
尾叶桉核心种质初步构建
2014-08-31 08:02刘德浩张卫华张方秋
华南农业大学学报 2014年6期
刘德浩, 张卫华, 张方秋
(1 华南农业大学 林学院,广东 广州 510642;2 广东省林业科学研究院,广东 广州 510520)
尾叶桉核心种质初步构建
刘德浩1,2, 张卫华2, 张方秋2
(1 华南农业大学 林学院,广东 广州 510642;2 广东省林业科学研究院,广东 广州 510520)
【目的】为了快速、准确地从丰富的尾叶桉Eucalyptusurophylla种质资源中鉴定出育种上迫切需要的优异基因,初步构建了尾叶桉核心种质,以便简化管理,提高遗传资源在尾叶桉改良中的利用效率.【方法】以尾叶桉27个种源194个家系的树高、胸径、材积等17个数量性状为依据,采用不同的取样比例(5%、10%、20%、30%和40%)、3种遗传距离计算方法(欧氏距离、标准欧氏距离和马氏距离)、4种系统聚类方法(最短距离法、最长距离法、不加权类平均法和离差平方和法)和2种取样方法(随机取样法和最小距离逐步取样法)构建尾叶桉核心种质资源库,利用均值差异百分率(MD)、方差差异百分率(VD)、变异系数变化率(VR)和极差符合率(CR)作为构建核心种质的评价指标.【结果和结论】结果表明采用10%的抽样比例、标准欧氏距离、最短距离系统聚类法和最小距离逐步取样法构建的包括25个种源100个家系的尾叶桉核心种质最能代表原有的种质群体.
尾叶桉; 种质资源; 核心种质
核心种质的概念最早是由Frankel等[1]161-170在1984年提出的,核心种质是指在整个种质资源中选择一部分样本,用最少的遗传资源数量,最大限度地代表整个遗传资源的多样性.当前,世界各国一直不断地搜集各种遗传资源构建种质资源库为育种等研究服务,此举将导致种质资源库的规模越来越大,规模巨大的种质资源库使育种家们需要花费更多的时间和资金来寻找符合育种目标的种质材料,同时有限的资金已经难以妥善保存众多的种质资源.而植物核心种质是种质资源群体中最具代表性的样本,因此,在育种工作中可以优先对核心种质进行保存、评价和利用,从而解决种质资源过大、难以进行管理和利用的难题.目前我国已在多种植物上开展核心种质的研究工作,其中以农作物为首,涉及到水稻、大麦、玉米、棉花、油菜、大豆、芝麻等,而林木核心种质的构建工作严重滞后于农作物,仅限于腊梅、桃和茶树等[2-4].尾叶桉是优良的速生用材林树种,其材质优良、用途广泛,是华南地区杂种桉(例如3229)的优异亲本材料,同时也是培育新一代优良杂种的骨干亲本[5].外引树种资源的有效保存和利用一直是个难题,保存的数量、方法、地点对于尾叶桉来说都至关重要,核心种质的理论与实践是种质资源学发展的重要方向和重大进展,用最小的样本数代表最大的遗传多样性,对于尾叶桉资源保存、高效育种均具有重要意义.本文利用尾叶桉试验林的27个种源194个家系为材料,以树高、胸径、干形和材性等17个指标的数据为依据,通过比较分析不同取样比例、遗传距离、聚类方法和取样策略构建出的尾叶桉核心种质各评价参数值,初步构建出较为合适的尾叶桉核心种质资源库.
1 材料与方法
1.1 材料
试验地设在广东省四会市贞山区大南山林场,位于东经112°45′,北纬23°22′.试验林建于2008年8月,参试的194个尾叶桉家系分别来自于27个不同种源,包括印度尼西亚、巴西、菲律宾、马来西亚和南非等地.试验采用随机完全区组设计,单株小区,50次重复,株行距2 m×3 m.穴植,穴的规格为50 cm×50 cm×50 cm.造林时每穴施基肥桉树有机肥1 kg.第1年施复合肥2次,第2年追肥1次,用量为0.25 kg.造林后前2年,每年1~2次抚育,于每年春末夏初和秋季进行,并松土扩穴.
1.2 方法
1.2.1 性状测定 每个家系随机抽取10个单株,测定每个单株的基部直径、胸径、树高、4 m处直径、枝下高、分枝大小、尖削度、树皮厚度、分枝数量、枝痕形状、干形、分枝均匀度、分枝角度、冠幅大小、叶片密度、树皮开裂方式,木质材性数据利用ST-300木质检测仪测定.干形指标分4个数量等级:Ⅰ级:主干通直圆满,得4分;Ⅱ级:主干直、不圆满,得3分;Ⅲ级:主干稍弯曲、不圆满,得2分;Ⅳ级:主干有两个以上弯曲,得1分.分枝均匀度分级标准:Ⅰ级:侧枝细小、树冠匀称,得3分;Ⅱ级:侧枝中等,树冠匀称,得2分;Ⅲ级:有明显大枝、树冠不均,得1分.数量性状采用标准化数据,1级≤X-2δ,10级>X+2δ,1~10级中间每级的间差为0.5δ,其中X为各性状平均值,δ为标准差.
1.2.2 分组方法 由于地理种源可以为生物多样性提供间接根据,所以本文采用地理种源分组.
1.2.3 取样比例 设定5%、10%、20%、30%和40%共5个取样比例.
1.2.4 遗传距离计算 遗传距离分别采用欧式距离(Euclidean distance,Euclid)、标准欧氏距离(Standardized Euclidean distance,Seuclid)和马氏距离(Mahalanobis distance,Mahal)[6]3种方法进行计算.
1.2.5 聚类方法 聚类方法分别采用不加权类平均法(Unweighted pair-group average method,Average)、最长距离法(Complete distance method,Complete)、最短距离法(Single distance method,Single)和离差平方和法(Ward’s method,Ward)进行聚类[7-8].
1.2.6 取样策略 按照胡晋等[9]提出的最小距离逐步取样法(Least distance stepwise sampling,LDSS)和多次聚类随机取样法(Stepwise cluster based on random sampling,SCR)进行样本抽取.
1.2.7 评价方法 通过使用胡晋等[9]提出的均值差异百分率(Mean difference percentage,MD)、方差差异百分率(Variance difference percentage,VD)、极差符合率(Coincidence rate of range,CR)和变异系数变化率(Changeable rate of coefficient of variation,VR)4个评价参数,评价不同方法构建出的尾叶桉核心种质的代表性,4个评价参数的计算方法分别为:
(1)
式(1)中,pij是第i个性状第j种表现型的频率,m是第i个性状表现型的数目,n是数量性状总数;
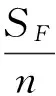
(2)
式(2)中,SF是核心子集与原始群体进行F测验得到的方差差异显著(α=0.05)的性状数,n是数量性状总数;
(3)
式(3)中,RC(i)是核心子集第i个性状的极差,RI(i)是原始群体第i个性状的极差,n是数量性状总数;
(4)
式(4)中,CVC(i)是核心子集第i个性状的变异系数,CVI(i)是原始群体第i个性状的变异系数,n是数量性状总数.Hu等[10]提出核心种质的MD小于20%,CR大于80%,就可以认为所筛选出的核心种质能够较好地代表原始群体的遗传多样性,并且MD越小,CR、VD和VR越大,核心子集越能够代表原始群体的遗传多样性.
2 结果与分析
2.1 不同取样比例构建的尾叶桉核心种质结果比较
在采用标准欧式距离、最短距离聚类方法和最小距离逐步取样法的前提下,抽样比例分别为5%、10%、20%、30%和40%的尾叶桉核心种质评价参数值见表1.由表1可以看出,在5种不同取样比例下的核心种质MD均接近于0,均符合构建核心种质的第1个条件,但是CR只有10%、20%、30%和40%这4种取样比例符合构建要求,5%取样比例下的CR只有75.83%,低于构建核心种质CR大于80%的要求.因此,5%取样比例明显不符合构建核心种质的要求.由表1还可以看出,10%和30%这2种取样比例的CR明显高于20%和40%取样比例,均达到90%以上,因此优先考虑10%和30%这2种取样比例;而10%取样比例下的CR和VR均大于30%取样比例下的CR和VR,而当2种取样比例下的MD值小于20%且相近时,CR、VR越大其代表性越高.由于较大的样本量无法满足生产和研究的需要,对5种取样比例下的核心种质数量进行比较发现,只有10%的取样比例是构建尾叶桉核心种质的合理取样比例,10%的取样比例用较小的样本量较好地保存了原始资源的遗传多样性.
表1不同取样比例下构建的核心种质评价参数值
Tab.1Evaluatingparametersofcorecollectionbasedondifferentsamplingpercentages
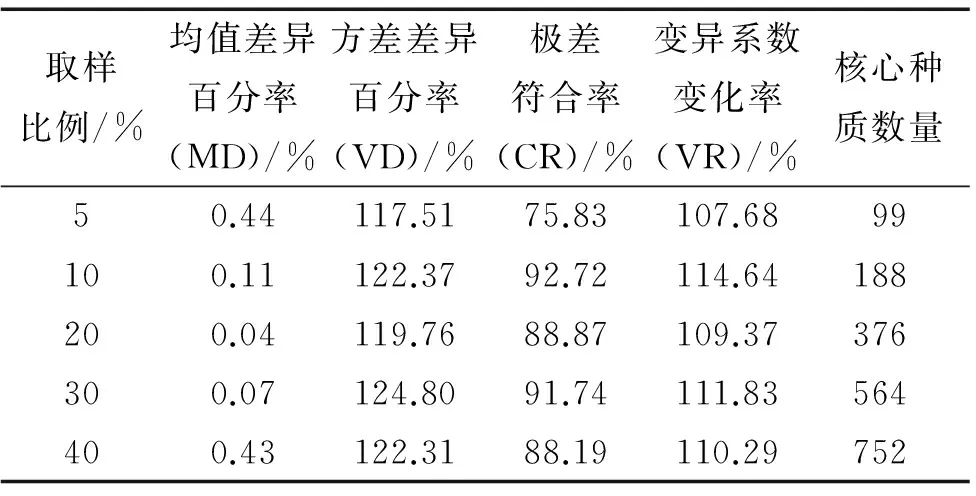
取样比例/%均值差异百分率(MD)/%方差差异百分率(VD)/%极差符合率(CR)/%变异系数变化率(VR)/%核心种质数量50.44117.5175.83107.6899100.11122.3792.72114.64188200.04119.7688.87109.37376300.07124.8091.74111.83564400.43122.3188.19110.29752
2.2 不同遗传距离构建的尾叶桉核心种质结果比较
在确定10%取样比例、最短距离聚类方法和最小距离逐步取样法的前提下,采用欧式距离、标准欧氏距离和马氏距离构建的尾叶桉核心种质评价参数值见表2.结果表明,采用3种遗传距离构建的尾叶桉核心种质的MD与CR均符合构建核心种质的要求,表明3种遗传距离构建的尾叶桉核心种质均能够较好地代表原始种质的遗传多样性.虽然3种方法构建的核心种质符合构建要求,但是在数值上还是存在明显的差异,标准欧式距离构建的核心种质的MD、CR明显高于欧式距离和马氏距离的构建结果,尽管在VD值上标准欧式距离稍有不足,但标准欧氏距离的VR值又优于欧氏距离和马氏距离.因此,综合看来标准欧式距离是构建尾叶桉核心种质的合理遗传距离.
表2 3种遗传距离下构建的核心种质评价参数值
Tab.2Evaluatingparametersofcorecollectionbasedonthreekindsofgeneticdistance%

遗传距离均值差异百分率(MD)方差差异百分率(VD)极差符合率(CR)变异系数变化率(VR)欧式距离0.16134.5391.54116.93标准欧式距离0.11114.6492.72122.37马氏距离0.15131.4091.96116.21
2.3 不同聚类方法构建的尾叶桉核心种质结果比较
在确定10%取样比例、标准欧式距离计算遗传距离和最小距离逐步取样法的前提下,采用最短距离法、最长距离法、不加权类平均法和离差平方和法4种聚类方法构建的尾叶桉核心种质遗传差异见表3.采用4种不同聚类方法构建出的尾叶桉核心种质均符合构建核心种质的基本要求,其MD接近于0,CR均达到90%以上,能够代表原始资源的遗传多样性.由表3可以看出,最短距离法和最长距离法的CR值高于不加权类平均法和离差平方和法,最短距离法的VR明显高于其他3种聚类方法.综合来看,4种聚类方法构建出的尾叶桉核心种质以最短距离法最优,离差平方和法最差,不加权类平均法与最长距离法相差不大.因此,确定最短距离法为构建尾叶桉核心种质的合理聚类方法.
表3 4种聚类方法构建的核心种质评价参数值
Tab.3Evaluatingparametersofcorecollectionbasedonfourclusteringmethods%
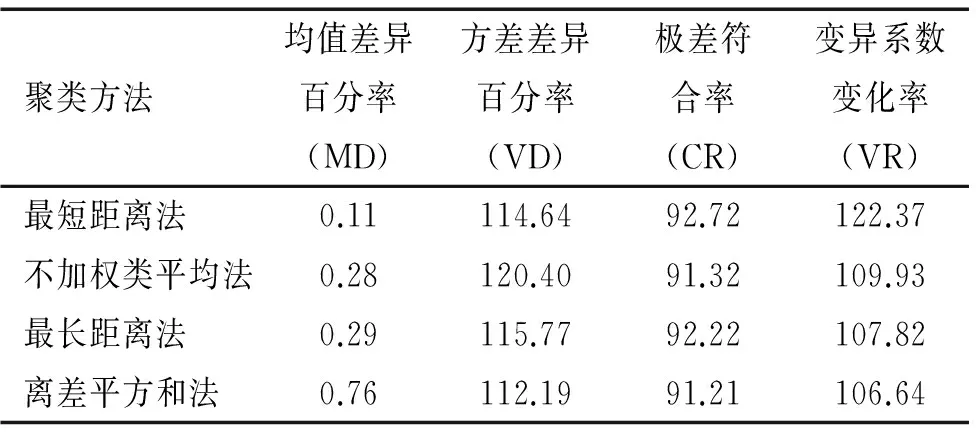
聚类方法均值差异百分率(MD)方差差异百分率(VD)极差符合率(CR)变异系数变化率(VR)最短距离法0.11114.6492.72122.37不加权类平均法0.28120.4091.32109.93最长距离法0.29115.7792.22107.82离差平方和法0.76112.1991.21106.64
2.4 不同取样策略构建的尾叶桉核心种质结果比较
在确定10%取样比例、标准欧式距离和最短距离聚类方法的前提下,采用多次聚类随机取样法和最小距离逐步取样法构建出的尾叶桉核心种质评级参数值见表4.结果表明,2种取样策略构建的尾叶桉核心种质的MD接近于0,CR均大于80%,能够较好地代表原始群体的遗传多样性.但是多次聚类随机取样法构建的核心种质的CR只有82.71%,勉强符合构建核心种质的要求;而最小距离逐步取样法构建的核心种质CR则高达92.72%,其MD、VD和VR与前者相比较均存在不同程度的优越.因此,在尾叶桉核心种质的构建过程中,最小距离逐步取样法比多次聚类随机取样法更合适.
表4 2种取样策略构建的核心种质评价参数值
Tab.4Evaluatingparametersofcorecollectionbasedontwosamplingstrategies%

取样策略均值差异百分率(MD)方差差异百分率(VD)极差符合率(CR)变异系数变化率(VR)多次聚类随机取样法(SCR)0.82109.1382.71102.47最小距离逐步取样法(LDSS)0.11114.6492.72122.37
2.5 尾叶桉核心种质的组成
根据前文研究结果,确定出构建尾叶桉核心种质的合理取样比例为10%,合理遗传距离计算方法为标准欧氏距离,合理聚类方法为最短距离聚类法,合理取样方法为最小距离逐步取样法.本文基于5种取样比例、2种取样方法、3种遗传距离计算方法、4种聚类方法构建出的尾叶桉核心种质资源库涵盖了尾叶桉25个种源的100个家系的188个单株,入选比例为10%.尾叶桉核心种质种源家系具体分布情况见表5.在表5中,入选家系占种源内家系比例达到80%以上家系的种源地多数比较分散,分别分布于巴西、南非、印度、泰国、老挝等地,而入选率低于80%的家系则多数来自于印度尼西亚种源,这表明构建出的尾叶桉核心种质与原始群体相比较,核心种质资源库较好地剔除了重复的遗传资源,即用最小的样本量最大限度地代表了尾叶桉原始种质的遗传多样性.
表5尾叶桉核心种质组成
Tab.5CompositionofEucalyptusurophyllacorecollection
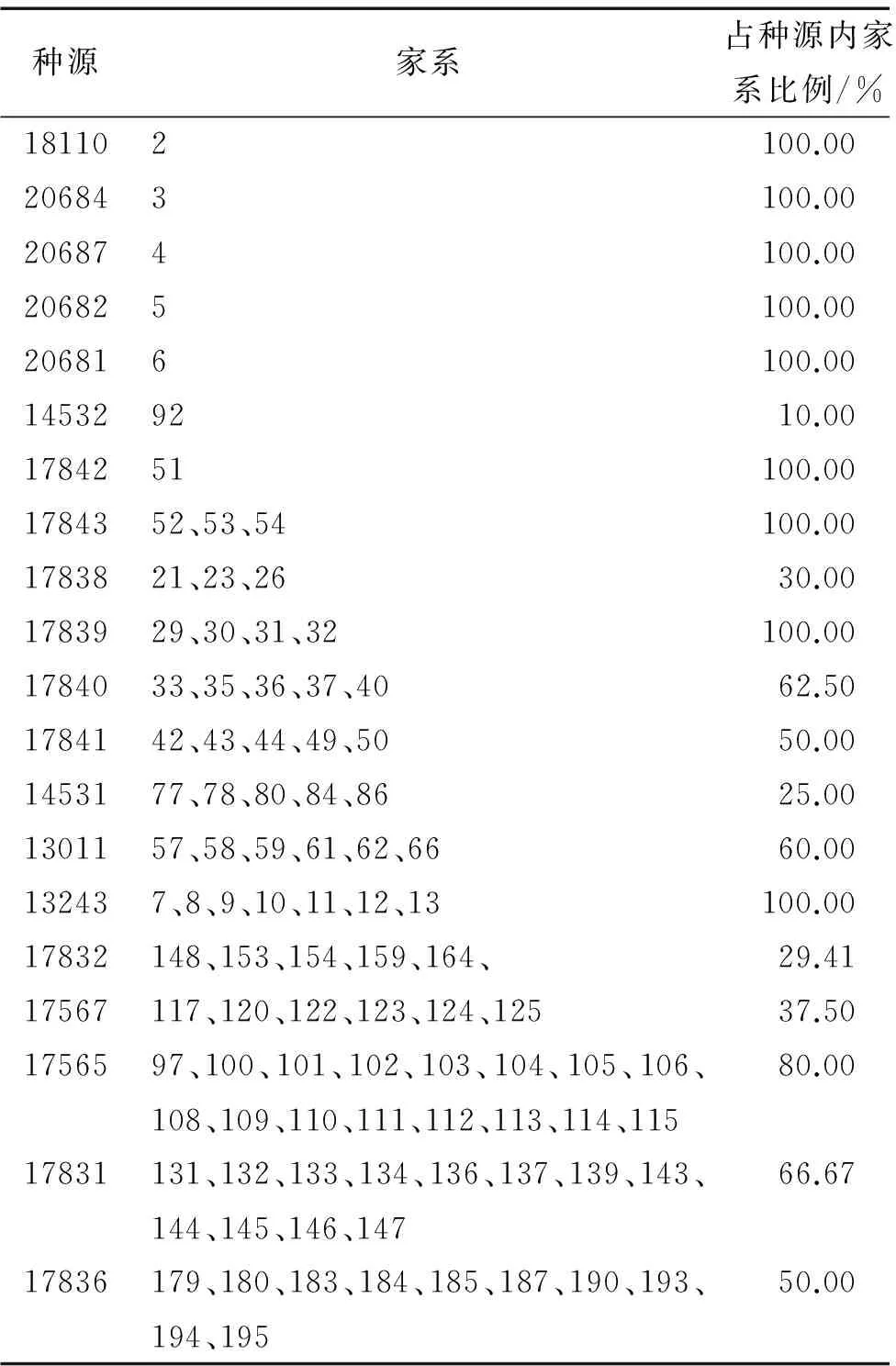
种源家系占种源内家系比例/%181102100.00206843100.00206874100.00206825100.00206816100.00145329210.001784251100.001784352、53、54100.001783821、23、2630.001783929、30、31、32100.001784033、35、36、37、4062.501784142、43、44、49、5050.001453177、78、80、84、8625.001301157、58、59、61、62、6660.00132437、8、9、10、11、12、13100.0017832148、153、154、159、164、29.4117567117、120、122、123、124、12537.501756597、100、101、102、103、104、105、106、108、109、110、111、112、113、114、11580.0017831131、132、133、134、136、137、139、143、144、145、146、14766.6717836179、180、183、184、185、187、190、193、194、19550.00
3 结论
一个合适的取样比例能够筛选出最具代表性的核心样本群体,所获得的核心群体能够最大限度地保持原始群体的遗传多样性[1]249-257.因此确定合适的取样比例是构建核心种质的关键,过高的取样比例一方面可能包含冗余较高的样本,另一方面也不利于开展生产和科研工作,而过低的取样比例则不足以代表原始群体的遗传多样性并将导致重要核心材料的丧失[11].在一个确定的取样比例下,合理的遗传距离计算方法与系统聚类方法是成功构建核心种质的必要条件,本研究中标准欧氏距离与马氏距离构建出的尾叶桉核心种质均具有较好的代表性,但标准欧式距离构建的尾叶桉核心种质的VR为122.38%,而马氏距离核心种质VR为116.21%.综合4种聚类方法来看,最短距离法构建的尾叶桉核心种质能极显著地增加各性状的变异系数,对VR的提高显著优于其他3种聚类方法.物种遗传多样性的分布是具有一定的组织与结构的,对核心子集进行取样时采用随机取样法和其他取样方法所构建出的核心种质具有非常大的差别,在本研究中最小距离逐步取样法与多次聚类随机取样法相比具有显著的优越性,因此本文在构建尾叶桉核心种质的过程中不能够采用多次聚类随机取样法,随机取样法的对照意义远大于实际意义.
核心种质最大程度地剔除了种质资源中的遗传重复,以极少的种质数量涵盖了原始资源群体中的全部或大多数遗传变异类型,核心种质实际上是一批样本量小而遗传多样性高且具有较高代表性的样品集[12].核心种质有效地解决了当前资源收集量与资源深入评价及有效利用之间所存在的突出矛盾,极大地推动和促进了种质资源研究的进一步发展.表型与分子水平的结合是核心种质走向应用的有效路径,本研究中通过表型性状构建的尾叶桉核心种质有必要与分子标记方法结合起来进行下一步的分析和评价[13].资源学与育种学的有效结合是一个重要的发展趋势,资源高效保存与高效育种的有机结合即优异核心种质育种在资源评价利用中具有非常重要的作用,努力做到核心种质田间保存与离体保存的结合是下一步的研究重点之一.
[1]FRANKEL O H, BROWN A H D.Plant genetic resources today:Acritical appraisal[M]∥HOLDEN J H W, WILLIAMS J T.Crop genetic resources: Conservation and evaluation.London: George Allen and Unwin, 1984: 161-170.
[2]PRASADA R K E, RAMANATH R V.The use of characteriza-tion data in developing a core collection of sorghum[M]∥HODGKIN T, BROWN A H D, VAN HINTUM M J L, et al.Core collections of plant genetic resources.Chichester:A Wiley-Sayee Publications,1995: 109-116.
[3]福建省农业科学院茶叶研究所.茶树品种志[M].福州:福建人民出版社,1979.
[4]刘旭,范传珠,谭富娟,等.小麦特殊遗传材料核心样品的建立[J].中国学术期刊文摘,1996,16(4):120.
[5]潘志刚,游应天.中国主要外来树种引种栽培[M].北京:北京科学技术出版社,1994: 606-610.
[6]张秀荣,郭庆元,赵应忠,等.中国芝麻资源核心收集品研究[J].中国农业科学,1998,31(3):49-55.
[7]刘旭,马缘生,谭富娟,等.小麦特殊遗传材料核心样品的建立[J].植物遗传资源科学,2000,1(2):1-8.
[8]李晓玲,李金泉,卢永根.水稻核心种质的构建策略研究[J].沈阳农业大学学报,2007,38(5):681-687.
[9]胡晋,徐海明,朱军.基因型值多次聚类法构建作物种质资源库[J].生物数学学报,2000,15(1):103-109.
[10]HU Jin, ZHU Jun, XU Haiming.Methods of constructing core collections by stepwise clustering with three sampling strategies based on the genotypic values of crops[J].Theor Appl Genet, 2000,101(1/2): 264-268.
[11]BROWN A H D.Core collections: A practical approach to genetic resources management [J].Genome, 1989, 31(2):818-824.
[12]WANG Jiancheng, HU Jin, XU Haiming, et al.A strategy on constructing core collections by least distance stepwise sampling[J].Theor Appl Genet, 2007, 115(1): 1-8.
[13]MILENE S, MIRIAN P M, MARIA B S,et al.Genetic diversity of aCoffeaGermplasm Collection assessed by RAPD markers[J].Genet Resour Crop Evol, 2008,55: 901-910.
【责任编辑李晓卉】
PreliminaryconstructionofcorecollectionofEucalyptusurophyllagermplasm
LIU Dehao1,2, ZHANG Weihua2, ZHANG Fangqiu2
(1 College of Forestry, South China Agricultural University, Guangzhou 510642, China;2 Guangdong Academy Forestry, Guangzhou 510520, China)
【Objective】 In order to rapidly and precisely identify the excellent gene for the breeding ofEucalyptusurophylla, the construction of core collection ofE.urophyllawas necessary.Establishing a core collection ofE.urophyllacould simplify the management and enhance the utilization of genetic resources inE.urophyllaresearch programs.【Method】 Based on 17 quantitative traits, height, diameter and volume of 194 families from 27 provenances ofE.urophylla, core collections were constructed using different sampling proportions (5%, 10%, 20%, 30%, and 40%), 3 distances (Euclidean distance, Standardized Euclidean distance, and Mahalanobis distance), 4 hierarchical clustering methods (Single distance method, Complete distance method, Unweighted pair-group average method and Ward’s method), and 2 sampling strategies (random sampling and least distance stepwise sampling) were employed.The genetic variation of the quantitative traits among these core collections was compared by evaluating the mean difference percentage (MD), variance difference percentage (VD), changeable rate of coefficient of variation (VR) and coincidence rate of range (CR) of the traits.【Result and conclusion】 The results showed that the primary core collection was constructed with 10% sampling proportion, standardized Euclidean distance, Single distance method, and least distance stepwise sampling, which was composed of 100 families from 27 provenances ofE.urophyllaand the best representative of total germplasm.
Eucalyptusurophylla; germplasm; core collection
2013- 06- 13优先出版时间2014- 09- 30
优先出版网址:http:∥www.cnki.net/kcms/detail/44.1110.S.20141003.1242.017.html
刘德浩(1988—),男,硕士研究生,E-mail :15013126056@139.com; 通信作者: 张卫华(1977—),女,教授级高工,博士,E-mail: zwh523@sinogaf.cn
林业公益性行业科研专项(201004009-7,201104003)
刘德浩, 张卫华, 张方秋.尾叶桉核心种质初步构建[J].华南农业大学学报,2014,35(6):89- 93.
S722.3
A
1001- 411X(2014)06- 0089- 05
猜你喜欢
今日农业(2022年13期)2022-09-15
中国饲料(2021年13期)2021-12-05
中国牛业科学(2021年3期)2021-09-29
中国畜牧杂志(2019年3期)2019-03-19
乡村科技(2019年7期)2019-02-21
中国麻业科学(2018年6期)2018-04-09
中学生数理化·七年级数学人教版(2017年10期)2017-04-23
东方教育(2016年8期)2017-01-17
西南农业学报(2016年5期)2016-05-17
广西林业科学(2016年3期)2016-03-16
