
脂肪酶降解几丁质的研究
2013-12-24 06:44尚利明陈小龙
浙江农业科学 2013年7期
尚利明,陈小龙
(浙江工业大学发酵工程研究所,浙江杭州 310014)
几丁质 (Chitin)又名甲壳素,是一种由1 000~3 000个乙酰葡萄糖胺残基通过β-1,4糖苷链相互连接而成的天然氨基多糖高分子聚合物[1]。几丁质来源丰富,且有良好的生物相容、生物降解、无毒和易成膜等特性,是目前世界上唯一含阳离子的可食性动物纤维,已广泛应用于农业、医药和环保等领域。由于几丁质分子量大,具有紧密的晶体结构,分子间有较强的氢键作用,不溶于水和一般的有机溶剂,使其在应用方面受到很大限制。相比之下,几丁寡糖水溶性好,易被分散和吸收,且具有诱导植物抗病性反应[2]、调控植物生长发育[3]、抗菌、抑制肿瘤生长、提高机体免疫机能、促进肠道功能、在宿主体内累积效应弱等优点,可广泛应用于农业、医药和食品等领域。因此,几丁寡糖的研究引起了国内外的重视[4]。
利用高分子量的几丁质制备低分子量的几丁寡糖的方法主要有化学降解和酶法降解2种。传统的化学降解法对反应条件要求高,难以控制,且后处理繁琐,不易得到低聚糖,尤其是在反应过程中会消耗大量的浓酸和浓碱,对环境造成严重污染[5-6]。酶法降解几丁质因其具有条件温和、易于控制、不发生副反应、无污染及产物均一性好等特点而备受关注[7]。专一性水解酶在几丁质水解中效果明显,但由于获取困难且价格昂贵从而限制了其广泛应用。因此,研究者将视线转向来源广、价格低、效果较好的非专一性水解酶[8]。常用的非专一性水解酶主要有纤维素酶[9]、蛋白酶[10-11]、溶菌酶[12]和脂肪酶[13]。与其他水解酶相比,目前对于脂肪酶降解几丁质的研究较少,而脂肪酶生产成本更低。本文利用猪胰脂肪酶水解几丁质,旨在为利用脂肪酶催化几丁质制备几丁寡糖提供理论依据。
1 材料与方法
1.1 脂肪酶与主要试剂
猪胰脂肪酶,北京凯泰新世纪生物技术有限公司;几丁质,浙江金壳生物化学有限公司;3,5-二硝基水杨酸 (分析纯,后同),国药集团化学试剂有限公司;NaOH,上海试四赫维化工有限公司;酒石酸钾钠,上海美兴化工股份有限公司;苯酚,杭州双林化工试剂厂;亚硫酸钠,浙江省永嘉县化工试剂厂;乙酰氨基葡萄糖,阿拉丁。
1.2 胶体几丁质制备
将5 g几丁质溶于88 mL浓盐酸中,磁力搅拌器200 r·min-1搅拌1 h后,4℃下静置24 h,然后加入500 mL去离子水,混匀后于4 000 r·min-1下离心5 min,移去上清液,将沉淀物洗至中性,再用去离子水定容至250 mL,即成2%胶体几丁质,在4℃下保存。
1.3 DNS配制
将3,5-二硝基水杨酸溶液1.575 g和2 mol·L-1NaOH溶液65.2 mL添加到125 mL含有46.25 g酒石酸钾钠的热水溶液中,加入1.25 g结晶苯酚和1.25 g亚硫酸钠,搅拌至溶解,冷却后加蒸馏水定容至250 mL,储存在棕色瓶中备用。
1.4 脂肪酶水解几丁质酶活力测定
取脂肪酶0.03 g,2%胶体几丁质2 mL放入25 mL具塞试管中,40℃水浴反应60 min,然后沸水浴1 min终止酶反应,10 000 r·min-1离心5 min,取1 mL上清液加入2 mL DNS,置沸水浴中反应10 min后迅速冷却,加蒸馏水定容至10 mL,对照加灭活后的脂肪酶,540 nm测D值。根据N-乙酰氨基葡萄糖标准曲线计算出酶活力。
酶活力单位 (U):40℃下每分钟产生相当于1 μmol N-乙酰氨基葡萄糖的还原糖量所需的酶量。
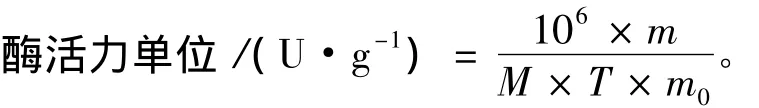
式中:m为生成的还原糖的质量 (g);M为乙酰氨基葡萄糖的分子质量 (221.21 g·mol-1);m0为酶液质量 (g);T为反应时间 (min);106为比例系数。
1.5 反应体系
反应在25 mL锥形瓶中进行,初始反应体系为胶体几丁质溶液2 mL(2%,pH值7.4),猪胰脂肪酶0.03 g。在40℃恒温水浴140 r·min-1摇床中反应1 h。
2 结果与分析
2.1 酶加量对酶促反应的影响
取胶体几丁质溶液2 mL(2%,pH值7.4),分别添加0.01,0.02,0.03,0.04和0.05 g猪胰脂肪酶,40℃下水浴1 h。不同酶加量对酶促反应的影响如图1所示。
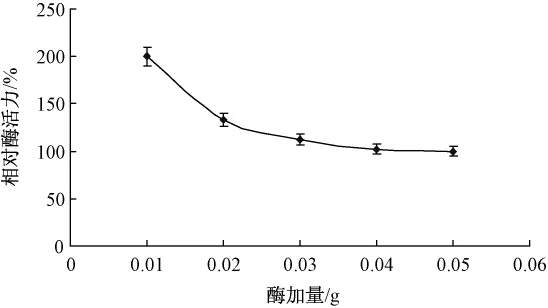
图1 酶加量对酶促反应的影响
随着脂肪酶添加量的增多,反应体系的粘度不断增加,使得酶不能充分溶解于底物溶液中,活性中心不能完全暴露出来与底物充分接触,导致酶促反应速率开始降低。综合考虑最适酶量为0.01 g。
2.2 底物浓度对酶促反应的影响
酶加量0.01 g,分别添加浓度为1%,2%,3%,4%和5%的胶体几丁质溶液2 mL(pH 7.4),40℃下水浴1 h。具体影响如图2所示。
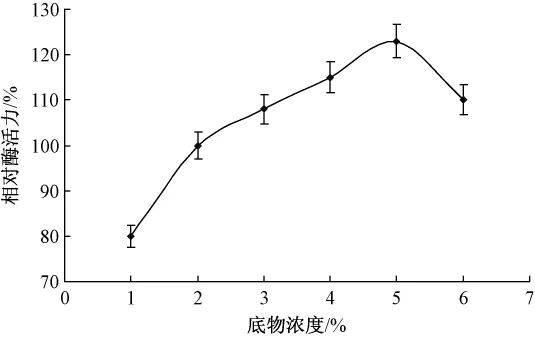
图2 底物浓度对酶促反应的影响
随着底物浓度的增加,酶的活性中心与底物的接触面积逐渐增大,酶促反应速率也随之加快。但当底物浓度超过5%时,开始限制酶促反应的进行,这是因为过高的溶液黏度会大大限制酶分子的自由移动,故最适底物浓度为5%。
2.3 温度对酶促反应的影响
酶量0.01 g,胶体几丁质溶液2 mL(5%,pH值7.4),分别在35℃,40℃,45℃,50℃,55℃,60℃,65℃和70℃条件下水浴1 h。不同温度对酶促反应的影响如图3所示。
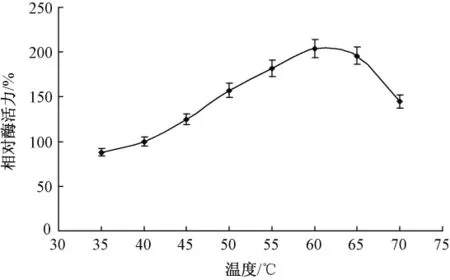
图3 温度对酶促反应的影响
温度的升高使反应物分子获得能量,使一部分原来能量较低分子变成活化分子,增加了活化分子的百分数,使得有效碰撞次数增多,故酶促反应速率加大。当然,由于温度升高,使分子运动速率加快,单位时间内反应物分子碰撞次数增多,这也使酶促反应速率相应加快。但当温度超过60℃后,酶促反应速度开始降低,主要原因是酶本质上是一种蛋白质,遇热易变性。当超过其最适温度后,随着温度的升高,酶促反应速度加快,但同时酶开始变性失活,从而使活性酶的浓度大为降低,因此,酶促反应最适温度为60℃。
2.4 pH对酶促反应的影响
酶量0.01 g,5%胶体几丁质溶液2 mL,溶液pH分别调节为6.0,6.5,7.0,7.5和8.0,60℃下水浴1 h,不同pH对酶促反应的影响如图4所示。
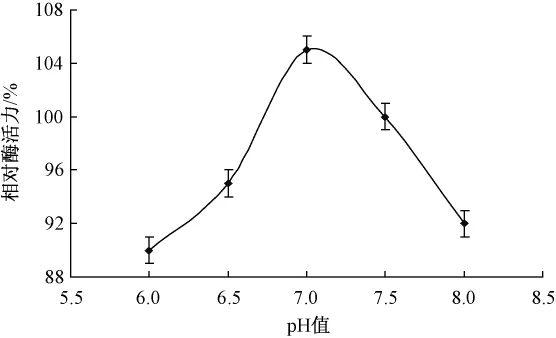
图4 pH对酶促反应的影响
在酸性条件下,随着pH值的升高,酶促反应速度也随之逐渐升高;当pH超过值7.0后,酶促反应速度随着pH值的升高开始降低,但酶活力变化不大,说明在pH值6.0~8.0范围内,脂肪酶活力保持相对稳定,最适值pH为7.0。
pH值对酶促反应速度的影响主要表现在两方面:一是环境过酸、过碱会影响酶蛋白的构象,使酶本身变性失活;二是pH值的改变会影响酶分子侧链上极性基团的解离,改变其带电状态,从而使酶活性中心的构象发生变化。在最适pH值条件时,酶分子活性中心上的有关基团的解离状态最适于与底物结合,表现为酶促反应达到最高。当pH值低于或高于最适pH值时,活性中心上有关基团的解离状态发生改变,酶和底物的结合力降低,因而酶促反应速度降低。同时,pH值也能影响底物分子的解离。底物分子上某些基团只有在一定的解离状态下才适于与酶结合发生反应。pH值的改变偏离了最适条件,则会影响这些基团的解离,使之与酶的结合能力降低,酶促反应速度也随之减慢。因此,pH值的改变会影响酶与底物的结合,也会影响酶促反应中间产物的生成,从而影响酶促反应速度。
2.5 反应时间对转化率的影响
酶加量0.01 g,胶体几丁质溶液2 mL(5%,pH值7.0),60℃下反应不同时间,结果如图5所示。随着时间的延长,酶促反应速率逐渐降低,转化率也在增高,当超过30 h后,酶促反应趋于停滞,转化率也趋于稳定。原因可能是酶在长时间高温作用下,稳定性变差,活性逐渐减弱,酶活性部位与底物的结合能力变差,导致酶促反应趋于停滞。反应30 h后,几丁质转化率为6%,与初始条件下相比,转化率提高了2倍。
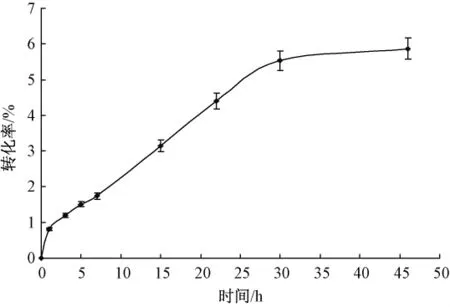
图5 反应时间对转化率的影响
3 小结与讨论
通过对猪胰脂肪酶水解几丁质的条件研究认为,在反应体系为2 mL,酶加量0.01 g,胶体几丁质浓度5%,pH值7.0,60℃条件下反应30 h后,几丁质转化率达到最高 (6%)。为了进一步提高酶法水解几丁质的转化效率,可考虑采用流加廉价脂肪酶的方法。同时,作为非专一性水解酶,猪胰脂肪酶的活性中心不能与几丁质分子完全契合,这也是导致其转化率不高的原因之一。因此,可考虑对脂肪酶的活性中心进行改造。
同化学法降解几丁质相比,利用脂肪酶降解几丁质具有对环境无污染、反应条件温和、易于操作、产物易于分离纯化、脂肪酶成本低廉且可以重复利用等多重优点。因此,随着对环境保护越来越重视,利用脂肪酶制备几丁寡糖将具有良好的应用前景。
[1] 张灿,黄德智,李丰硕,等.海洋产几丁质酶菌株的筛选及发酵优化 [J].吉林农业大学学报,2012,34(2):141-146.
[2] 唐勇军,彭丽莎,邹俊,等.几丁质酶产生菌发酵液对水稻生长及防御酶活性的影响 [J].热带作物学报,2011,32(3):393-397.
[3] 徐翠莲,杨楠,杨国玉,等.壳寡糖膦酸酯对烟草花叶病毒抗性及其机理的初步研究 [J].中国烟草学报,2012,18(1):58-1901.
[4] 乔兴忠,李永娴,汤熙翔.发酵法产酶生产几丁寡糖的工艺研究[J].厦门大学学报,2010,49(2):251-255.
[5] Allan G G,Peyron M.Molecular weight manipulation of chitosan I:kinetics of depolymerization by nitrous acid[J].Carbohydrate Research,1995,277(2):257-272.
[6] Allan G G,Peyron M.Molecular weight manipulation of chitosan II:prediction and control of extent of depolymerization by nitrous acid [J].Carbohydrate Research,1995,277(2):273-282.
[7] Jeon Y J, Kim S K. Continuous production chitin oligosaccharides using a dual reactor system [J].Process Biochemistry,2000,35(1):623-632.
[8] Jeon Y J,Shahidi F,Kim S K.Preparation of chitin and chitosan oligomersand theirapplicationsin physiological functional foods [J].Food Reviews International,2000,16(2):159-176.
[9] Qin C Q,Zhou B,Zeng L T,et al.The physicochemical properties and antitumor activity of cellulose-treated chitosan[J].Food Chemistry,2004,84:107-115.
[10] Kumar A B,Gowda L R,Tharanathan R N.Non-specific depolymerization of chitosan by pronase and charac terization of the resultant products [J].European Journal of Biochemistry,2004,271(4):713-723.
[11] Kumar A B,Varadaraj M C,Lalitha R G,et al.Low molecular weight chitosans:preparation with the aid of papain and characterization [J].Biochimica et Biophysica Acta,2004,1670(2):137-146.
[12] Masaki A,Fukamizo T.Lysozyme catalyzed reaction of chitin oligosaccharides [J]. Journal of Biochemistry, 1981(90):527.
[13] Xia W S,Muzzarelli R A A.Depolymerization of chitosan and substuted chitosans with the aid of lipase[C].Proceedings of the second international.Wuxi:Conference on Food Science and Technology,1994:147-157.
猜你喜欢
动物营养学报(2022年8期)2022-09-01
湖南农业科学(2022年3期)2022-05-18
科技视界(2021年32期)2021-12-08
食品工业科技(2021年22期)2021-11-14
动漫界·幼教365(中班)(2020年7期)2020-07-14
小天使·三年级语数英综合(2019年3期)2019-05-08
中成药(2017年8期)2017-11-22
小星星·阅读100分(高年级)(2017年9期)2017-09-22
中国酿造(2017年8期)2017-09-03
中国当代医药(2015年31期)2015-03-01
