
产角质酶酵母菌的发现与鉴定及产酶条件的优化
2014-01-14 04:39冉琴琴张效宁张文坤张学俊张萌萌
生物技术通报 2014年5期
冉琴琴 张效宁 张文坤 张学俊 张萌萌
(1.贵州大学 贵州省发酵工程与生物制药重点实验室,贵阳 550025;2.贵州大学酿酒与食品工程学院,贵阳 550025)
从植物中提取天然活性成分化合物,即从植物的组织结构中溶出这些化合物,并使之与固体组织分离。传统的天然产物提取方法有碱煮法、酸煮法以及水煮或醇煮法,但这些方法都涉及到高温蒸煮,对药效好的高活性药物可能造成活性丧失;特别是酸性或碱性条件下,还有可能改变化合物的药效结构,如苷键、酯键的水解等。环境污染明显也是传统方法的一大弊端。酶降解提取植物中天然药物成分方法是一种温和的提取方法,因为组织结构的破坏,多种组分可一并溶出,显著地减少提取液的用量,降低污染,降低单一组分的提取成本;多种天然产物的同时溶出,可提高分离纯化的效率,降低纯化的成本;此外,酶法提取最大的特点就是反应条件温和,酶作为一种生物催化剂,具有高效性、底物专一性等特点,在反应中不会发生副反应,保证了植物中有效成分的结构性质不被破坏,有利于后续生产。杜仲(Eucommia ulmoides Oliv)是中国特有的名贵经济树种[1,2],杜仲叶与杜仲皮中的化学成分十分相似,有着同样的药理作用,而且杜仲叶中还含有多种杜仲皮中所没有的成分,杜仲属于落叶乔木,其叶子是可再生的,资源丰富[3],相比于杜仲皮更具有开发价值。杜仲叶中的杜仲胶、绿原酸、桃叶珊瑚苷、杜仲黄酮、木脂素等多种成分的同时提取是对自然资源的充分利用和保护,最终使整个生产获得较高的经济效益和社会效益。
但是,用纤维素酶、果胶酶、蛋白酶、淀粉酶等生物酶降解叶片植物组织和细胞壁[4,5]时,在叶片组织的最外层有一层保护植物组织不受外界生物浸蚀的角质保护层,阻挡了各种酶降解植物组织的作用。当致病菌入侵植物时,必须分泌出角质酶降解角质层,破坏角质层后方可侵入到植物内部。同样,欲用酶水解植物叶片,必须先水解表面的角质层。
目前的研究已证明,角质酶是降解角质层的最主要的功能酶。而在自然界中,角质酶的来源十分有限,仅有微生物和花粉[6]两个途径。产角质酶的微生物也很少,主要为植物的病原真菌,至今尚无有关于产角质酶酵母菌的报道。相对于病原真菌的培养来说,酵母菌的意义更为重大,因为酵母菌生长周期比其他病原真菌短,且对环境来说不具有病原真菌的危害性,是安全的。
角质酶(EC3.1.1.74)是一种多功能酶[7],具有属于α/β 水解酶折叠的共同结构的蛋白质,可以降解角质产生大量的脂肪酸[8],也可以水解甘油三酯等化合物[8,9]。角质酶是丝氨酸水解酶家族中较小的成员。因为角质酶能够水解短链或长链酯类,还能够催化酸与醇的酯化、脂肪酸盐与醇的转酯化等反应,在食品工业、制药工业、化工工业及印染工业等许多领域得到广泛应用[10],因此,对产角质酶菌株的研究意义十分重大。本研究从病变的杜仲叶分离出一株能高产角质酶的菌种,鉴定分析证实该菌株是胶红酵母,并对该菌株发酵产角质酶的条件进行优化,旨在为角质酶的大规模生产提供参考。
1 材料与方法
1.1 材料
1.1.1 试验材料 对硝基苯丁酸酯(p-nitrophenyl butyrate,PNB)购自Sigma 公司;杜仲叶购买于贵州省遵义市遵义县新舟镇金仲村;马铃薯购于附近市场;NaNO3、K2HPO4、MgSO4、KCl、FeSO4·7H2O、琼脂、葡萄糖、草酸、草酸铵、Na2HPO4、KH2PO4等试剂均为国产分析纯。
1.1.2 培养基 基础培养基(PDA):马铃薯200 g,葡萄糖20 g,琼脂20 g(液体培养基中不加琼脂),蒸馏水 1 000 mL。富集培养基[11]:马铃薯20%,葡萄糖 2%,KH2PO4·3H2O 0.5%,硫胺素(VB1)0.008%。筛选培养基:NaNO33.0 g;K2HPO41.0 g;KCl 0.5 g;FeSO4·7H2O 0.01 g,琼 脂17 g,蒸 馏 水1 000 mL,角质0.2%。发酵基础培养基[12]:葡萄糖 1.0%,NaNO30.6 g/L,K2HPO40.6 g/L,MgSO40.2 g/L,KCl 0.2 g/L,FeSO4·7H2O 0.1 g/L,角质0.1%。
以上培养基除需要特别要求的pH 用HCl 或NaOH 调节外,其余的均为自然pH,于121℃灭菌20 min,冷却至室温备用。
1.2 方法
1.2.1 菌种分离 挑选病变的杜仲叶揉碎,取少量加入富集培养基中在30℃恒温振荡培养箱中培养2 d 后,取其培养液用无菌水按一定倍数稀释涂布,接种于筛选培养基的平板上培养24 h 后进行观察识别,将识别的不同菌株分别在不同的PDA 平板上划线培养,连续分离操作3 次,对菌株反复进行分离、纯化。
1.2.2 角质酶酶活的测定 酶活测定[13]:以对硝基苯丁酸酯(PNB)为底物测定酶的活力。反应体系为1.8 mL,包括200 μL 的粗酶液、200 μL 0.4%的Tritonx-100、1 380 μL 50 mmol/L 磷 酸 盐 缓 冲 液(pH7.0)和20 μL 1.76%的PNB 溶液。于37℃反应10 min 后,在波长405 nm 处测定吸光度值。在反应条件下,将每毫升酶液每分钟产生1 μg 的对硝基苯酚定义为1 个酶活单位,即1 U/mL。
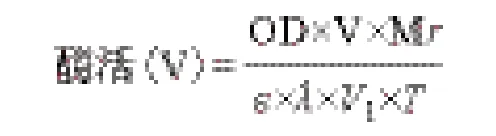
其中,OD:吸光值;V:反应体系的体积,1.8 mL;Mr:产物对硝基苯酚的分子量,139.11 g/mol;ε:摩尔消光系数,6 830 L/(mol·cm);λ:比色皿的光程,0.5 cm;V1:酶液体积,200 μL;T:反应时间10 min。
1.2.3 培养方法 种子培养:将储备于低温冰箱中的菌种接入斜面种子培养基上,在30℃培养48 h,然后接种于装有100 mL 液体PDA 的250 mL 锥形瓶中,30℃、160 r/min 摇床培养36 h。
发酵培养:将菌种活化后接种到装有100 mL 基础培养的250 mL 锥形瓶里,置于30℃恒温摇床中、160 r/min 环境下培养。6 d 后将发酵液用12 层纱布过滤,滤液再于5 500 r/min 4℃下离心30 min,上清即为粗酶液,用于酶活测定。
1.2.4 菌种鉴定方法 首先按SK8229 真菌基因组DNA 抽提试剂盒的要求提取DNA,并以提取得到的真菌DNA 作为模板,再使真菌通用引物扩增18S rDNA 的序列。上游引物NS1 为:5'-GTAGTCATATGCTTGTCTC-3',下游引物NS6 为:5'-GCATCACAGACCTGTTATTGCCTC-3'。PCR 扩增的反应条件为:98℃预变性3 min;98℃变性25 s,55℃退火25 s,72℃延伸1 min,30 个循环;72℃延伸10 min,4℃终止反应。取10 μL PCR 的反应产物在2%琼脂糖凝胶上电泳。产物在上海生工生物工程股份有限公司对该菌株进行基因和结构方面的鉴定。
1.2.5 分离方法与依据 为了有针对性的选育能有效地降解杜仲叶表面角质层的菌株,首先将采来的杜仲叶发水,有利于霉菌在杜仲叶表面附着繁殖。在不同的环境,室内、室外、阴暗处、土壤表面等条件下进行培养。结果发现,在实验室内置于的大盆中,覆盖有塑料膜的潮湿杜仲叶有一部分发生了病变,长出较少的菌斑。仔细观察发现,有的菌斑剥离后,杜仲叶表面有溃疡现象,说明这部分菌斑有产角质酶的能力。
植物角质是高等植物表层的一种生物聚酯,其单体主要由十六碳、十八碳的羟基脂肪酸和环氧脂肪酸组成,而角质水解的过程中降解出来的脂肪酸产物会降低培养基中的pH。能产生角质酶的菌种在一种经改良的查彼克—多克斯(Zapek-Dox)选择性培养基上生长,在这种培养基中含有作为菌种唯一碳源的纯角质和一种酸碱指示剂。能产生角质酶的菌株,在这种培养基中,由于酸碱性指示剂的作用,在菌丝的前端产生颜色变化带。这是由于菌种产生角质酶将角质降解生成脂肪酸,使培养基中的pH值下降而导致的。这是一种简便、快速而灵敏的测定真菌产生角质酶的分析法[14]。据文献报道此方法最初是用于观察引起木瓜炭疽病菌产生的缺乏角质酶突变体,但它也可以用来测定其它真菌产生的角质酶。
酸碱指示剂一般是有机弱酸或有机弱碱,其共轭酸碱形式具有明显不同的颜色。但酸碱指示剂本身具有酸碱性质,也会消耗少量的脂肪酸,用量过大会使色带的变化不明显而引起较大的误差。在不同浓度下测验指示剂,以了解由于pH 的降低所引起的指示剂最佳颜色变化。本试验中所选用指示剂指示范围见表1。
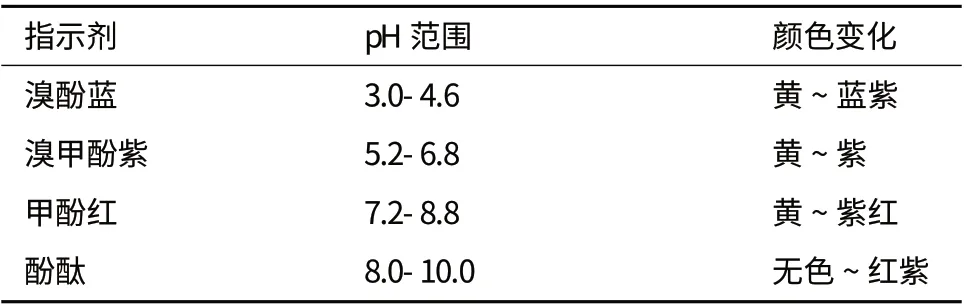
表1 指示剂指示范围
2 结果
2.1 指示剂的确定
将分离前的菌株群接入含有不同指示剂的筛选培养基上,置于27℃下培养,并观察记录各菌种生长及培养基颜色变化,结果见表2。将筛选培养基中的角质替换成0.2%的葡萄糖,其它操作处理同上,置于27℃下培养,并观察记录各菌种生长及培养基颜色变化,结果见表3。

表2 分离前的菌群在含不同指示剂的培养基中的生长情况

表3 分离前的菌群在不含角质的培养基中的生长情况
2.2 产角质酶菌种的筛选
指示剂显色反应证明,发生病变的杜仲叶通过富集培养后,得到的菌株群具有产角质酶的功能。但是这是多种菌株混合菌群。经过多次反复的分离、培养,最终分离纯化得到3 株比较强壮的菌种,分别为1 号粉红色菌、2 号粉墨绿色菌和3 号白色菌(图1)。
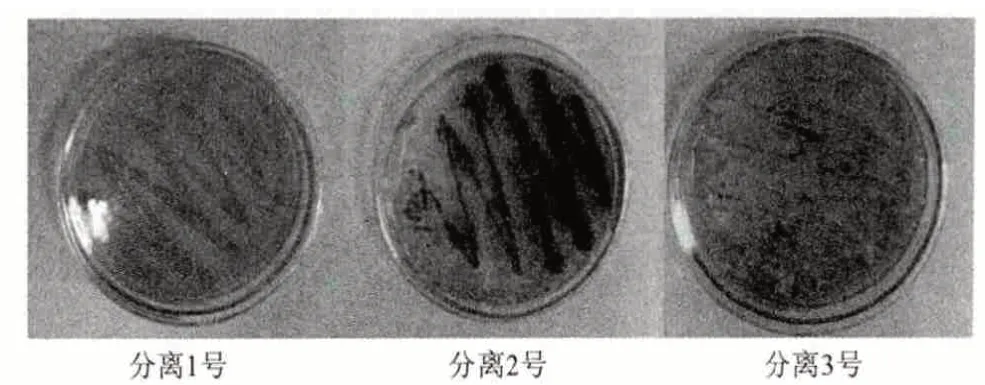
图1 分离得到的3 种菌
将所选的菌种接入加有指示剂的PDA 平板培养基上,于27℃下培养48 h 后,观察各菌种生长情况,挑选各菌种生长比较好的平板,在平板中间放入灭过菌的牛津杯,向其中加入0.1 mL 1.76%的PNB,6 h 后观察记录各菌平板颜色变化(表4)。

表4 病菌生长情况及培养基颜色变化
指示剂的颜色反应表明,1 号和2 号分离菌株具有明显的产角质酶功能,白色的3 号菌株几乎不产角质酶。为了进一步研究这3 株菌株的产角质酶功能,对它们进行了产角质酶酶活的测定。在实验室中以确有产角质酶功能的棉花枯萎病菌、苹果炭疽病菌和烟草赤星病菌为对照,同时对以上分离所得的3 种菌株进行了发酵培养,得到粗酶液进行酶活测定。从图2 中可看出,分离得到的1 号粉红菌株的酶活(产酶量)较高,仅次于棉花枯萎病菌的产酶量,高于苹果炭疽病菌和烟草赤星病菌,是一株理想的产角质酶的菌株,而2 号墨绿色菌株和3号白色菌株也具有产角质酶的功能,只是产酶量和酶活仅为1 号粉红菌株的一半,甚至更低。因此把研究的重点放在了1 号粉红色菌株。
2.3 菌株的鉴定
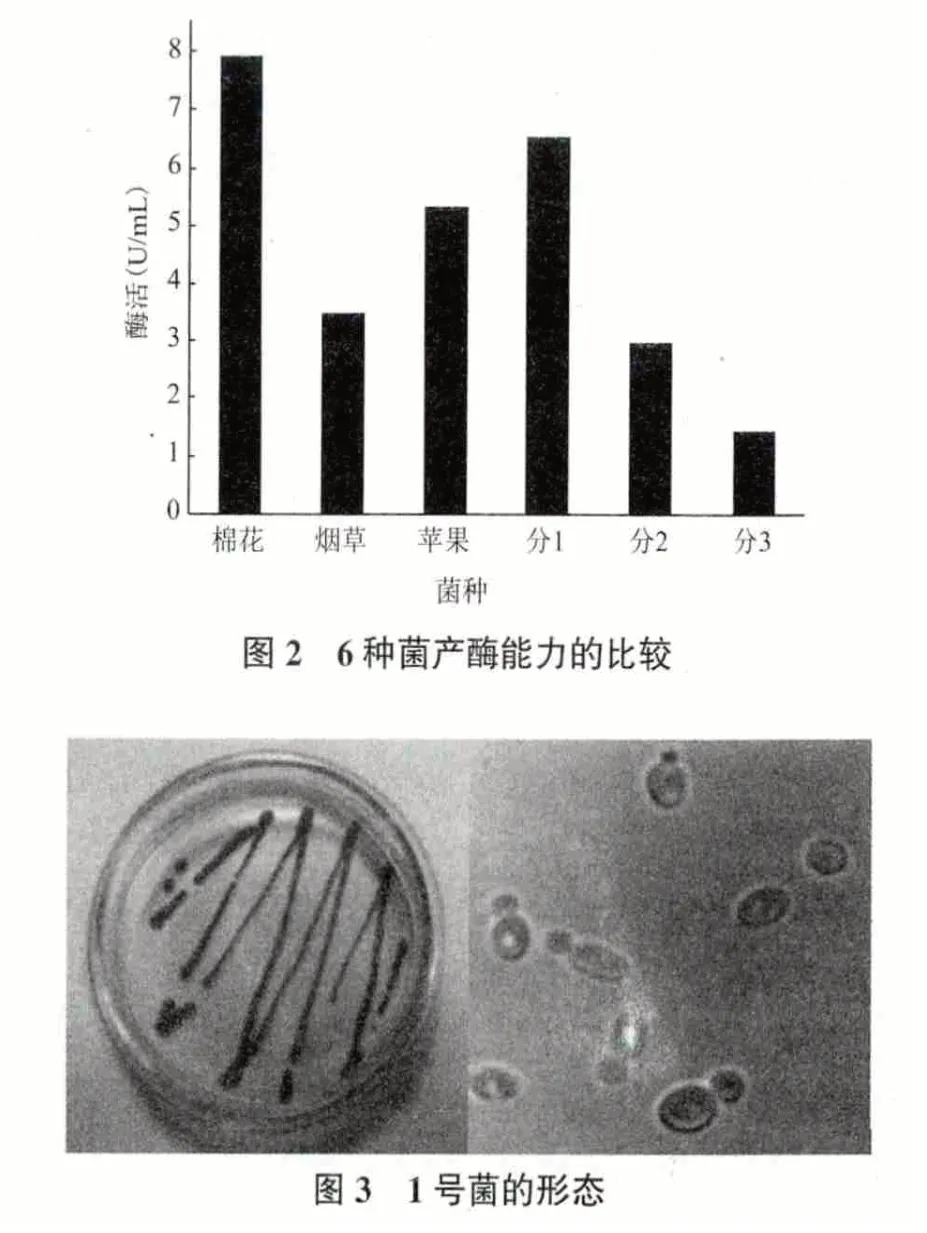
在图2 中的6 种菌中,分离1 号菌种的产酶量相对较高,仅次于实验室已有的棉花枯萎病菌,是在实验室中从杜仲叶上生长出来的未知的菌株。对这一新菌株进行了形貌、生化及结构的研究。对分离1 号菌种进行制片和显微镜观察(图3),可看出菌种表面平滑,在显微镜下呈椭圆形,出芽繁殖,有芽孢。初步判定该菌为酵母菌。
将分离得到的1 号菌株接到斜面上,培养2 d后,进行凝胶电泳分析。结果(图4)显示,扩增的DNA 片段IS180 在约1 500 bp(base pair)处有一条明亮的PCR 特征性条带,用Applied Biosystems 3730XL 测序软件得出该菌株的DNA 序列为TAAGTTTAAG-----------GGACTATCCA 且长度为1 339 bp。将所获得的DNA 在NCBI 上进行序列比对发现,有5 种菌株匹配性最高,且都属于Rhodotorula mucilaginosa(胶红酵母),它们的匹配E 值为0,Ident 值为100%,覆盖率为99%。通过比对,从NCBI 的核酸基因数据库中得到11 条与1 号菌的PCR 扩增序列匹配性、覆盖率、E 值、匹配一致性最相符的序列。
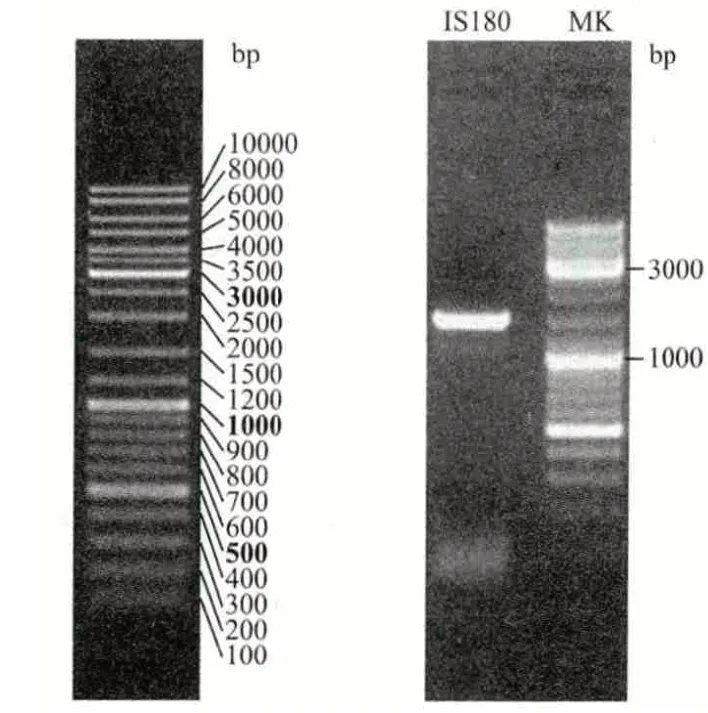
图4 DNA 电泳图
将这11 条序列与1 号菌PCR 扩增所得序列在MEGA5.0 软件上构建系统发育树(图5)。
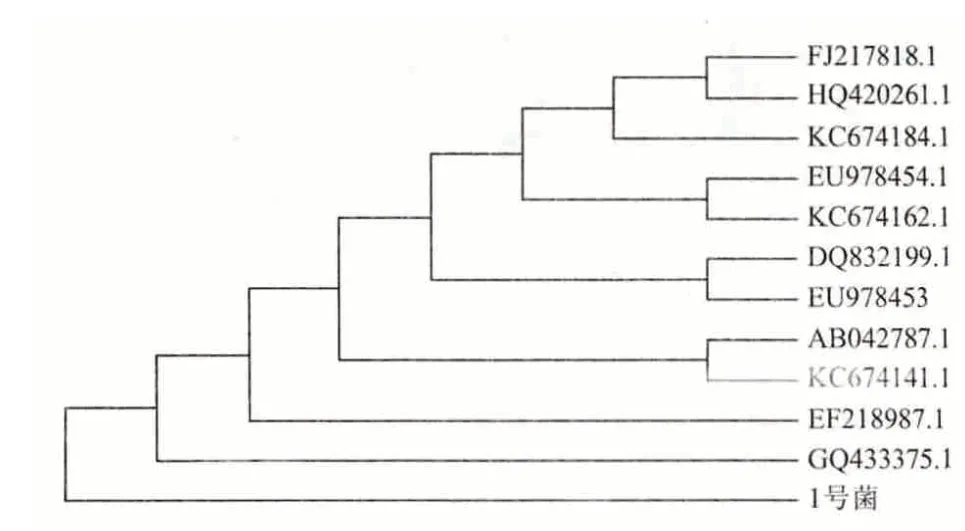
图5 MEGA5.0 构建的系统发育树
图5 显示,1 号菌与胶红酵母18S rRNA 克隆的部分序列(Uncultured fungus clone nco85d05c1 18S ribosomal RNA gene,partial sequence)都属于真菌胶红酵母(Rhodotorula mucilaginosa)。而且,分离1号菌与胶红酵母菌株ZM-1 18S rRNA 部分序列[(Rhodotorula mucilaginosa strain ZM-1 18S ribosomal RNA gene,partial sequence(序列号为GQ433375.1)]的亲缘关系最近,因此可判定1 号菌株是胶红酵母菌(Rhodotorula mucilaginosa)。
2.4 1号菌株产酶条件的优化
2.4.1 最佳碳源 在发酵基础培养基中分别添加蔗糖、可溶性淀粉、乳糖、麦芽糖、葡萄糖作为碳源,浓度均为1%,培养6 d 后测酶活,结果(图6)显示,在分离1 号菌发酵产角质酶的培养基中,最适合其发酵生产角质酶的碳源是乳糖,其次是葡萄糖,因此将该菌的发酵培养基中碳源换为乳糖。
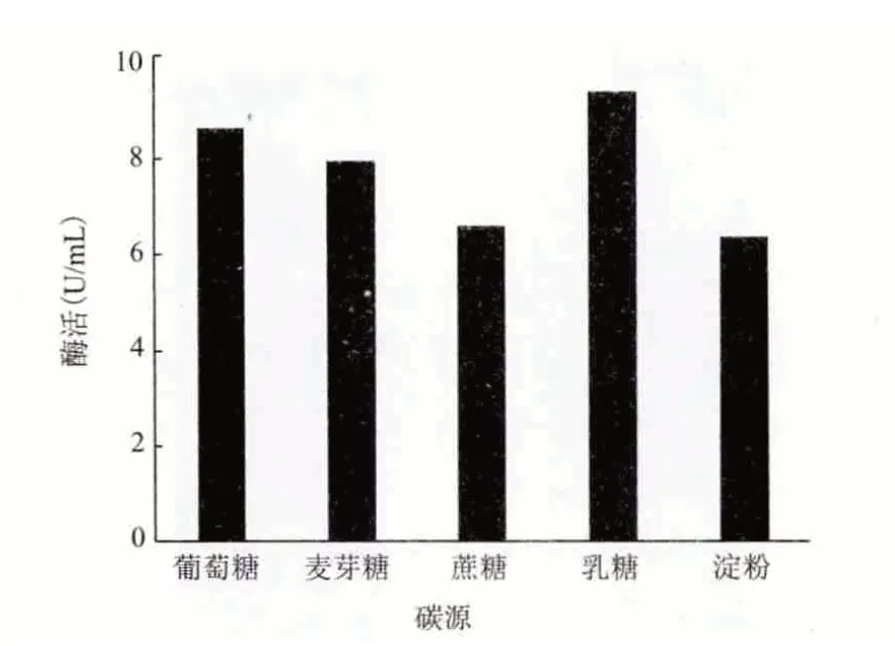
图6 最佳碳源的筛选结果
2.4.2 最佳氮源 发酵基础培养基中分别以酵母膏、蛋白胨、硫酸铵、氯化铵和硝酸钠为其氮源,浓度均为1%进行培养。培养6 d 后测酶活,结果(图7)显示,最适合该菌发酵生产角质酶的氮源是酵母粉,蛋白胨次之,因此将该菌的发酵培养基中氮源定为酵母粉。
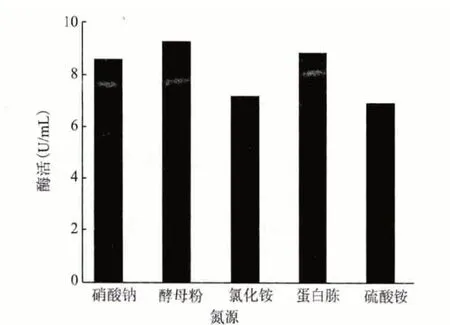
图7 最佳氮源的筛选结果
2.4.3 1 号菌的生长曲线 将保存的1 号菌接入液体PDA 培养基中振荡培养18 h 后,以10%的接种量接入装有200 mL 液体PDA 培养基的500 mL 锥形瓶中,置于30℃、160 r/min 的摇床中振荡培养,每隔2 h 取其发酵液,用未接种的空白培养液做参比,在波长为600 nm 处测定其吸光度。生长曲线(图8)表明该菌培养4 h 后开始进入对数生长期,生长繁殖迅速,培养到30 h 后,该菌进入稳定期,生长繁殖基本进入平稳期。
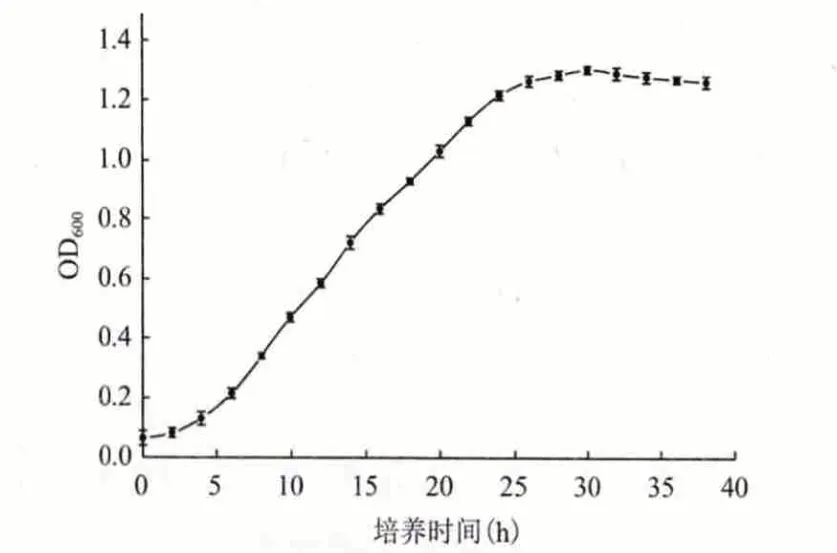
图8 生长曲线
2.4.4 产角质酶最佳时间 通过生长曲线的分析,将保存的分离1 号菌株活化1 d 后,接入装有100 mL 发酵基础培养基(将葡萄糖换为乳糖,氮源换为酵母粉)的250 mL 锥形瓶中,置于30℃摇床中,160 r/min 振荡培养,每天均匀取该菌发酵液8 mL 于5 500 r/min、4℃离心30 min,上清液用于测定酶活。结果(图9)显示,该菌的产酶最佳时间为5 d,前5 d 角质酶的含量持续增加,第5 天达到最高,之后角质酶含量有轻微的减少,最佳时间选为5 d。对比图8 和图9,说明菌的生物量达到最大时的时间很短,而产生角质酶最大酶活的时间则需5 d 时间。

图9 产酶最佳时间
2.4.5 最适温度 保存的分离1 号菌活化1 d 后,接入装有100 mL 发酵基础培养基(将葡萄糖换为乳糖,氮源换为酵母粉)的250 mL 锥形瓶中,分别置于20℃、25℃、28℃、30℃、35℃和40℃的摇床中,160 r/min 振荡培养5 d 后,按照1.2.3 的方法处理,用于测定酶活。温度对角质酶活性的影响(图10)显示,该菌最适发酵产酶的温度为28℃。
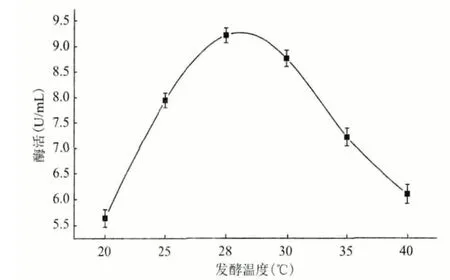
图10 产酶最适温度
2.4.6 培养基的初始pH 分别用稀盐酸和稀氢氧化钠将发酵基础培养基(碳源换为乳糖)的pH 调至:5.0、5.5、6.0、6.5、7.0、7.5 和8.0 七个梯度,将分离1 号菌种活化1 d 后,分别接入装有100 mL 不同pH 梯度培养基的250 mL 锥形瓶中,置于28℃摇床中,160 r/min 振荡培养5 d 后,测定酶活。图11 显示,初始pH 为6.5 时最有利于该菌发酵培养产角质酶。
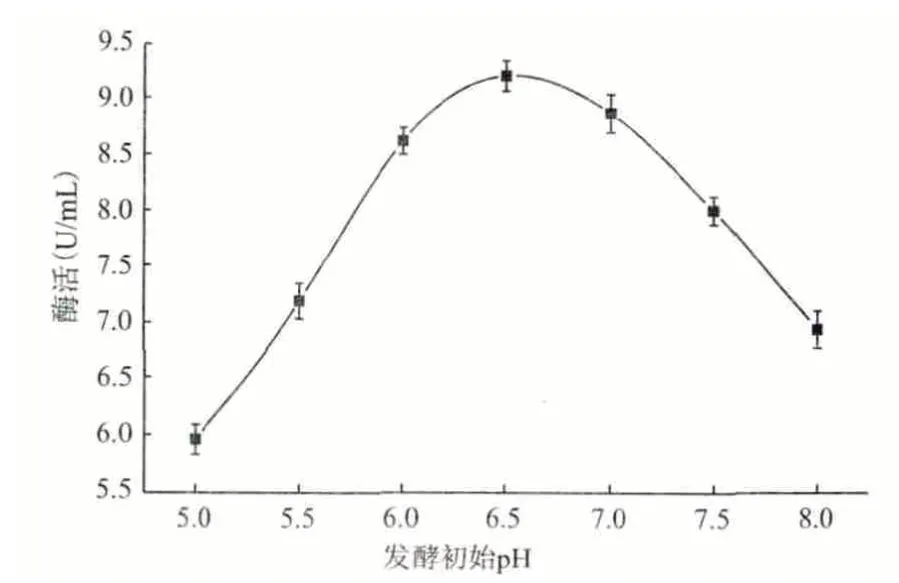
图11 初始pH 对产酶的影响
3 讨论
杜仲叶中含有大量的活性药物成分,存在叶片细胞内或者细胞间,在提取这些物质时破坏植物组织和细胞壁就可以释放出这些药物成分。杜仲叶的细胞壁可用纤维素酶和果胶酶降解,但其最外层的角质层保护叶片免受生物酶和微生物的破坏,成为如今对降解植物组织最为关注的技术难点。角质酶(EC3.1.1.74)是一种多功能酶,理论上可降解角质层。将角质酶用于酶解法提取杜仲叶中的活性成分,将比传统的碱浸法带来更大的经济和物质效益。
鉴于目前无大规模生产角质酶的研究,国内外众多学者对角质酶进行了大量研究,江南大学做了很多关于产角质酶发酵条件优化[15]、基因重组改良等方面研究[16],但还未见有分离可产角质酶的微生物的报道。本试验从杜仲中分离能产角质酶的微生物,对该菌株进行鉴定,发现该菌株属于胶红酵母。这是首次发现酵母菌能产角质酶,而且酶活较高,可达到10 U/mL 左右。这一发现具有重大的实践意义,因为通常高产角质酶的真菌都是植物的病原真菌,在生产角质酶进行培养时必须严格控制生产环境,而本研究发现的是酵母菌,可以大规模培养,不会对环境带来危害。本试验还对该菌产酶条件进行了优化,在优化培养条件的过程中,除了要选择的适合菌株生长并且不会对角质酶产量造成影响的碳源和氮源外,还要选择合适的温度、初始pH 和适当的菌龄,要适宜菌体生长及产角质酶。本研究分别考察了发酵的碳源、氮源、温度、时间和初始pH 值等对胶红酵母产角质酶的影响。在选择适当的菌龄时,一般以对数生长期的后期较为适宜,太年轻的种子接种后,往往会使发酵前期菌体生长过于缓慢,使得发酵周期延长;过老的种子虽然菌量较多,但接种后菌体会过早自溶,导致生产能力下降。所以本研究通过测定菌种的生长曲线,得出将该菌活化1 d 后的种子液接入发酵培养基培养,以保证产酶量。
由于胶红酵母比植物病原真菌更安全,因此可广泛用于角质酶制剂的生产,可以继续进行菌株的诱变研究,进一步提高该酵母的角质酶产量。
4 结论
在本试验将病变的杜仲叶分离所得的3 株菌中,分离1 号菌产角质酶量最高,经形态观察及菌种鉴定、序列比对、系统发育树的构建等判定分离1 号菌为胶红酵母(Rhodotorula mucilaginosa)。这是首次发现酵母菌产角质酶且酶活较高(10 U/mL)的菌株。对胶红酵母产角质酶的发酵条件进行优化,确定了该菌发酵生产角质酶的最佳碳源(乳糖)、氮源(酵母粉)、最佳温度(28℃)、时间(5 d)和初始pH 值(6.5)。
[1] 陆志科, 谢碧霞, 杜红岩.杜仲胶提取方法的研究[J].福建林学院学报, 2004, 4:353-356.
[2] 宫本红.杜仲叶多糖的提取分离及生物活性研究[D].贵阳:贵州大学, 2008.
[3] 龚桂珍, 宫本红, 张学俊, 等.杜仲叶和杜仲皮中化学成分的比较[J]. 西南大学学报:自然科学版, 2010, 7:167-172.
[4] 宋磊, 张学俊, 董大鹏, 王庆辉.杜仲胶性质及提取研究的进展[J].贵州化工, 2006, 4:4-8.
[5] Zhang X, Cheng C, Zhang M, et al. Effect of alkali and enzymatic pretreatments of Eucommia ulmoides leaves and barks on the extraction of gutta percha[J]. J Agric Food Chem, 2008, 56(19):8936-8943.
[6] 张守亮, 陈坚.角质酶产生菌Thermobifida fusca WSH03-11 诱变及高产突变株发酵条件优化[J].化工进展, 2006, 25(5):533-537.
[7] 张瑶, 陈晟, 吴丹, 等.角质酶及其在纺织工业中的应用[J].中国生物工程杂志, 2010, 30(9):105-109.
[8] 陈晟, 张芙华, 陈坚, 等.流加发酵对重组Bacillus subtilis 发酵生产角质酶的影响[J].中国生物工程杂志, 2010, 30(1):62.
[9] Murphy CA, Cameron JA, Huang SJ, et al. Fusarium polycaprolactone depolymerase is cutinase[J]. Applied and Environmental Microbiology, 1996, 62(2):456-460.
[10] 王义勋, 柳艳军, 陈敏, 等.真菌角质酶研究进展[J].湖北林业科技, 2011, 4:12.
[11] 薛海燕.降解玉米秸秆产纤维素酶和木质素酶菌种的筛选[J].酿酒科技, 2008, 10:44-47.
[12] Fernandes S, Johansson G, Hatti-Kaul R. Purification of recombinant cutinase by extraction in an aqueous two-phase system facilitated by a fatty acid substrate[J]. Biotechnology and Bioengineering, 2001, 73(6):465-475.
[13] Davies KA, DELorono I, Foster SJ, et al. Evidence for a role of cutinase in pathogenicity of Pyrenopeziza brassicae on brassicas[J]. Physicl Mol Plant Pathol, 2000, 57:63-75.
[14] Dickman MB, 谢旭阳.检验植物病原真菌产生角质酶的一种快速而灵敏的平板分析法[J].国外农学, 植物保护, 1988, 4:17.
[15] 张芙华, 华兆哲, 陈坚, 等.温度两阶段控制策略发酵生产重组角质酶[J].应用与环境生物学报, 2009, 15(5):730-733.
[16] 郭森.重组大肠杆菌产角质酶-CBM 发酵优化[D].无锡:江南大学, 2012.
猜你喜欢
科学大众(2022年9期)2022-06-05
山东化工(2019年20期)2019-11-19
中成药(2018年8期)2018-08-29
陕西画报(2016年1期)2016-12-01
癌变·畸变·突变(2016年5期)2016-08-22
中国塑料(2016年3期)2016-06-15
文物保护与考古科学(2016年1期)2016-04-16
小雪花·小学生快乐作文(2015年9期)2015-09-29
小雪花·小学生快乐作文(2015年9期)2015-09-29
中国医疗美容(2015年5期)2015-02-03
