
毛乌素沙地南缘沙丘生物结皮对凝结水形成和蒸发的影响
2013-12-09 06:30尹瑞平吴永胜田秀民李泽坤苗恒录
生态学报 2013年19期
尹瑞平,吴永胜,*,张 欣,哈 斯,田秀民,李泽坤,王 健,苗恒录,任 杰
(1.水利部牧区水利科学研究所,呼和浩特 010020; 2. 北京师范大学 资源学院,北京 100875)
毛乌素沙地南缘沙丘生物结皮对凝结水形成和蒸发的影响
尹瑞平1,吴永胜1,*,张 欣1,哈 斯2,田秀民1,李泽坤1,王 健1,苗恒录1,任 杰1
(1.水利部牧区水利科学研究所,呼和浩特 010020; 2. 北京师范大学 资源学院,北京 100875)
在水分极度匮乏的荒漠生态系统,凝结水是除降雨之外最重要的水分来源之一,它对荒漠生态系统结构、功能和过程的维持产生重要的影响。为探明半干旱沙区生物结皮表面的凝结水形成和蒸发特征,采用自制的微型蒸渗计(直径7 cm、高5 cm的PVC管)实验观测了不同类型地表(裸沙、浅灰色藻类结皮、黑褐色藻类结皮和苔藓结皮)对凝结水形成和蒸发的影响。结果表明:(1)观测期间共有20次凝结水形成记录,除降雨天气外,几乎每天都能观测到水分凝结现象;(2)不同类型地表凝结水总量依次为(1.998±0.075),(2.326±0.083),(2.790±0.058)和(3.416±0.068) mm,生物结皮表面的凝结水总量显著大于裸沙(Plt;0.05);随生物结皮的发育,不同类型生物结皮表面的凝结水总量呈增加的趋势,凝结水总量之间差异显著(Plt;0.05);观测期间不同类型地表日平均凝结水量依次为(0.100±0.003),(0.116±0.004),(0.140±0.002)和(0.171± 0.003) mm,不同类型地表日平均凝结水量之间差异极显著(Plt;0.01);(3)凝结水形成过程的观测结果显示,凝结水19:00开始形成,23:00—凌晨1:00形成不明显,1:00—7:00继续形成,除浅灰色藻类结皮外,太阳升出后在黑褐色藻类结皮和苔藓结皮表面继续形成少量的凝结水;凝结水7:30开始蒸发,10:30到11:00之间结束蒸发,凝结水在裸沙和浅灰色藻类结皮中的保持时间显著大于黑褐色藻类结皮和苔藓结皮中的保持时间(Plt;0.05);(4)凝结水的形成受大气温度、地表温度、空气相对湿度和大气地表温度差等气象因素的影响,但其形成过程不与某一个气象因素呈简单的线性关系。
毛乌素沙地;生物结皮;凝结水;蒸发
水分是影响沙地生态系统植被生产力和生产的最大限制因子[1]。在水资源极度匮乏的沙地环境,凝结水是除降雨之外的重要水分来源[2]。虽然沙生灌木、半灌木和草本植物对凝结水和雾水的直接利用价值有限[1],但对低矮植物(如苔藓)、昆虫和微生物等生物体而言,凝结水却是他们赖以生存的重要水源之一[3]。此外,凝结水在沙地生态系统水量平衡中的贡献是相当可观的。毛乌素沙地南缘沙区年凝结水量的研究结果表明,凝结水占该区域年均降雨量的12.6%之多[4],这在干旱沙区是十分可贵的,因此有关干旱沙区凝结水形成和蒸发特征的研究具有重要的生态学和水文学意义。
国外学者对凝结水的研究较早,围绕凝结水的观测方法[5- 6]、形成机理[7]、影响因素[8- 10]和季节变化[11]等方面开展了丰富的研究。我国学者对凝结水的研究也较多,研究地区分布在古尔班通古特沙漠[12]、腾格里沙漠沙坡头地区[13- 15]、库布齐沙漠[16]、科尔沁沙地[17- 18]和毛乌素沙地[2]等地。
生物结皮(由微生物、蓝绿藻、地衣、苔藓等微小生物体与沙粒胶结形成的有机复合体)广泛存在于荒漠生态系统,其分布面积几乎占到该地区生物覆盖率的40%[19]。我国干旱和半干旱沙区固定、半固定沙丘上生物结皮广泛发育[20]。随着生物结皮水文学研究的不断深入,生物结皮对凝结水形成的影响引起了国内学者的关注,并在古尔班通古特沙漠和腾格里沙漠沙坡头地区生物结皮对凝结水的影响方面取得了进展[12- 14],但已有的研究结果多集中在干旱或极端干旱沙区,鲜见半干旱沙区生物结皮对凝结水影响的相关报道,研究现状与我国半干旱沙区生物结皮广泛发育的现状是不对称的。基于此,本文选择地处半干旱区的毛乌素沙地南缘沙区,观测生物结皮对凝结水形成和蒸发的影响,丰富我国不同气候区(极端干旱、干旱到半干旱沙区)生物结皮水文学相关研究内容的同时,旨在为脆弱生态系统的合理管理、利用和生态修复提供基础数据。
1 研究区概况及方法
1.1 研究区概况
研究区位于毛乌素沙地南缘,是鄂尔多斯高原向陕北黄土高原的过渡区,地理坐标为108°50′54″—108°58′00″E,37°38′42″—37°42′42″N,海拔1350 m,属半干旱气候。研究区多年平均降水量为394.7 mm,降水集中在夏、秋两季,降水变率大,最大降水量达744.6 mm(1964年),最小降水量仅205 mm(1965年);多年平均蒸发量为2484.5 mm,是多年平均降水量的6.32倍。地貌景观以流动、半固定、固定沙丘与湖盆滩地相间。人工植被恢复区固定、半固定沙丘上生物结皮广泛发育,迎风坡和背风坡以浅灰色藻类结皮和黑褐色藻类结皮为主,丘间地则以苔藓结皮为主。不同类型生物结皮的基本特征如下:浅灰色藻类结皮(LC)表面较光滑,颜色呈浅灰色,厚度约3—5 mm,抗剪强度为0.3—0.5 kg/cm2,主要生物组分为微鞘藻(Microcoleusvaginatus)、颤藻属(Oscillatoriaspp.)、鞘丝藻属(Lyngbyaspp.);黑褐色藻类结皮(DC)表面较粗糙,呈黑褐色,厚度约5—8 mm,抗剪强度为1.0 kg/cm2左右,主要生物组分为颤藻属、鞘丝藻属;苔藓结皮(MC)表面长有苔藓植物,干旱缺水时呈深颜色,雨后呈鲜绿色,厚度和抗剪强度均显著提高,分别为10—12 mm和2.1 kg/cm2左右,主要苔藓植物为双色真藓(BryumdichotomumHedw.),真藓(BryumargenteumHedw.),土生对齿藓(Didymodonvinealis(Brid.)Zand)。沙丘沉积物机械组成以细沙为主,中沙次之[20]。研究区属暖温性草原带,植被覆盖度低,主要天然植物有油蒿(ArtemisiaordosicaKrasch.)、沙米(AgriophyliumsquarrosumMoq.)、软毛虫实(CorispermumpuberulumIljin)、沙竹(Psammochloavillosa(Trin.) Bor.)等。
1.2 方法
目前为止,国际上没有通用的凝结水观测方法,采用微型蒸渗计进行实验观测的较常见[12- 14]。本文采用自制微型蒸渗计(由直径7 cm,高5 cm的PVC管制成)观测不同类型地表(裸沙(BS)、浅灰色藻类结皮(LC)、黑褐色藻类结皮(DC)和苔藓结皮(MC))对凝结水形成和蒸发的影响。采样时在研究区选取具有代表性的半固定沙丘,按照丘顶、背风坡中下部、丘间地和迎风坡中上部的顺序选择生物结皮样地,依次代表裸沙、黑褐色藻类结皮、苔藓结皮和浅灰色藻类结皮。采样前用喷壶湿润生物结皮表面,以减少对地表的扰动。具体操作是把PVC管垂直按入结皮表面,采集原状土,用防水胶带把PVC管底部封住。每种地表类型采集4份作为重复。实验前把原状土晒干,直到其重量保持恒定。为最大程度的反映实际情况,采自沙丘不同地貌部位的结皮样品置于相应的位置。考虑所选样地为半固定沙丘,且项目区风季已过,实验过程中对置于丘顶的样品进行了防沙处理,即把微型蒸渗计置于直径10 cm、高5 cm的PVC环中,以防止流沙对实验结果造成不利影响。观测期间如遇降雨事件,雨后把蒸渗计晒之恒重之后继续用于凝结水的观测。实验观测时间为2010年8月13日—8月29日(17、20和21日下雨)和9月13—9月23日(17—19日下雨,20日未观测)。观测方法为称重法,即19:00和次日7:00采用精度为0.01 g的电子天平对样品进行称重,蒸渗计的重量之差视为夜间形成的凝结水量,每次称重前擦净蒸渗计表面和底部沙粒以保证实验精度。凝结水形成过程的观测自19:00开始每2 h进行1次,蒸发过程的观测自7:00开始每30 min进行1次。由于蒸渗计的底部被防水胶带封住,因此把蒸渗计重量变化视为空气中水汽和蒸渗计中土壤水分在上界面交换的结果。蒸渗计重量增加则视为形成了凝结水,重量减少了则视为水分蒸发。最后以质量表示的凝结水量转换成以高度表示的凝结水量,具体转换公式是:
H=10m/ρπr2
式中,H为凝结水量(mm);m为样品重量变化(g);r为蒸渗计的半径(cm);ρ为水的密度(g/cm3)。观测期间的大气温度、地表温度和空气相对湿度等气象数据来自实验站附近的小型自动气象站(DELTA-T, Devices Inc., Cambridge, UK)。不同类型地表凝结水量的差异采用单因素方差分析法(ANOVA)。
2 结果与分析
2.1 凝结水量
观测期间共有20次实验记录。除降雨天气外,每天都能观测到水分凝结现象,但凝结水的日变化较大(图1)。不同类型地表日凝结水量有差异,基本呈现出裸沙lt;浅灰色藻类结皮lt;黑褐色藻类结皮lt;苔藓结皮的趋势,生物结皮表面的日凝结水量大于裸沙,随着生物结皮的发育,日凝结水量呈增加的趋势。

图1 观测期间不同类型地表凝结水量的日变化Fig.1 Daily dew amount of different soil surface types during experimental periods图中误差棒代表标准误 (n=4)
不同类型地表凝结水总量有差异(表1),凝结水总量依次是(1.998±0.075),(2.326±0.083),(2.790±0.058)和(3.416±0.068) mm,不同类型地表凝结水总量之间差异显著(Plt;0.05),生物结皮表面的凝结水总量显著大于裸沙,生物结皮表面的凝结水总量随结皮的发育呈增加的趋势,不同发育阶段生物结皮表面的凝结水量之间差异显著(Plt;0.05)。不同类型地表日凝结水量变率较大,裸沙日凝结水量介于0.025—0.232 mm之间、浅灰色藻类结皮日凝结水量介于0.069—0.276 mm之间、灰褐色藻类结皮日凝结水量介于0.083—0.301 mm之间,苔藓结皮日凝结水量介于0.071—0.322 mm之间,凝结水最大值和最小值之间相差数倍。不同类型地表日平均凝结水量具有差异,日平均凝结水量依次是(0.100±0.003),(0.116±0.004),(0.140±0.002)和(0.171±0.003) mm,不同类型地表日平均凝结水量之间差异极显著(Plt;0.01)。
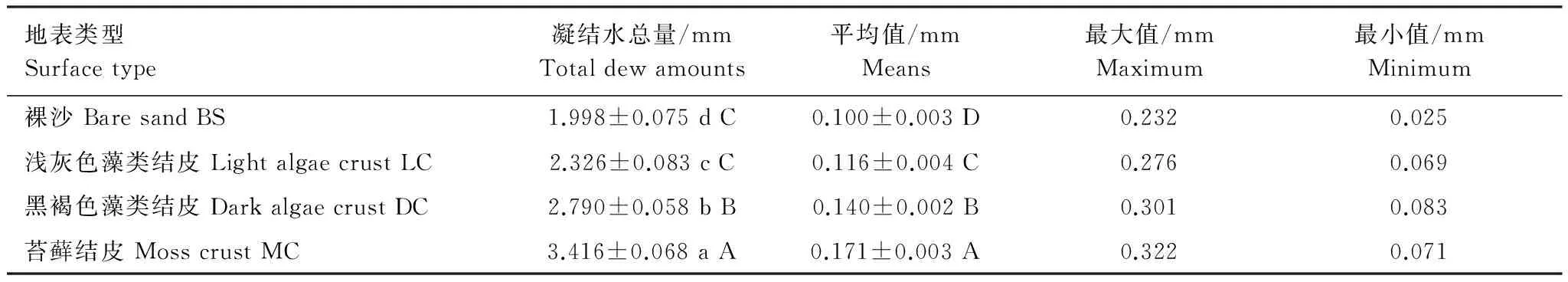
表1 不同类型地表凝结水量分析
不同的小写字母表示0.05水平上的显著性差异,不同的大写字母表示0.01水平上的显著性差异,表中数字为平均值±标准误(n=4)
2.2 凝结水形成和蒸发过程
为探明凝结水的形成过程,对不同类型地表的凝结水量每2 h观测1次,所选日期为8月16日和9月21日晚。观测期间夜间晴朗,风速较低,能够代表观测期间大多数夜间的情况。不同类型地表凝结水形成过程的观测结果显示,凝结水自19:00开始形成,19:00—23:00之间形成较明显,23:00—凌晨1:00之间凝结水形成不明显,裸沙和浅灰色藻类结皮表面尤其不明显,凌晨1:00开始凝结水继续形成,5:00—7:00之间凝结水虽然继续形成,但形成速度放缓(图2)。
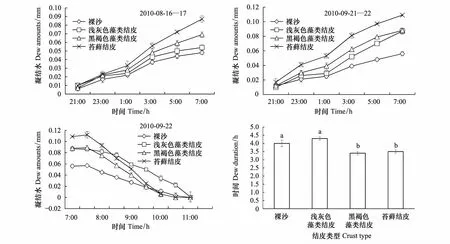
图2 不同类型地表凝结水形成和蒸发过程以及凝结水保持时间Fig.2 Measured cumulative dew deposition, evaporation and dew duration on different soil surface types 图中误差棒为标准误(n=4);不同的小写字母表示差异显著(Plt;0.05); BC: 裸沙; LC: 浅灰色藻类结皮;DC: 黑褐色藻类结皮;MC: 苔藓结皮
为了解凝结水的蒸发过程,自7:00开始每隔30 min观测1次不同类型地表的凝结量,所选择的日期为2010年9月22日。观测期间天气晴朗,风速很小,能够代表晴朗天气结皮表面的凝结水蒸发特征。7:00—7:30之间,除浅灰色藻类结皮外,黑褐色藻类结皮和苔藓结皮表面继续形成少量的凝结水,苔藓结皮表面尤其较明显。7:30开始,夜间形成的凝结水开始蒸发,10:30—11:00之间不同类型地表的凝结水蒸发结束(图2)。凝结水在不同类型地表中的保持时间有差异(图2)。凝结水在裸沙、浅灰色藻类结皮、黑褐色藻类结皮和苔藓结皮中的保持时间分别是(4.0±0.2),(4.3±0.1),(3.4±0.1)和(3.5±0.1) h,凝结水在裸沙和浅灰色藻类结皮中的保持时间显著大于在黑褐色藻类结皮和苔藓结皮中的保持时间(Plt;0.05)。
2.3 凝结水与气象因子之间的关系
凝结水的形成与空气相对湿度、地表温度、空气温度和空气地表温度差之间的关系密切,不同类型地表凝结水量与气象因子之间的关系如图3所示。凝结水与空气相对湿度之间关系密切,夜间空气相对湿度上升,不同类型地表的凝结水量随之增加,到翌日7:00—7:30时达最高;当太阳升起后,空气相对湿度开始减少,地表的凝结水也开始蒸发(图3)。当地表温度达到露点温度时表层土壤、空气中的水汽和土壤孔隙中的水汽变成液态水。随着地表温度的减少,凝结水开始形成,温度达最低时地表所形成的凝结水量最高;当太阳升起后地表温度开始上升,凝结水开始蒸发(图3)。空气温度对凝结水的影响较明显,随着空气温度的降低,地表凝结水呈增加的趋势,空气温度达最低时地表形成的凝结水量达到较高的水平(图3)。当空气温度高于地表温度时,水汽随温度梯度由空气向地表移动,有利于凝结水的形成,反之则不易形成凝结水。观测初始,空气温度大于地表温度,水分向地表移动,开始出现水分凝结现象。随着空气温度的进一步降低,地表温度逐渐开始大于空气温度,此时结皮表面水分凝结现象不明显或出现少量水分蒸发的现象。随着空气温度的进一步降低,地表温度虽然大于空气温度,继续观测到了水分凝结现象,这可能是此时的空气温度低于露点温度,从而继续发生了水分凝结现象,直到太阳升起后随着空气和地表温度的增加,凝结水开始蒸发(图3)。上述结果表明凝结水的形成与大气温度、地表温度、空气相对湿度和空气与地表温度差之间的关系较密切,但凝结水的形成过程不与某一个气象因子呈简单的线性关系。
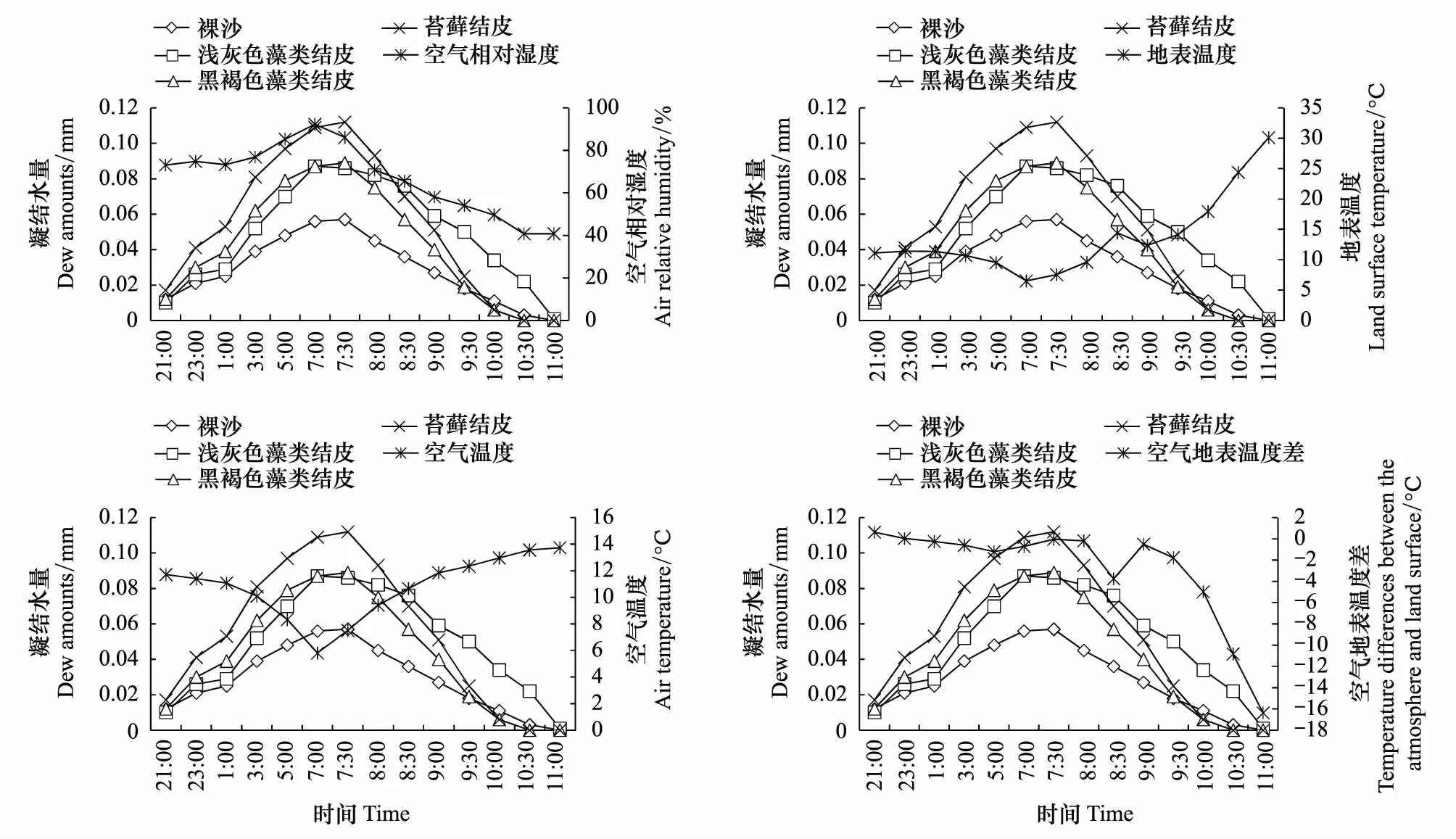
图3 凝结水量与气象因子之间的相关性Fig.3 Correlations between dew amounts and meteorological factorsBS:裸沙,LC:浅灰色藻类结皮,DC:黑褐色藻类结皮,MC:苔藓结皮; RH: 空气相对湿度; AT: 空气温度; TD: 空气地表温差
3 讨论
凝结水的来源主要包括近地面空气中的水汽、土壤中的水汽和植物周围呼吸或蒸腾出的水汽[7]。植被稀疏的干旱区,近地面空气中的水汽是凝结水的主要来源[13]。当地表温度低于空气近地表温度时便可形成凝结水,温差较大的季节尤其容易形成。本文通过不同类型地表凝结水形成和蒸发特征的观测,分析讨论了影响生物结皮表面凝结水形成和蒸发的因素,比较了我国不同气候区生物结皮对凝结水形成的影响,阐明了凝结水在荒漠生态系统中所发挥的重要生态学和水文学意义。
3.1 裸沙对凝结水形成和蒸发的影响
研究区裸沙机械组成以细沙为主,中沙次之[20],其机械组成中无粉粒和黏粒等细颗粒物,而结皮层含有丰富的粉粒和黏粒等细颗粒物。裸沙生物组分与生物结皮相比较单一,主要是一些耐贫瘠的微生物。裸沙表面的凝结水形成和蒸发过程与其生物和非生物组分密切相关。裸沙较粗糙的质地结构可能使裸沙表面对温度变化更加的敏感,从而影响了裸沙对凝结水的捕获能力,造成裸沙表面的凝结水量小于生物结皮。在水分蒸发过程中,裸沙与生物结皮相比表面积较小,相对较小的表面积一定程度上减少了水分蒸发。此外,裸沙机械组成中不含粉粒和黏粒等细颗粒物,裸沙表面补水的毛管作用力很小,较小的毛管作用力可能是裸沙表面水分蒸发速度较慢的原因之一。
3.2 藻类结皮对凝结水形成和蒸发的影响
藻类结皮的生物和非生物成分明显与裸沙不同,藻类结皮对凝结水形成和蒸发的影响与其生物和非生物成分密切相关。构成藻类结皮的生物成分主要以藻类和微生物为主,机械组成中开始出现粉粒和黏粒等细颗粒物。首先,组成藻类结皮的多数藻都是亲水性的,而且有些藻类可以移动,在干旱缺水时处于结皮层的底部,环境适当时能移到结皮表面部位迅速吸收水分,以致吸收后的体积可达原来体积的4—5倍之多[21]。另一方面,微生物和藻体分泌的胞外多糖等物质在凝结水捕获方面发挥着积极的作用,结皮中微小生物体所分泌的胞外多糖等物质吸收水分,提高了藻类结皮对凝结水的捕获能力[22- 23]。不同类型藻类结皮对凝结水的捕获能力也有所不同。黑褐色藻类结皮表面积与浅灰色藻类结皮相比较粗糙,意味着更大的表面积,黑褐色藻类结皮较大的表面积在凝结水捕获方面具有更大的优势,使黑褐色藻类结皮表面的凝结水量大于浅灰色藻类结皮。在水分蒸发过程中,吸水膨胀后的藻体可将堵塞40%之多的结皮层孔隙,一定程度上阻碍了水分从结皮表面蒸发[24],降低了结皮表面水分蒸发速率,使水分在浅灰色藻类结皮中的保持时间较长。黑褐色藻类结皮表面的凝结水蒸发速度较前者快,这可能是黑褐色藻类结皮表面颜色较深的原因,当太阳升起后黑褐色藻类结皮表面能吸收更多的太阳能,进而加快了黑褐色藻类结皮表面的水分蒸发速度[19]。
3.3 苔藓结皮对凝结水形成和蒸发的影响
苔藓结皮的生物组分主要以苔藓植物为主,在种类组成和细颗粒物含量等方面明显与其他类型生物结皮不同。苔藓结皮表面的凝结水总量和日平均凝结水量均大于其他类型的生物结皮,可能是由于苔藓植物体在干旱缺水时皱缩,进入休眠状态,当遇到水分(即使是很少量的露水或雾)之后苔藓植物体能够恢复其生理学活性,恢复生物学活性的苔藓植物体露在结皮表面,与其它类型的生物结皮相比有更多的表面积,较大的表面积在凝结水捕获方面具有更大的优势。另一方面,夜间苔藓结皮覆盖区温度下降较快,当温度低于近地表气温时空气中的水汽开始向地表凝结,苔藓结皮覆盖区较明显的昼夜温差使苔藓结皮表面更容易形成凝结水。白天苔藓结皮表面增温较快,高温促使地表凝结水的快速蒸发与近地表相对湿度的下降,造成了苔藓结皮表面凝结水量的快速下降。这与古尔班通古特沙漠和腾格里沙漠沙坡头地区的研究结果基本一致[12,14]。进一步观测中发现,太阳升出后较短的时间之内凝结水继续形成,发育良好的苔藓结皮表面尤其比较明显,这与张静等在古尔班通古特沙漠地区生物结皮对凝结水影响的研究结果类似[12],这可能是虽然太阳已升出,但地表和空气温度仍然较低,结皮表面继续形成了少量的凝结水。在水分蒸发方面,白天苔藓结皮对空气温度变化较敏感是苔藓结皮表面水分蒸发速率较快的主要原因。此外,发育良好的苔藓结皮中细颗粒物含量较高,较大的毛管作用力使水分不断向苔藓结皮表面移动,进而影响了苔藓结皮表面的水分蒸发速率[25]。再者,多数苔藓植物体遇水之后能够恢复其生物生理学活性,个别苔藓植物体能够在遇水后的短短30 min之内恢复其正常生理活性[26],而苔藓植物体恢复其生物生理活性所需的水分也可能是苔藓结皮表面水分快速减少的一个原因。
3.4 我国不同气候区生物结皮对凝结水影响的比较
我国温带荒漠地区分布着大面积沙漠和沙地,涵盖了从极端干旱到半湿润的不同气候区。目前在古尔班通古特沙漠和腾格里沙漠沙坡头地区已有相关的研究报道。张静等在古尔班通古特沙漠地区生物结皮对凝结水影响的研究结果表明,不同类型生物结皮表面的日平均凝结水量随生物结皮的发育呈增加的趋势[12]。在年均降水量不到200 mm的腾格里沙漠沙坡头地区,不同类型生物结皮的凝结水量表现出与前者类似的规律,生物结皮的日平均凝结水量较前者大或相当[14]。在年均降水量接近400 mm的毛乌素沙地南缘沙区,生物结皮表面的凝结水随生物结皮的发育呈增加的趋势,生物结皮的日平均凝结水量较前两个地区有所增加,尽管不同气候区生物结皮对凝结水影响的研究在观测时间、时段上尚未统一,但日平均凝结水量基本在降水梯度(从极端干旱、干旱到半干旱)上呈增加的趋势。在水分蒸发方面,凝结水在发育早期生物结皮中的保持时间较长,在苔藓和黑褐色藻类结皮中的保持时间较短,这与张静等在古尔班通古特沙漠地区生物结皮对凝结水影响的实验研究结果较类似[12],但凝结水在结皮中保持时间较前者短。腾格里沙漠沙坡头地区的研究结果表明,在小降雨事件中生物结皮的发育加快了水分蒸发过程,而较大降雨事件中生物结皮的发育一定程度上抑制水分蒸发过程,起到保护表层水分的作用[27],如果把夜间形成的凝结水视为很小的降雨事件的话,本文的研究结果与沙坡头地区的研究结果基本吻合。从上述比较可知,不同气候区生物结皮对凝结水形成和蒸发的影响有所差异,是不同气候区生物结皮和气象因素共同作用的结果,因此后续的研究应加强凝结水形成过程的实时观测。
3.5 生物结皮表面凝结水的生态学意义
对水分匮乏的沙区环境来说,凝结水的重要作用是不容忽视的。以色列内盖夫沙漠地区凝结水的观测结果表明,出现水分凝结现象的天数能达到观测天数的50%[28]。生物结皮作为干旱、半干旱沙区重要地表景观之一,其分布面积几乎占到该地区生物覆盖率的40%[19],生物结皮表面形成的凝结水对沙地生态系统产生的重要影响是不容忽视的。首先,生物结皮表面的凝结水在一定程度上减少了沙地生态系统水分蒸发支出的“赤字”,缓解荒漠地区微小生物体的缺水状态。次之,凝结水成为荒漠地区微小生物体或隐花植物的珍贵水资源,能够使其保持生物学活性,进而影响荒漠生态系统的功能和过程。再者,发育良好的苔藓结皮表面的凝结水量最高,蒸发速度也较快,因此生物结皮的进一步发育一定程度上加快了沙土表层水分循环过程,进而对沙地生态系统产生影响。
[1] Wang X P, Li X R, Kang E S, Zhang J G, Zhou H Y, Yang S X, Lei Z D. The infiltration and redistribution of precipitation in revegetated sand dunes in the Tengger Desert, Shapotou, China. Acta Ecologica Sinica, 2003, 23(6): 1234- 1241.
[2] Zhang X Y, Li X Y, Wang W, Ma Y J. Experimental observation analysis on dew formation in southern Mu us sandy land. Arid Meteorology, 2008, 26(3): 8- 13.
[3] Li X R, Zhang Y M, Zhao Y G. A study of biological soil crusts: recent development, trend and prospect. Advances in Earth Science, 2009, 24(1): 11- 24.
[4] Zhang X Y. Study on Dew Formation Mechanism in Southern Mu Us Sandy Land [D]. Beijing: Beijing Normal University, 2009.
[5] Boast C W, Roberts T M. A “micro-lysimeter” method for determining evaporation from bare soil: description and laboratory evaluation. Soil Science Society of America Journal, 1982, 46(4): 689- 696.
[6] Kidron G J. A simple weighing method for dew and fog measurements. Weather, 1998, 53(12): 428- 433.
[7] Garratt J R, Segal M. On the contribution of atmospheric moisture to dew formation. Boundary Layer Meteorology, 1988, 45(3): 209- 236.
[8] Kidron G J. Altitude dependent dew and fog in the Negev Desert, Israel. Agricultural and Forest Meteorology, 1999, 96(1/3): 1- 8.
[9] Kidron G J. Analysis of dew precipitation in three habitats within a small arid drainage basin, Negev Highlands, Israel. Atmospheric Research, 2000, 55(3/4): 257- 270.
[10] Kidron G J. Angle and aspect dependent dew and fog precipitation in the Negev desert. Journal of Hydrology, 2005, 301(1/4): 66- 74.
[11] Zangvil A. Six years of dew observations in the Negev Desert, Israel. Journal of Arid Environments, 1996, 32(4): 361- 372.
[12] Zhang J, Zhang Y M, Zhou X B, Zhang B C, Wei M L. The influence of biological soil crusts on dew deposition and characteristics of soil surface in Gurbantunggut Desert. Acta Ecologica Sinica, 2009, 29(12): 6600- 6608.
[13] Feng J Z, Liu L C, Xiao H L, Zeng W B, Liu C M. Dynamic measurement and theoretical calculation on water absorption and condensation of sandy soil in Shapotou region. Journal of Desert Research, 1998, 18(1): 12- 17.
[14] Liu L C, Li S Z, Duan Z H, Wang T, Zhang Z S, Li X R. Effects of microbiotic crusts on dew deposition in the restored vegetation area at Shapotou, northwest China. Journal of Hydrology, 2006, 328(1/2): 331- 337.
[15] Pan Y X, Wang X P, Zhang Y F. Dew formation characteristics in a revegetation-stabilized desert ecosystem in Shapotou area, Northern China. Journal of Hydrology, 2010, 387(3/4): 265- 272.
[16] Lan S B, Hu C X, Rao B Q, Wu L, Zhang D L, Liu Y D. Non-rainfall water sources in the topsoil and their changes during formation of man-made algal crusts at the eastern edge of Qubqi Desert, Inner Mongolia. Science China (Life Science), 2010, 53(9): 1135- 1141.
[17] Jiang J, Wang K F, Zhang W J. A study on the coagulation water in the sandy soil and its role in water balance. Arid Zone Research, 1993, 10(2): 1- 9.
[18] Liu X P, He Y H, Zhao X Y, Li Y S, Li Y Q, Li Y Q, Li S M. Soil condensation water in different habitats in Horqin sandy land: An experimental study. Chinese Journal of Applied Ecology, 2009, 20(8): 1918- 1924.
[19] Belnap J. The potential roles of biological soil crusts in dryland hydrologic cycles. Hydrological Processes, 2006, 20(15): 3159- 3178.
[20] Wu Y S, Ha S, Li S Q, Liu H Q, Jia Z J. Development characteristics of biological soil crusts on sand dune in southern Mu Us sandy lands. Journal of Soil and Water Conservation, 2010, 24(5): 258- 261.
[21] Booth W E. Algae as pioneers in plant succession and their importance in erosion control. Ecology, 1941, 22(1): 38- 46.
[22] Zhong Z P, Shi D J. Differentiation of hormogonia from filamentous Cyanobacteria and their regulation mechanisms. Chinese Bulletin of Botany, 2000, 17(3): 204- 210.
[23] Hu C X, Liu Y D. Soil algal biomass and its influential factors in desert soil crusts. Acta Ecologica Sinica, 2003, 23(2): 284- 291.
[24] Verrecchia E, Yair A, Kidron G J, Verrecchia K. Physical properties of the psammophile cryptogamic crust and their consequences to the water regime of sandy soils, North-western Negev Desert, Israel. Journal of Arid Environments, 1995, 29(4): 427- 437.
[25] Gill B S, Jalota S K. Evaporation from soil in relation to residue rate, mixing depth, soil texture and evaporativity. Soil Technology, 1996, 8(4): 293- 301.
[26] Tuba Z, Csintalan Z, Proctor M C F. Photosynthetic responses of a moss,Tortularuralis, ssp.ruralis, and the lichensCladoniaconvolutaandC.furcatato water deficit and short periods of desiccation, and their ecophysiological significance: A baseline study at present day CO2concentration. New Phytologist, 1996, 133(2): 353- 361.
[27] Zhang Z S, Liu L C, Li X R, Zhang J G, He M Z, Tan H J. Evaporation properties of a revegetated area of the Tengger Desert, North China. Journal of Arid Environments, 2008, 72(6): 964- 973.
[28] Kidron G J, Herrnstadt I, Barzilay E. The role of dew as a moisture source for sand microbiotic crusts in the Negev Desert, Israel. Journal of Arid Environments, 2002, 52(4): 517- 533.
参考文献:
[1] 王新平, 李新荣, 康尔泗, 张景光, 周海燕, 杨诗秀, 雷志栋. 腾格里沙漠东南缘人工植被区降水入渗与再分配规律研究. 生态学报, 2003, 23(6): 1234- 1241.
[2] 张晓影, 李小雁, 王卫, 马育军. 毛乌素沙地南缘凝结水观测实验分析. 干旱气象, 2008, 26(3): 8- 13.
[3] 李新荣, 张元明, 赵允格. 生物土壤结皮研究:进展、前沿与展望. 地球科学进展, 2009, 24(1): 11- 24.
[4] 张晓影. 毛乌素沙地南缘凝结水形成机理研究 [D]. 北京: 北京师范大学, 2009.
[12] 张静, 张元明, 周晓兵, 张丙昌, 魏美丽. 生物结皮影响下沙漠土壤表面凝结水的形成与变化特征. 生态学报, 2009, 29(12): 6600- 6608.
[13] 冯金朝, 刘立超, 肖洪浪, 曾文炳, 刘成敏. 沙坡头地区土壤水分吸湿凝结的动态观测与理论计算. 中国沙漠, 1998, 18(1): 12- 17.
[17] 蒋瑾, 王康富, 张维静. 沙地凝结水及在水分平衡中作用的研究. 干旱区研究, 1993, 10(2): 1- 9.
[18] 刘新平, 何玉惠, 赵学勇, 李玉霖, 李玉强, 李衍青, 李世民. 科尔沁沙地不同生境土壤凝结水的试验研究. 应用生态学报, 2009, 20(8): 1918- 1924.
[20] 吴永胜, 哈斯, 李双权, 刘怀泉, 贾振杰. 毛乌素沙地南缘沙丘生物土壤结皮发育特征. 水土保持学报, 2010, 24(5): 258- 261.
[22] 钟泽璞, 施定基. 丝状体蓝藻藻殖段的分化及其调节机制. 植物学通报, 2000, 17(3): 204- 210.
[23] 胡春香, 刘永定. 土壤藻生物量及其在荒漠结皮的影响因子. 生态学报, 2003, 23(2): 284- 291.
EffectsofbiologicalcrustsondewdepositionandevaporationintheSouthernEdgeoftheMuUsSandyLand,NorthernChina
YIN Ruiping1,WU Yongsheng1,*, ZHANG Xin1, HA Si2, TIAN Xiumin1, LI Zekun1, WANG Jian1, MIAO Henglu1, REN Jie1
1InstituteofWaterResourcesforPastoralArea,MinistryofWaterResources,Hohhot010020,China2CollegeofResourcesScienceandTechnology,BeijingNormalUniversity,Beijing100875,China
Dew is one of the most important moisture sources except rainfall for extremely water limited desert ecosystems and they have a significant effect on the maintenance of ecosystem structure, function and processes. To better understand the influence of biological crusts on dew deposition and evaporation processes in a semiarid sandy ecosystem, dew quantities of different surface types (bare sand, light algae crusts, dark algae crusts and moss crusts) were measured in the southern edge of the Mu Us sandy land in northern China using micro-lysimeters with a diameter of 7 cm and a height of 5 cm. The main conclusions were as follows: (1) There are 20 records in the experimental periods where dew was deposited almost every day except rainy days; (2) The total dew amounts of different soil surface types were (1.998±0.075), (2.326±0.083), (2.790±0.058) and (3.416±0.068) mm for bare sand, light algal crusts, dark algal crusts and moss crusts, respectively. Moss crusts were characterized by having the greatest amount of dew at dawn, whereas bare sand yielded the lowest amount of dew and the total dew amounts of biological crusts were significantly higher than that in bare sand (Plt;0.05). Therefore it can be concluded that the formation of biological crusts benefited dew deposition. Dew quantities of biological soil crusts increased with crust development and the total dew amounts of different types of biological crusts varied significantly (Plt;0.05). Daily average dew amounts of different soil surface types were (0.100±0.003), (0.116±0.004), (0.140±0.002) and (0.171±0.003) mm for the bare sand, light algal crusts, dark algal crusts and moss crusts, respectively. The maximum amount of dew for each type of crust studied was several times greater than the minimum and the daily average dew amounts of different soil surface types were significantly different from each other (Plt;0.01); (3) Results of dew formation processes at 2 hour intervals indicated that dew formation begins at 19:00, inconspicuous dew was deposited from 23:00 to 1:00 which then proceeded from 1:00 to 7:00. From 7:00 to 7:30, continuous dew was deposited on the surface of dark algae crusts and moss crusts even after sunrise but not on light algae crusts. Results of dew evaporation processes at 30 min intervals indicated that dew evaporation begins at 7:30, and ends from 10:30 to 11:00. Dew duration time in bare sand and light algae crusts was significantly higher than that in dark algae crusts and moss crusts (Plt;0.05). It can be concluded that well developed biological crusts may potentially enhance hydrological circulation in the upper sand layer in sandy land. (4) Dew formation processes can be affected by air temperature, land surface temperature, relative air humidity and temperature differences between the atmosphere and land surface. Dew formation processes are not simply linearly related to any meteorological factors. These data further the understanding of dew formation characteristics of biological crusts in semi-arid sandy areas and assist in evaluating the significant ecological roles of dew in semi-arid environments.
Mu Us sandy land; biological crusts; dew; evaporation
国家自然科学基金资助项目(41201015); 中国水利水电科学研究院科研专项资助项目
2013- 04- 03;
2013- 07- 15
*通讯作者Corresponding author.E-mail: yswuwork@126.com
10.5846/stxb201304030594
尹瑞平,吴永胜,张欣,哈斯,田秀民,李泽坤,王健,苗恒录,任杰.毛乌素沙地南缘沙丘生物结皮对凝结水形成和蒸发的影响.生态学报,2013,33(19):6173- 6180.
Yin R P,Wu Y S, Zhang X, Ha S, Tian X M, Li Z K, Wang J, Miao H L, Ren J.Effects of biological crusts on dew deposition and evaporation in the Southern Edge of the Mu Us Sandy Land, Northern China.Acta Ecologica Sinica,2013,33(19):6173- 6180.
猜你喜欢
新疆大学学报(自然科学版)(中英文)(2022年3期)2022-06-04
军事文摘(2021年18期)2021-12-02
润·文摘(2021年11期)2021-11-20
草业学报(2021年6期)2021-06-28
水土保持通报(2020年6期)2020-03-10
中华建设(2019年12期)2019-12-31
儿童故事画报·自然探秘(2017年7期)2018-03-14
新疆大学学报(自然科学版)(中英文)(2018年4期)2018-02-10
中国中西医结合皮肤性病学杂志(2016年4期)2016-07-18
电站辅机(2016年4期)2016-05-17
