
贡嘎山雅家埂峨眉冷杉林线种群的时空动态
2014-09-19 10:58梁一鸣王根绪
生态学报 2014年23期
冉 飞,梁一鸣,2,杨 燕,杨 阳,王根绪,*
(1. 中国科学院水利部成都山地灾害与环境研究所山地表生过程与生态调控重点实验室, 成都 610041; 2. 中国科学院大学,北京 100049)
自工业革命以来,由于人类活动的影响,全球大气CO2、甲烷(CH4)和氧化亚氮(N2O)等温室气体浓度显著增加,由此造成全球气候系统的变暖已是毋庸置疑的事实。预计到本世纪末,全球平均气温将升高1.8 — 4.0 ℃,其中高纬度、高海拔地区的增温幅度更大[1]。如此大的增温幅度将会对植被产生巨大的生态效应:温度能直接或间接的影响植物的多种生理过程和功能性状,如光合色素含量、气孔导度、蒸腾速率、光合速率、表观量子效率、水分利用效率、植物的生长以及生物量分配等[2- 8],进而影响植物的定居、存活、发育和繁殖[9- 10],改变植物群落的物种组成和分布范围[11- 12]。
高山林线作为山地郁闭林与树种线之间的生态过渡带[13],对气候变化异常敏感[14- 16]。已有大量研究表明气候变暖会导致林线向上扩张,植被带上移[11,17- 20],但同时有研究表明气候变暖后林线无明显变化[21- 22], 或者其变化不是由气候变暖引起的[23- 24],学术界对于气候变暖是否会导致林线上移还存在一定争议[22,24]。尽管有影像资料表明青藏高原东南部横断山区的林线在过去一段时间有显著地爬升,但缺乏对该区域林线动态的详细研究[22, 25]。
作为横断山区众多高山典型代表的贡嘎山,位于青藏高原东南缘, 横断山系大雪山脉的中段, 东临大渡河, 西控雅砻江,其主峰高达7556 m,而东坡山麓海拔仅为1600 m,近6000 m的海拔落差导致贡嘎山东坡的植被具有丰富的垂直带谱,是开展植被-气候关系研究的天然实验室。气象监测数据表明,贡嘎山东坡峨眉冷杉(AbiesfabriCraib)成熟林分布区的年均温自20世纪80年代末以来一直以0.35 ℃/10a的速度增加,其增温速率远高于北半球和全球的平均水平。因此,研究贡嘎山东坡峨眉冷杉种群海拔分布范围对气候变暖的响应具有非常重要的理论和现实意义:一方面,该研究有助于进一步揭示植物对气候变暖的响应,为进一步研究全球气候变化对亚高山针叶林生态系统的影响提供科学依据;另一方面,该研究还有助于提高我们对气候变暖背景下植物生长、繁殖和更新的认识,增强我们在全球环境变迁条件下管理这些生态系统的能力,促进我国生态环境建设。
1 研究区域与对象
本研究以贡嘎山东坡雅家埂地区峨眉冷杉的林线种群为研究对象。该区地处中国东部亚热带湿润季风区与青藏高原寒冷气候的过渡带,据中国科学院贡嘎山高山森林生态系统观测试验站多年气象观测数据显示,贡嘎山东坡3000 m海拔多年平均气温3.8 ℃,最冷月(1月)和最热月(7月)平均温度分别为-4.3 ℃和11.9 ℃,年平均降水量1940 mm,年际变化不显著,但季节分布集中且不均匀,集中在6—12 月;年平均蒸发量1578.6 mm,年平均空气相对湿度90.2%。气候和植被呈寒温带特征,土壤层较厚,主要是含沙量高、渗透性强的山地暗棕壤[26]。作为贡嘎山东坡亚高山地区的优势树种之一的峨眉冷杉,常分布于海拔2800—3800 m的阴坡或半阴坡,在保持水土、维护区域生态系统的稳定性方面起着重要作用。雅家埂地区峨眉冷杉分布于海拔2900 m以上,低海拔地区曾被大规模砍伐,海拔3200 m以上始见纯林(多为次生林),3500 m及以上区域因海拔较高、交通不便等原因受到人为扰动较小,该区林线为郁闭型林线(即森林突然在林线位置停止向上爬升,使林线成为一条划分树林与草甸或灌丛的截然界限),其树线在阳坡可达3650 m,阴坡可达3800 m。
调查地点位于泸定县天保工程雅家埂管护站东西两侧林线区域,调查样地基本信息详见表1。该区峨眉冷杉形成纯林,高15—28 m,盖度45%—75%;林下灌木层种类较为简单,只有一层,毡毛栎叶杜鹃(Rhododendronphaeochrysumvar.levistratum)占绝对优势,盖度达60%—80%,偶有冰川茶藨子(Ribesglaciale)和花楸(Sorbusrufopilosa);草本层以鹿蹄草(Pyrolarotundifoliassp.)、猪殃殃(Galiumacutum)和东方草莓(Fragariaorientalis)为主,总盖度1%—5%。
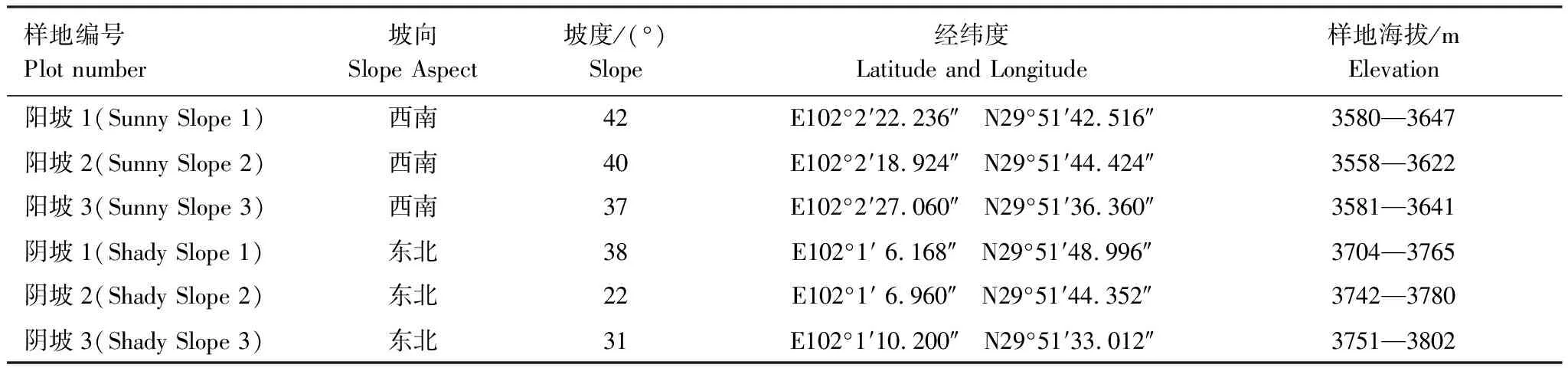
表1 调查样地基本情况
2 研究方法
2.1 样方设置与调查
在阴阳坡林线位置分别设置3个3000 m2的样方,以树线位置为样方上端,水平方向宽30 m,垂直方向长100 m,相同坡向的两个样方间隔在200 m以上。确定样方后,用GPS测定样方顶端与底端的海拔、样地经纬度,用坡度仪测定坡度,目测法测定乔木层郁闭度和灌木层、草本层的盖度,然后分别对样方中每一株立木和死树进行测量。
测定每一株胸径(DBH)>2.5 cm的峨眉冷杉(包括枯立木和倒木)的胸径、坐标,然后以十字交叉法钻取每一株胸径>5.0 cm的立木树芯,根据两条交叉树芯确定准确树龄,幼树(2.5 cm≤DBH≤5.0 cm)的测定则是通过数茎干上的节间数来确定树龄[22]。参照沈泽昊等[27]的方法,根据各径级立木的胸径和树龄数据,建立立木的胸径-树龄指数模型(图1),并以此模型估算“空心”大树(无法钻取完整的树芯)的树龄。枯立木和倒木根据其分解程度及树干直径大致估计其在50a前和100a前的生活状态。
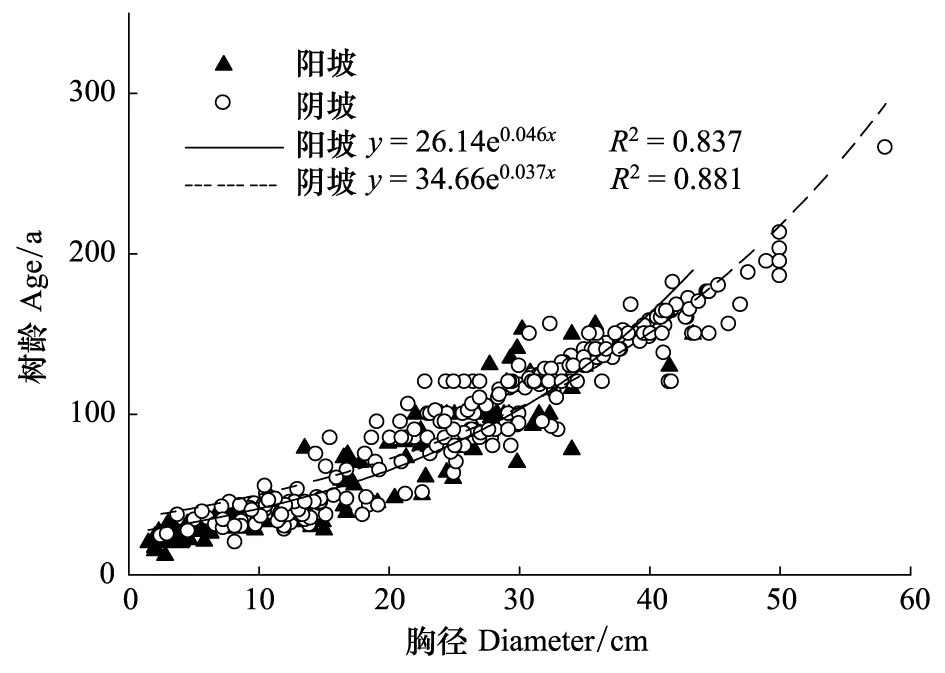
图1 冷杉立木的胸径-树龄关系
2.2 树线与林线位置的确定
树线和林线位置的确定参照Liang等[22]的标准,树线是指树高大于2 m、树冠对称的乔木生长上限(通过数节间和测树高,发现林线附近能达到这一标准的峨眉冷杉的最小树龄为16a)。林线则是指树高大于5 m、盖度不低于30%的乔木林分布的最高海拔(在30 m(水平方向)×10 m(垂直方向)范围内)。50a前、100a前树线位置的确定是根据所调查区域峨眉冷杉当前的树龄和空间分布往前逆推50a、100a得到的,逆推方式为目前树龄减去50a或100a后,树龄大于16a的乔木分布上限。
2.3 数据处理
采用SPSS Statistics 16.0 统计软件对阴坡和阳坡的树线(林线)的海拔高度、林线附近峨眉冷杉种群密度进行独立样本t检验,并用字母法标记,显著水平为P<0.05;冷杉立木的胸径-树龄指数回归方程的建立通过SPSS Statistics 16.0中regression-nonlinear分析完成。
3 结果与分析
3.1 林线峨眉冷杉种群龄级结构
通过数主干的节间得知,林线附近胸径≥2.5 cm的峨眉冷杉最小树龄为16a。按照50a为间隔绘制的林线峨眉冷杉种群年龄结构如图2所示:树龄在16—50a的峨眉冷杉最多,其次为100—150a,再次为50—100a,150a以上的峨眉冷杉最少。

图2 林线冷杉种群年龄结构
3.2 林线峨眉冷杉种群的时空动态
目前、50a前、100a前林线附近峨眉冷杉种群的空间分布如图3所示,从该图可以看出,阴阳坡树线高度在过去100a间并无明显爬升,但林线附近峨眉冷杉种群密度在过去50a间有显著升高(图4)。
3.3 林线峨眉冷杉种群的坡向分异
如图5所示,阴阳坡之间树线海拔高度存在显著差异,阴坡和阳坡的树线海拔高度分别为(3771.5±7.7)m和(3635.7±7.8)m,阴坡比阳坡高135.8 m;林线高度也存在显著的坡向差异,阴阳坡林线海拔高度分别为(3770.4±6.6)m和(3617.9±10.6)m,阴坡林线比阳坡高约152.5 m。然而,在同一坡向,林线与树线之间的海拔高差不大。

图3 林线冷杉种群的时间-空间动态

图4 林线冷杉种群密度的时间动态

图5 阴阳坡峨眉冷杉树线、林线海拔高度比较
4 讨论
作为一种典型的生态过渡带,高山林线指示着森林分布的上界,林线树种对分布边界极端环境的形态、生理和生态适应一直是引人关注的热点问题[27- 28]。已有大量研究表明气候变暖会导致林线向上扩张,植被带上移[11,17- 20],但同时有研究表明气候变暖后林线无明显变化[21- 22], 或者其变化不是由气候变暖引起的[23- 24]。在本研究中,发现雅家埂阴阳坡树线的海拔高度在过去100a间并无明显变化,尽管这与Penuelas[11]等人的研究结果存在差异,但却与Liang[22]等人的研究结果是一致的。在Penuelas等人的研究中,其林线树种为水青冈属植物(欧洲水青冈),而在Liang等人和本研究中,林线树种均为冷杉属植物(分别为急尖长苞冷杉和峨眉冷杉),这两个属一个为落叶阔叶植物,一个为常绿针叶植物, 不同的植物属性可能导致林线树种对林线附近垂直环境梯度的响应格局并不一致,进而导致这种差异的出现。此外,林线植被的异质格局是多种环境因素共同作用的结果[27,29- 31],离散的极端事件(如干扰)对当前林线的位置和群落结构起着特别重要的作用[27,32- 33],如Van[24]等人就曾指出,在气候变暖过程中,干扰对树线的移动有重要影响,它不仅能够决定树线何时向上爬升,而且能决定树线是否向上爬升,如瑞典北部Tornetrask地区林线附近幼树的更新和树线的爬升就与该区驯鹿种群周期性地变小密切相关。在本研究中,贡嘎山东坡雅家埂地区存在牧民放牧,林线区域偶尔有牦牛和山羊经过(受地理位置的影响,阳坡放牧强度大于阴坡),林线峨眉冷杉种群的更新和生长可能会受到放牧的干扰,从而可能在一定程度上限制树线向上爬升。因此,在运用气候驱动模型来预测未来树线位置或者用树线位置来进行生物气候监测时应特别谨慎,在那些不是由气候因素决定树线位置的区域更是如此。此外,还发现阴阳坡林线附近峨眉冷杉的种群密度在最近50a中有显著地升高(图4)。尽管有研究报道林线附近幼树更新在最近几十年有所减少[34- 35],但越来越多的研究表明林线附近乔木的种群密度会对温度的升高做出迅速的响应[22,36],尤其是在最近100a的小时间尺度上,林线海拔位置的变化有可能不是很明显,但林线树木密度却有显著地增加[35,37- 38]。
温度通常被认为是决定林线位置的最重要的环境因子,Körner and Paulsen[13]认为生长季平均土壤(10 cm深)温度(6.7±0.8)℃是全球林线分布的控制因子[39],王襄平等[40]也认为限制中国林线高度的气候因子是生长季节温度8.2 ℃。根据上述观点,由于阳坡所接受太阳辐射大于阴坡,导致阳坡气温高于阴坡,阳坡的林线高度应比阴坡高。然而,在本研究中,发现阴坡林线和树线的海拔高度显著高于阳坡(图4),这表明除气候因素外,其它因素也限制了林线和树线位置的变化。除温度外,高山地区的环境胁迫、干扰、繁殖更新障碍以及植物的碳平衡失调都可能影响林线的海拔高度[39]。雅家埂地区阳坡平均坡度为39.67°,阴坡平均坡度为30.33°,阳坡较陡的坡度及相对较高的放牧强度可能对林线附近峨眉冷杉种子的传播、幼苗定植及生长产生了一定的不利影响;此外,阳坡降水量低于阴坡也可能是其林线和树线高度低于阴坡的一个重要原因。因此,雅家埂地区目前的林线格局应该是温度、降水、坡度以及干扰等多种因素共同作用的结果。
除树线、林线的海拔高度存在明显的坡向差异外,阴阳坡林线附近冷杉立木的胸径-树龄关系也存在一定的坡向分异:如图1所示,阴阳坡林线附近冷杉立木的胸径-树龄指数方程分别为y=26.14e0.046x(R2=0.837)和y=34.66e0.037x(R2=0.881),阳坡峨眉冷杉在早期的生长速率大于阴坡,但这种差异随树龄的增加逐渐缩小,通过求解方程可知,在树龄低于110a(胸径小于31.35 cm)之前,阳坡峨眉冷杉生长速率高于阴坡,但超过110a,阴坡峨眉冷杉的生长速率逐渐大于阳坡。Körner[41]总结了不同环境因子对高山植物光合作用的影响,发现光照是高山林线光合作用的主要限制因子。阳坡峨眉冷杉早期的生长速率高于阴坡,可能是由于阳坡相对较高的气温和太阳辐射导致其有更高的光合速率所致。然而,随着树龄的增加,植物叶片的光合氮利用效率会逐渐降低,如Ishida 等[42]发现Macarangagigantea(Reichb.f.和Zoll.)Muell幼苗的光合氮利用效率效率是成树的1.6倍,考虑到植物体内氮利用效率和水分利用效率之间存在某种平衡关系[43]而阴坡降水量大于阳坡,阴坡峨眉冷杉可能会通过降低水分利用效率来提高养分利用效率,从而使其生长速率逐渐接近甚至超过阳坡的峨眉冷杉。
5 结论
尽管贡嘎山东坡雅家埂林线附近峨眉冷杉种群的密度在过去100a间(主要是最近50a)密度有显著升高,但树线位置并未发生显著的变化。此外,阴阳坡林线和树线格局存在显著的坡向差异:阴坡林线和树线的海拔高度显著高于阳坡;阳坡峨眉冷杉的生长速率在早期高于阴坡但在后期低于阴坡。然而,由于缺乏直接的气象、土壤养分、植物养分利用效率和水分利用效率的数据,有关上述阴阳坡差异成因的探讨仅仅是一个初步的推测,不能过度解读。
:
[1] IPCC. Climate change 2007: The physical science basis. Contribution of Working Group I to the fourth assessment report of the Intergovernmental Panel on Climate Change // Solomon S, Qin D, Manning M, Chen Z, Marquis M, Averyt K B, Tignor M, Miller H L. Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA, 2007.
[2] Saxe H, Cannell M G R, Johnsen B, Ryan M G, Vourlitis G. Tree and forest functioning in response to global warming. New Phytologist, 2001, 149(3): 369- 399.
[3] Kellomäki S, Wang K Y. Growth and resource use of birch seedlings under elevated carbon dioxide and temperature. Annals of Botany, 2001, 87(5): 669- 682.
[4] Vu J C V, Newman Y C, Allen L H Jr, Gallo-Meagher M, Zhang M Q. Photosynthetic acclimation of young sweet orange trees to elevated growth CO2and temperature. Journal of Plant Physiology, 2002, 159(2): 147- 157.
[5] Niu S L, Li Z X, Xia J Y, Han Y, Wu M Y, Wan S Q. Climatic warming changes plant photosynthesis and its temperature dependence in a temperate steppe of northern China. Environmental and Experimental Botany, 2008, 63(1/3): 91- 101.
[6] Xu X, Yang F, Xiao X W, Zhang S, Korpelainen H, Li C Y. Sex-specific responses ofPopuluscathayanato drought and elevated temperatures. Plant Cell and Environment, 2008, 31(6): 850- 860.
[7] Yin H J, Liu Q, Lai T. Warming effects on growth and physiology in the seedlings of the two conifersPiceaasperataandAbiesfaxonianaunder two contrasting light conditions. Ecological Research, 2008, 23(2): 459- 469.
[8] Way D A, Oren R. Differential responses to changes in growth temperature between trees from different functional groups and biomes: a review and synthesis of data. Tree Physiology, 2010, 30(6): 669- 688.
[9] Loik M E, Still C J, Huxman T E, Harte J. In situ photosynthetic freezing tolerance for plants exposed to a global warming manipulation in the Rocky Mountains, Colorado, USA. New Phytologist, 2004, 162(2): 331- 341.
[10] Aerts R, Cornelissen J H C, Dorrepaal E. Plant performance in a warmer world: general responses of plants from cold, northern biomes and the importance of winter and spring events. Plant Ecology, 2006, 182(1/2): 65- 77.
[11] Penuelas J, Boada M. A global change-induced biome shift in the Montseny mountains (NE Spain). Global Change Biology, 2003, 9(2): 131- 140.
[12] Wilson R J, Gutiérrez D, Gutiérrez J, Martínez D, Agudo R, Monserrat V J. Changes to the elevational limits and extent of species ranges associated with climate change. Ecology Letters, 2005, 8(11): 1138- 1146.
[13] Körner C, Paulsen J. A world-wide study of high altitude treeline temperatures. Journal of Biogeography, 2004, 31(5): 713- 732.
[14] Holtmeier F K, Broll G. Treeline advance-driving processes and adverse factors. Landscape, 2007, 1: 1- 33.
[15] Harsch M A, Hulme P E, McGlone M S, Duncan R P. Are treelines advancing? A global meta-analysis of treeline response to climate warming. Ecology Letters, 2009, 12(10): 1040- 1049.
[16] Wang X D, Liu H Q. The dynamics response ofBetulaermaniipopulation and climate change on different slopes aspect of north slope, Changbai Mountains. Scientia Geographica Sinica, 2012, 32(2): 199- 206.
[17] Sanz-Elorza M, Dana E D, Gonzalez A. Changes in the high-mountain vegetation of the central Iberian peninsula as a probable sign of global warming. Annals of Botany, 2003, 92(1): 273- 280.
[18] Danby R K, Hik D S. Variability, contingency and rapid change in recent subarctic alpine tree line dynamics. Journal of Ecology, 2007, 95(2): 352- 363.
[19] Kirdyanov A V, Hagedorn F, Knorre A A, Fedotova E V, Vaganov E A, Naurzbaev M M, Moiseev P A, Rigling A. 20th century tree-line advance and vegetation changes along an altitudinal transect in the Putorana Mountains, northern Siberia. Boreas, 2012, 41(1): 56- 67.
[20] Dai J H, Pan Y, Cui H T, Tang Z Y, Liu H Y, Cao Y L. Impacts of climate change on alpine vegetation on Wutai Mountains. Quaternary Sciences, 2005, 25(2): 216- 223.
[21] Cullen L E, Stewart G H, Duncan R P, Palmer J G. Disturbance and climate warming influences on New Zealand Nothofagus tree-line population dynamics. Journal of Ecology, 2001, 89(6): 1061- 1071.
[22] Liang E Y, Wang Y F, Eckstein D, Luo T X. Little change in the fir tree-line position on the southeastern Tibetan Plateau after 200 years of warming. New Phytologist, 2011, 190(3): 760- 769.
[23] Batllori E, Gutiérrez E. Regional tree line dynamics in response to global change in the Pyrenees. Journal of Ecology, 2008, 96(6): 1275- 1288.
[24] Van B R, Haneca K, Hoogesteger J, Jonasson C, De Dapper M, Callaghan T V. A century of tree line changes in sub-Arctic Sweden shows local and regional variability and only a minor influence of 20th century climate warming. Journal of Biogeography, 2011, 38(5): 907- 921.
[25] Baker B B, Moseley R K. Advancing treeline and retreating glaciers: implications for conservation in Yunnan, P R China. Arctic, Antarctic and Alpine Research, 2007, 39(2): 200- 209.
[26] Yang L D, Wang G X, Yang Y, Yang Y. Responses of leaf functional traits and nitrogen and phosphorus stoichiometry inAbiesfabiriseedlings in Gongga Mountain to simulated nitrogen deposition. Chinese Journal of Ecology, 2012, 31(1): 44- 50.
[27] Shen Z H, Fang J Y, Liu Z L, Wu J. Structure and dynamics ofAbiesfabripopulation near the alpine timberline in Hailuo clough of Gongga Mountain. Acta Botanica Sinica, 2001, 43(12): 1288- 1293.
[28] Dai J H, Cui H T. A review on the studies of alpine timberline. Acta Geographica Sinica, 1999, 19(3): 243- 249.
[29] Cairns D M, Malanson G P. Environmental variables influencing the carbon balance at the alpine treeline: a modeling approach. Journal of Vegetation Science, 1998, 9(5): 679- 692.
[30] Cairns D M. Multi-scale analysis of soil nutrients at alpine treeline in Glacier National Park, Montana. Physical Geography, 1999, 20: 256- 271.
[31] Moir W H, Rochelle S G, Schoettle A W. Micro-scale patterns of tree establishment near upper treeline, Snowy Range, Wyoming, USA. Arctic Antarctic and Alpine Research, 1999, 31(4): 379- 388.
[32] Shankman D. Tree regeneration following fire as evidence of timberline stability in the Colorado Front Range, U. S. A. Arctic Antarctic and Alpine Research, 1984, 16(4): 413- 417.
[33] Szeicz J M, MacDonald G M. Recent white spruce dynamics at the subarctic alpine treeline of north-western Canada. Journal of Ecology, 1995, 83(5): 873- 885.
[34] Villalba R, Veblen T T. Regional patterns of tree population age structures in northern Patagonia: climatic and disturbance influences. Journal of Ecology, 1997, 85(2): 113- 124.
[35] Wang T, Zhang Q B, Ma K P. Treeline dynamics in relation to climatic variability in the central Tianshan Mountains, northwestern China. Global Ecology and Biogeography, 2006, 15(4): 406- 415.
[36] Camarero J J, Gutiérrez E. Pace and pattern of recent treeline dynamics: response of ecotones to climatic variability in the Spanish Pyrenees. Climatic Change, 2004, 63(1/2): 181- 200.
[37] Klasner F L, Fagre D B. A half century of change in alpine treeline patterns at Glacier National Park, Montana, U. S. A. Arctic Antarctic and Alpine Research, 2002, 34(1): 49- 56.
[38] Wang X C, Zhou X F, Sun Z H. Research advances in the relationship between alpine timberline and climate change. Chinese Journal of Ecology, 2005, 24(3): 301- 305.
[39] Li M H, Kräuchi N. The state of knowledge on alpine treeline and suggestions for future research. Journal of Sichuan Forestry Science and Technology, 2005, 26(4): 36- 42.
[40] Wang X P, Zhang L, Fang J Y. Geographical differences in alpine timberline and its climatic interpretation in China. Acta Geographica Sinica, 2004, 59(6): 871- 879.
[41] Körner C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems. Berlin, Heidelberg: Springer-Verlag, 1999: 171- 200.
[42] Ishida A, Yazaki K, Hoe A L. Ontogenetic transition of leaf physiology and anatomy from seedlings to mature trees of a rain forest pioneer tree, Macaranga gigantea. Tree Physiology, 2005, 25(5): 513- 522.
[43] Li C Y, Wu C C, Duan B L, Korpelainen H, Luukkanen O. Age-related nutrient content and carbon isotope composition in the leaves and branches ofQuercusaquifolioidesalong an altitudinal gradient. Trees-Structure and Function, 2009, 23(5): 1109- 1121.
参考文献:
[16] 王晓东, 刘惠清. 长白山北坡林线岳桦种群动态对气候变化响应的坡向分异. 地理科学, 2012, 32(2): 199- 206.
[20] 戴君虎, 潘嫄, 崔海亭, 唐志尧, 刘鸿雁, 曹燕丽. 五台山高山带植被对气候变化的响应. 第四纪研究, 2005, 25(2): 216- 223.
[26] 羊留冬, 王根绪, 杨阳, 杨燕. 峨眉冷杉幼苗叶片功能特征及其N、P化学计量比对模拟大器氮沉降的响应. 生态学杂志, 2012, 31(1): 44- 50.
[27] 沈泽昊, 方精云, 刘增力, 伍杰. 贡嘎山海螺沟林线附近峨眉冷杉种群的结构与动态. 植物学报, 2001, 43(12): 1288- 1293.
[28] 戴君虎, 崔海亭. 国内外高山林线研究综述. 地理科学, 1999, 19(3): 243- 249.
[38] 王晓春, 周晓峰, 孙志虎. 高山林线与气候变化关系研究进展. 生态学杂志, 2005, 24(3): 301- 305.
[39] 李迈和, Kräuchi N. 全球高山林线研究现状与发展方向. 四川林业科技, 2005, 26(4): 36- 42.
[40] 王襄平, 张玲, 方精云. 中国高山林线的分布高度与气候的关系. 地理学报, 2004, 59(6): 871- 879.
猜你喜欢
广西植物(2022年2期)2022-03-16
山西林业科技(2022年4期)2022-03-12
南方农业(2022年23期)2022-02-14
思维与智慧·下半月(2021年7期)2021-08-13
防护林科技(2019年7期)2019-08-19
花火A(2019年1期)2019-03-22
现代农村科技(2018年11期)2018-11-15
现代养生·上半月(2017年7期)2017-07-07
厦门航空(2017年11期)2017-01-29
天津诗人(2016年4期)2016-11-26
