
重组大肠杆菌表达双功能谷胱甘肽合成酶的发酵工艺研究
2013-10-15 10:14王德正李志敏
化学与生物工程 2013年8期
张 松,王德正,李 娓,李志敏,叶 勤
(华东理工大学 生物反应器工程国家重点实验室,上海200237)
谷胱甘肽(Glutathione,GSH)是一种具有多种重要生理功能的活性三肽,由谷氨酸、半胱氨酸和甘氨酸经肽键缩合而成。GSH在维持体内氧化还原平衡、迅速增强机体免疫力、促进消化系统及呼吸系统机能、抗肿瘤等方面发挥着重要作用[1],广泛应用于医药、食品、体育运动等领域。GSH的生物合成分为两步[2]:第一步,谷氨酸的γ-羧基与半胱氨酸的氨基在γ-谷氨酰半胱氨酸合成酶(GSHⅠ,即γ-GCS)的作用下催化形成肽键,合成γ-谷氨酰半胱氨酸;第二步,在谷胱甘肽合成酶(GSHⅡ,即GS)的作用下与甘氨酸反应生成谷胱甘肽。其中GSHⅠ是谷胱甘肽合成的限速酶,受到还原型谷胱甘肽的反馈抑制[3],而谷胱甘肽对GSHⅡ没有反馈抑制[4]。
最近研究报道,除了GSHⅠ和GSHⅡ外,在某些菌[5,6](如无乳链球菌、李斯特氏菌)中还存在另外一种特殊的双功能谷胱甘肽合成酶(GshF),同时具有GSHⅠ和GSHⅡ的活性,对产物抑制不敏感,在谷胱甘肽生物合成中有着重要的应用价值。目前的研究主要集中在新酶的开发和分子机制方面,未涉及发酵及诱导表达GshF的工艺优化。
作者在此以重组表达嗜热链球菌gshF基因的大肠杆菌BL21为对象,进行了重组菌BL21/pET-gshF 5L罐的发酵研究,分别考察了不同培养基、诱导剂种类、诱导时机、诱导温度、补料流加碳源类型等因素对菌体生长、GshF蛋白表达及其酶活的影响,拟为GshF在谷胱甘肽生物合成中的应用奠定基础。
1 实验
1.1 菌株
重组大肠杆菌BL21/pET-gshF,自行构建;目的基因片段gshF来自嗜热链球菌。
1.2 培养基
种子培养基:含30mg·L-1卡那霉素的LB培养基。
基本 培 养 基 (g·L-1):Na2HPO4·12H2O 15.12,KH2PO43,NH4Cl 3,MgSO4·7H2O 0.5,NaCl 0.5,酵母提取物5。葡萄糖5,115℃单独灭菌30min;CaCl20.011,单独灭菌。1%维生素B1用微孔滤膜(0.22μm)过滤除菌,待培养基灭菌冷却后按0.12mL·L-1的比例加入。微量元素混合溶液(μmol·L-1,FeCl350,CaCl220,MnCl2·4H2O 10,ZnSO4·7H2O 10,CoCl2·6H2O 2,CuCl2·2H2O 2,NiCl2·6H2O 2,Na2MoO4·5H2O 2,Na2SeO3·5H2O 2,H3BO32)用微孔滤膜(0.22μm)过滤除菌,待培养基灭菌冷却后按0.12mL·L-1的比例加入。
补料培养基Ⅰ(g·L-1):葡萄糖500,MgSO4·7H2O 25。
补料培养基Ⅱ(g·L-1):甘油250,MgSO4·7H2O 25。
自诱导培养基(g·L-1):蛋白胨10,酵母提取物5,Na2HPO47.1,KH2PO46.8,(NH4)2SO46.6,MgSO40.12,甘油20,葡萄糖0.5,乳糖2,琥珀酸钠4.05,按0.12mL·L-1的比例加入微量元素储备液。
1.3 培养方法
1.3.1 种子培养
一级种子培养:将-20℃保存的甘油管菌株接种到装有30mL LB培养基的250mL三角瓶中,加卡那霉素至终浓度为30mg·L-1,于37℃、220r·min-1培养6h。
二级种子培养:将上述种子液按1%接种量接种到装有125mL LB培养基的500mL三角瓶中,于37℃、220r·min-1培养8h。
1.3.2 分批发酵
将二级种子液按5%接种量接种到装有2.5L发酵培养基的5L发酵罐(BIOTECH-BG-5型,上海保兴生物设备公司)中,于37℃进行分批发酵。用NaOH控制pH值为7.0左右,通气量为1VVm,调节搅拌转速控制溶氧在30%以上。
1.3.3 补料分批发酵
发酵罐装液量为2.5L,接种量为5%,用25%氨水控制pH值为7.0,通气量为1VVm,调节搅拌转速保持溶氧在30%以上。初糖耗尽、溶氧跳升后通过指数流加(控制比生长速率0.3h-1)进行补料,分别以葡萄糖和甘油两种不同碳源流加。在对数生长初期及中后期补加IPTG或者乳糖进行诱导。
1.4 分析方法
1.4.1 还原型谷胱甘肽的含量测定
用HPLC法检测GSH含量。使用C18色谱柱(4.6mm×150mm),流动相为0.01mol·L-1Na2HPO4和0.05mol·L-1K2HPO4溶液(含0.01mol·L-1庚烷磺酸钠,pH值3.0)。检测时流动相和甲醇按95∶5的比例进柱,柱温30℃,流速0.6mL·min-1。
1.4.2 GshF蛋白定量
按照分子克隆对发酵样品进行蛋白凝胶电泳,并以蛋白Marker和牛血清蛋白为标准确定目标蛋白GshF的大小和含量[7]。
1.4.3 酶活分析
取1mL菌液,离心收集菌体,加pH值为8.5的0.15mmol·L-1Tris-HCl、0.2mmol·L-1EDTA,0.1mmol·L-1KCl至1mL,在冰浴中超声破碎后,于12 000r·min-1离心10min;取上清液500μL,加入40mmol·L-1Glu、40mmol·L-1Gly、20mmol·L-1Cys、40mmol·L-1MgCl2、40mmol·L-1ATP,30℃反应30min;加入与反应液等体积的20%三氯乙酸终止反应,然后用HPLC检测生成的GSH含量。
酶活单位(U)为:每分钟转化底物生成1mg GSH的酶量。
比酶活(U·g-1)为:每克GshF蛋白每分钟转化底物生成1mg GSH的酶量。
1.4.4 菌体密度的测定
将待测发酵液稀释至一定浓度后,用紫外分光光度计在600nm波长下测定发酵液的吸光度值。发酵液的细胞密度即为吸光度值与稀释倍数的乘积。
1.4.5 其它物质的测定
葡萄糖、甘油、乙酸、乳糖、琥珀酸的含量均采用HPLC法检测[8]。色谱柱为 Aminex HPX-87H,用紫外检测器和视差折光检测器检测;流动相为5mmol·L-1H2SO4,流速0.6mL·min-1,检测温度65℃。
2 结果与讨论
2.1 自诱导培养基分批培养
分批培养是一种简单的培养方式,可以初步获得菌体生长和重组蛋白表达的基本规律,为后续优化提供借鉴,但分批培养后期由于营养物质的消耗,难以实现细胞的高密度培养和蛋白的高表达。Studier[9]在自诱导培养基中培养重组大肠杆菌,目标蛋白得到很好的表达。因此,本研究首先考察37℃时使用自诱导培养基进行分批培养时的菌体生长、营养物质代谢和GshF蛋白表达,结果见图1。

图1 分批培养时菌体生长、代谢情况和GshF表达量Fig.1 The cell growth,metabolism and GshF expression in batch culture
由图1可见,在37℃自诱导培养基中,前8h菌体生长迅速,乙酸积累了1.9g·L-1;培养10h时甘油耗尽,OD600达到最大值18.5,并开始利用积累的乙酸,菌体生长基本停滞;培养10h时GshF表达量达到最大值0.26g·L-1,随后随着菌体浓度的减小,出现下降;初始葡萄糖只有0.5g·L-1,因此很快就被完全消耗,过程中基本检测不到。
2.2 以葡萄糖为碳源流加实验
补料流加培养是生产重组蛋白最常用的方式,能使基质浓度保持在较低水平,有效地抑制副产物的积累,提高外源蛋白的表达效率。由于自诱导培养基中使用了较多的复合成分,增加了生产成本,因此补料流加工艺以添加少量复合成分的基本培养基(添加酵母提取物的M9培养基)为初始发酵培养基,考察IPTG诱导温度和诱导时机、乳糖作为碳源和诱导剂时对菌体生长和蛋白诱导表达的影响。
2.2.1 IPTG诱导温度的影响
采用初始发酵培养基在37℃下进行菌体培养,4.8h开始控制比生长速率为0.3h-1,指数流加补料培养基Ⅰ,10.5h添加0.5mmol·L-1IPTG进行诱导,比较诱导温度为37℃、30℃时的菌体生长和GshF表达量,结果见图2。

图2 IPTG诱导温度对菌体生长、GshF表达量的影响Fig.2 Effect of induction temperature on cell growth,GshF expression
以添加酵母提取物的M9培养基为初始发酵培养基,并限制性流加葡萄糖的补料分批培养过程中未检测到残余葡萄糖和乙酸生成。由图2可见,37℃进行诱导时,GshF表达量随着诱导时间的延长而增加,比酶活最大达到74.6U·g-1;30℃诱导时,比酶活最大为70.9U·g-1。表明,诱导温度对菌体生长和GshF的表达影响不大。
2.2.2 IPTG诱导时机的影响
考虑到过早诱导可能增加表达外源蛋白的代谢负荷而抑制菌体生长,从而影响GshF表达量。因此尝试推迟诱导剂添加时间,在OD600达到50的时候加入0.5mmol·L-1IPTG 进行诱导。结果发现,添加IPTG诱导后,菌体在最初的2h内保持高速指数生长,之后生长速率减慢,OD600在18h达到最大值96.8,但GshF表达量最高只有2.13g·L-1(表1 )。表明推迟诱导剂添加时间虽然有利于菌体生长,但GshF表达量并没有显著提高。

表1 诱导时机对菌体生长、GshF表达量的影响Tab.1 Effect of induction time on cell growth,GshF expression
2.2.3 乳糖诱导的影响
维持初始发酵培养基和流加培养基不变,在9h OD600达到33时停止流加葡萄糖以防止葡萄糖对乳糖诱导的抑制作用,饥饿处理0.5h至葡萄糖完全耗尽,然后恒速(40mL·h-1)流加20%乳糖进行诱导,结果见图3。

图3 乳糖诱导对菌体生长、GshF表达量的影响Fig.3 Effect of induction by lactose on cell growth,GshF expression
由图3可见,虽然乳糖既有诱导作用,又能作为碳源被菌体代谢,但作为碳源,和葡萄糖相比还是难以利用,菌体生长几乎停止;GshF表达量也远远达不到IPTG诱导的效果,最高值仅为0.19g·L-1。这可能是因为,乳糖只有借助于乳糖透酶的作用进入到菌体细胞内,并经过β-乳糖苷酶的作用转化为异乳糖才会起到诱导剂的作用[10]。虽然乳糖诱导的GshF表达量低,但其比酶活远远高于IPTG诱导时的比酶活,在诱导3h时达到最大值359U·g-1,即出现了蛋白含量低、酶活高的现象[11]。原因可能是IPTG诱导产生GshF较乳糖快,使得蛋白来不及正确折叠,从而形成了部分包涵体,导致酶活较低。
2.3 以甘油为碳源流加实验
葡萄糖作为一种很容易被菌体利用的碳源,能使菌体短时间内大量增殖,但过快的生长速度并不利于外源蛋白的表达,而且葡萄糖过量也容易引起乙酸等代谢物的积累,这对蛋白表达也是不利的。甘油是一种较慢被大肠杆菌利用的碳源,进行甘油补料流加不仅产生较少的副产物乙酸,而且可以实现重组大肠杆菌的高密度发酵,因此初步尝试在生长阶段和诱导阶段均流加20%的甘油,结果见图4。
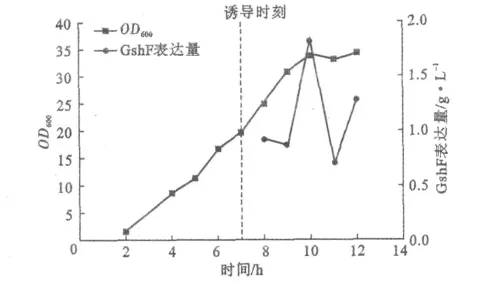
图4 诱导前后均流加甘油时的菌体生长、GshF表达量Fig.4 Cell growth,GshF expression with glycerol fed before and after induction
由图4可见,流加甘油,OD600最终达到34.3;在OD600为20时开始诱导表达,GshF表达量在诱导3h后达到最大值1.83g·L-1,随后出现波动降低。这一方面是由于发酵液中蛋白酶的作用,也与后期菌浓不再增加有关。甘油流加工艺中GshF表达量稍低于葡萄糖流加工艺,且后期菌浓停止增加,不利于继续提高蛋白表达和保持蛋白活性。
3 结论
在5L发酵罐中培养重组大肠杆菌BL21/pET-gshF,考察了不同培养基、诱导方式和流加碳源对菌体生长、GshF蛋白表达的影响,结果表明:
(1)使用自诱导培养基分批发酵,能够代替添加IPTG实现GshF蛋白的自发诱导,但表达量较低,培养基成本过高。
(2)以M9培养基加5g·L-1酵母提取物为初始发酵培养基,葡萄糖作为流加碳源并在OD600达到30时加入IPTG诱导,诱导温度对菌体生长和GshF表达量影响均不大;推迟IPTG诱导时机,菌体生长量大幅提高,但GshF表达量并没有显著提高;在使用乳糖作为诱导剂进行诱导时,GshF表达量较低,但比酶活高于IPTG诱导。乳糖成本低,需要进一步优化工艺使其能在保持高酶活的前提下提高蛋白表达量。
(3)以甘油为碳源进行流加培养和蛋白表达,也实现了GshF的高效表达,但后期菌体生长停滞,蛋白降解严重,因此甘油作为碳源工艺条件还不成熟,需要进一步探索。
[1]钱小明,吴学豪.还原型谷胱甘肽在急危重症中的应用[J].中国急救医学,2003,23(6):411-412.
[2]Blach K C.The synthesis of glutathione in isolated liver[J].Biol Chem,1949,179(3):1245-1254.
[3]Ikner A,Shiozaki K.Yeast signaling pathways in the oxidative stress response[J].Mutat Res,2005,569(1-2):13-27.
[4]毛珍,裘娟萍.酵母菌中谷胱甘肽的主要生理功能及其代谢调控[J].微生物学杂志,2005,25(1):94-96.
[5]Janowiak B E,Griffith O W.Glutathione synthesis in Streptococcus agalactiae.One protein accounts for gamma-glutamylcysteine synthetase and glutathione synthetase activities[J].J Biol Chem,2005,280(12):11829-11839.
[6]Gopal S,Borovok I,Ofer A,et al.A multidomain fusion protein in Listeria monocytogenes catalyzes the two primary activities for glutathione biosynthesis[J].J Bacteriol,2005,187(11):3839-3847.
[7]邓永康,吴民泸,刘盛邦,等.乳糖诱导重组尿酸酶基因在大肠杆菌中的表达[J].中国生物工程杂志,2009,29(7):74-79.
[8]Liu Y,Wu H,Li Q,et al.Process development of succinic acid production by Escherichia coli NZN111using acetate as an aerobic carbon source[J].Enzyme and Microbial Technology,2011,49(5):459-464.
[9]Studier F W.Protein production by auto-induction in high density shaking cultures[J].Protein Expression and Purification,2005,41(1):207-234.
[10]杨书慧,赵胜军,刘军,等.乳糖诱导甜蛋白Monellin在大肠杆菌中的表达[J].中国生物工程杂志,2008,28(3):53-58.
[11]余永恒,李清雄,王革非.乳糖诱导重组人睫状神经营养因子在大肠杆菌中的可溶性表达[J].中国生物制品学杂志,2007,20(1):43-46.
猜你喜欢
江苏卫生保健(2022年5期)2022-05-24
中国医学影像学杂志(2021年6期)2021-08-13
幸福(2018年33期)2018-12-05
能源(2017年7期)2018-01-19
中学科技(2017年11期)2017-12-26
天然产物研究与开发(2016年6期)2016-06-05
中国民族医药杂志(2016年4期)2016-05-09
中国继续医学教育(2015年3期)2016-01-06
饮食科学(2015年4期)2015-11-28
化工生产与技术(2014年4期)2014-02-27
