(Glycinemax)seeds[J].Journal of the Science of Food and Agriculture,1997,73(1):1~9.
16 Donkor ON,Henriksson A,Vasiljevic T,et al.Rheological properties and sensory characteristics of set-type soy yogurt[J].Journal of Agricultural and Food Chemistry,2007,55(24):9 868~9 876.
17 Lucey J,Van Vliet T,Grolle K,et al.Properties of acid casein gels made by acidification with glucono-δ-lactone.1.Rheological properties[J].International Dairy Journal,1997,7(6):381~388.
18 Lucey J,Singh H.Formation and physical properties of acid milk gels:a review[J].Food Research International,1997,30(7):529~542.
19 Harte F,Clark S,Barbosa-Cánovas G.Yield stress for initial firmnessdetermination on yogurt[J].Journalof Food Engineering,2007,80(3):990~995.
20 Barnes H A.Thixotropy-a review[J].Journal of Non-Newtonian Fluid Mechanics,1997,70(1):1~33.
21 Qi X,Chen R,Wilson K A,et al.Characterization of a soybean[beta]-conglycinin-degrading protease cleavage site,Plant Physiol[J].Plant Physiology,1994,104(1):127~133.
22 王洪晶,华欲飞.大豆分离蛋白凝胶研究进展[J].粮食与油脂,2005,2(3):3~5.
23 李锋,华欲飞.大豆酸奶的微结构以及分形特征[J].食品科学,2005,26(7):110~115.
猜你喜欢
世界最新医学信息文摘(2020年68期)2020-12-25
现代园艺(2020年7期)2020-04-22
湖北农业科学(2019年20期)2019-12-11
延安大学学报(医学科学版)(2019年1期)2019-03-29
农业与技术(2018年5期)2018-04-25
农产品加工(2017年19期)2017-11-14
上海农业学报(2016年2期)2016-10-27
华东理工大学学报(自然科学版)(2015年4期)2015-12-01
动物营养学报(2015年10期)2015-12-01
云南中医学院学报(2014年3期)2014-07-31
