
力竭运动后小鼠前脑皮层和海马神经元功能的变化及其机制
2014-09-20 07:21,
山东体育科技 2014年5期
,
(1. 山东省体育科学研究中心, 山东 济南 250102;2.曲阜师范大学 生命科学学院, 山东 曲阜 273165)
力竭运动后小鼠前脑皮层和海马神经元功能的变化及其机制
蔡成法1,李亚2
(1. 山东省体育科学研究中心, 山东 济南 250102;2.曲阜师范大学 生命科学学院, 山东 曲阜 273165)
目的:研究力竭游泳运动对小鼠前脑皮层和海马神经元功能的影响及其作用机制。方法:采用力竭游泳运动小鼠模型,检测反复(4周)力竭游泳运动后即刻(0小时)、12小时、24小时和1周小鼠前脑皮层、海马突触体膜流动性和突触体内游离Ca2+浓度的变化,通过Morris水迷宫测试小鼠空间学习记忆能力。结果:反复力竭游泳运动后,与正常对照组小鼠比较,力竭运动组在12 h和24 h小鼠的逃避潜伏期(训练周期1)明显增加,第一象限停留时间明显减少,其空间学习记忆能力显著降低(Plt;0.01,Plt;0.05);力竭运动组小鼠前脑皮层和海马突触体膜流动性在0 h、12 h显著降低(Plt;0.01,Plt;0.05) ;前脑皮层和海马突触体内游离Ca2+浓度在0 h、12 h和24 h显著增加 (Plt;0.05,Plt;0.01),力竭运动后1周前脑皮层和海马突触体内游离Ca2+浓度明显恢复(Plt;0.05)。结论:力竭游泳运动所致小鼠空间学习记忆功能损伤以及运动性中枢疲劳的产生和恢复可能与突触体膜流动性和突触体内游离Ca2+浓度的变化密切相关。
力竭运动;膜流动性;Ca2+浓度;空间学习记忆;海马;前脑皮层
力竭性运动表现为动物或人进行运动直到完全不能运动为止[1]。由于运动负荷过大,超过机体的承受能力,可引起机体多器官功能紊乱,尤其是导致中枢神经系统运动性疲劳,造成脑功能损伤。前脑皮层和海马是脑内运动的调控、学习记忆、神经内分泌及认知功能的重要区域,也是最易受运动应激损伤的脑区[2]。突触体膜的结构和功能变化可引起突触处信息传递效能发生改变,尤其是突触体膜流动性的改变将影响膜的离子通透性、跨膜物质转运、胞内信号转导等。在脑内神经元受损及脑的老化过程中,膜流动性也显示相应的改变[3]。本研究建立4周反复力竭游泳运动小鼠模型,测定力竭游泳运动和恢复期小鼠Morris 水迷宫空间学习记忆能力,检测小鼠前脑皮层和海马突触体膜流动性、突触体内游离Ca2+浓度的变化,探讨力竭游泳运动对小鼠空间学习记忆功能影响,以及运动性中枢疲劳产生与恢复的突触机制。
1 材料与方法
1.1 实验动物和分组
3月龄健康昆明品系小白鼠,雌雄各半,山东鲁抗医药股份有限公司动物实验中心提供。适应性喂养一周后随机分为:正常对照组,反复性力竭游泳运动组。
1.2 力竭游泳运动小鼠模型的建立
建立4周反复力竭游泳运动小鼠模型。实验用小鼠接受连续3d的适应性游泳训练,自由游泳每天一次,持续时间分别为15 min、30 min、45 min逐日递增。水温30±2 ℃。4周反复力竭游泳运动小鼠,每天游泳训练一次,每次游泳训练均至力竭,每周5天;为避免力竭训练组大鼠在训练过程中产生适应现象,在训练的后期采用负重(体重的2%)的方法增加游泳训练的强度,每次训练均要求达到力竭状态。力竭游泳运动标准为:1)游泳动作明显失调,不能再坚持;2)沉入水底超过3 s不能回到水面。停止游泳运动训练,及时捞起,吹干毛发,放回饲养笼中休息。力竭游泳运动后即刻(0小时)、12小时、24小时和1周,检测小鼠前脑皮层和海马突触体膜流动性和突触体内游离Ca2+浓度。
1.3 Morris水迷宫学习记忆能力检测
力竭游泳运动后12小时、24小时和1周,对小鼠实施Morris水迷宫实验。Morris 水迷宫包含定位航行实验(Place navigation test)和空间搜索实验(Spatial probe test)。定位航行实验历时2 d,每只小鼠每天接受8次找平台训练,分2个训练周期(上午和下午),每个训练周期4次,每次训练2 min。记录小鼠寻找到隐藏在水下平台时间即逃避潜伏期,检测小鼠在水迷宫中的学习能力;第3 d撤除平台进行空间搜索实验,从第三象限将小鼠放入水中,记录120 s内小鼠在4个象限游泳(停留)时间,检测小鼠对原平台(目标象限)记忆保持能力。
1.4 突触体膜流动性和突触体内游离Ca2+浓度的测定
将各组小鼠断头处死,在冰台上迅速分离两侧前脑皮层和海马并称重,加入10倍体积的0.32 M蔗糖溶液(内含10 mM Hepes,pH 7.4,W:V = 1:10)进行匀浆。按差速蔗糖梯度离心法分离制备突触体,最后用人工脑脊液(ACSF)缓慢悬浮,得到突触体悬液,所有操作均在0~4 ℃进行。分别取前脑皮层和海马突触体悬液,用ACSF调整蛋白浓度为0.3 mg/ml,与等体积的荧光探针DPH(Fluka公司产品,终浓度2×10-6M)稀释液,置于37 ℃恒温孵育30 min。用荧光偏振法检测突触体膜的荧光偏振度(P),测定温度37 ℃。用膜各向异性(anisotropy)r值表示突触体膜的流动性大小,r值越小,膜流动性越大。取前脑皮层和海马突触体悬液,用ACSF调整蛋白浓度为1.5~2.5 mg/ml,加入Fura 2-AM(Sigma公司产品,终浓度:5 μ M), 37 ℃恒温孵育30 min后,于11 000 ×g离心10 min,取沉淀,用ACSF悬浮,于0~4 ℃下保存直至荧光测定。采用双波长测定,选择Fura-2与Ca2+结合和游离的激发波长340 nm和380 nm,发射波长为509 nm。在37 ℃条件下,测定其荧光强度及荧光比值。计算突触体内钙离子浓度。
1.5 统计学分析
所有数据以均数±标准差(± s)表示,采用SPSS 13.0进行统计学分析。采用单因素方差分析(one-way ANOVA),对各象限游泳时间及力竭运动后各时间点数据进行多重比较,逃避潜伏期采用重复测量方差分析,行LSD-t检验或非配对t检验。Plt;0.05表示具有统计学显著性差异。
2 结果
2.1 小鼠定位航行学习能力的变化
统计分析结果表明(表1),正常对照组与力竭运动后各组小鼠逃避潜伏期均显著减少(Pgt;0.05,Plt;0.01)。与正常对照组比较,力竭运动后12 h、24 h在2天的四个训练周期,都有显著性差异(Plt;0.01)。而力竭运动后1周,小鼠在第三和第四训练周期时逃避潜伏期显著增多(Plt;0.05)。结果说明小鼠反复力竭运动后12 h、24 h空间学习能力下降,力竭运动后1周小鼠的空间学习能力有所恢复。
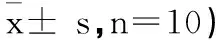
表1 各组小鼠的逃避潜伏期(秒,
2.2 小鼠空间搜索记忆能力的变化
结果显示,各组小鼠均在第一象限的停留时间最长(表2)。正常对照组力竭运动后1周小鼠在第一象限的停留时间较其它三个象限的停留时间明显增多(Plt;0.05,Plt;0.01);与正常对照组比较,力竭运动后12h和24h小鼠在第一象限的停留时间明显减少(Plt;0.05,Plt;0.01),结果表明,力竭运动后12h、24h小鼠的空间记忆保持能力明显减弱。
表2 小鼠慢性应激后在四个象限的游泳时间(秒,
2.3 小鼠突触体膜流动性和突触体内游离Ca2+浓度的变化
力竭运动后,力竭运动组小鼠前脑皮层和海马突触体膜的r值显著增加(表 3),表明力竭游泳运动引起突触体膜流动性显著降低。与正常对照组比较,力竭运动后即刻(0小时)、12小时,小鼠前脑皮层和海马突触体膜流动性明显降低(Plt;0.05,Plt;0.01)。力竭运动后24小时、1周小鼠前脑皮层和海马突触体膜的r值逐渐减小;与力竭运动后即刻比较,在力竭游泳后1周小鼠海马突触体膜流动性明显增加(Plt;0.05)。与正常对照组相比,力竭运动后即刻、12小时,小鼠前脑皮层和海马突触体内游离Ca2+浓度显著升高(Plt;0.05,Plt;0.01)。力竭运动后24小时、1周小鼠前脑皮层和海马突触体内游离Ca2+浓度逐渐降低;与力竭运动后即刻比较,力竭运动后1周小鼠前脑皮层和海马突触体内游离Ca2+浓度有统计学显著差异(Plt;0.05)。
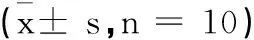
表3 小鼠突触体膜流动性和游离Ca2+浓度
3 讨论
已有研究报道,过度运动后可引起大鼠大脑皮质的超微结构发生明显改变,而力竭运动后的一定时间内,大脑皮质结构得损伤可持续存在,可以推测脑组织结构出现异常变化,引起大脑的调节功能减退,导致运动性中枢疲劳的产生[4]。研究表明,6周的力竭游泳训练可引起动物海马神经元损伤,而脑源性神经营养因子(BDNF)可能参与了力竭运动后脑损伤的保护和修复过程[5]。
大量运动实践表明,长期大运动量训练诱导机体运动机能下降或运动性疲劳发生时,机体应激反应增强,血液中皮质醇维持在高水平,而海马对于介导应激反应,具有重要的调节作用[6]。研究表明,慢性应激对海马介导的记忆有持久性损伤作用,这可能是通过改变海马突触可塑性而影响脑海马区的功能活动[7]。在中枢神经系统,应激因素除对海马有影响外,对其它脑区,如前额叶、杏仁核等也有作用[8]。已经明确,适宜的膜流动性是维持生物膜正常功能的必须条件。脑内海马神经细胞内Ca2+平衡失调,亦可导致海马神经细胞受损,引起动物空间学习记忆功能减退[9]。我们先前的研究表明,慢性应激后,小鼠前脑皮层和海马突触体膜流动性显著降低,而突触体内游离Ca2+浓度的显著增加[10],导致小鼠前脑皮层和海马功能受损。因此,可以推测,在本研究中反复力竭游泳运动后,突触体内游离Ca2+浓度显著增加,而突触体膜流动性明显降低,引起神经元突触传递效能降低,导致前脑皮层和海马神经元功能受损,空间学习记忆受损。力竭运动引起的这些变化可能与小鼠运动性中枢疲劳产生和恢复高度相关。
4 结论
4.1 力竭运动可损伤小鼠前脑皮层和海马神经元功能,继而降低其空间学习记忆功能。
4.2 突触体膜流动性和突触体内游离Ca2+浓度的变化可能是力竭运动所导致小鼠运动性中枢疲劳产生和恢复的重要机制之一。
[1]曹阳,成晓龙,范琳琳,等.力竭运动对大鼠下丘脑室旁核内NF-κB、Bcl-2、Bax表达的影响[J].中国运动医学杂志,2012,31(4):315-319.
[2]Lupien SJ, McEwen BS, Gunnar MR,etal.Effects of stress throughout the lifespan on the brain, behaviour and cognition [J]. Nat Rev Neurosci, 2009, 10(6):434-445.
[3]Li Y, Wang JJ, Cai JX. Aniracetam restores the effects of amyloid-b protein or ageing on membrane fluidity and intracellular calcium concentration in mice synaptosomes[J]. J Neural Transm,2007, 114(11): 1407-1411.
[4]王栋梁,张卫国,宋慧芳,等. 1 周力竭运动后大鼠额叶运动脑区NF-κB 表达变化[J].中国运动医学杂志, 2010, 29(3):326-328.
[5]张鹏云,和荣丽,何生,等.力竭运动后大鼠海马NF-κB、BDNF 的表达变化[J].神经解剖学杂志, 2012,28(1):63-68.
[6]Hill MN, Hunter RG, McEwen BS. Chronic stress differentially regulates cannabinoid CB1 receptor binding in distinct hippocampal subfields[J].Eur J Pharmacol, 2009, 614(1-3): 66-69.
[7]Bruel-Jungerman E, Rampon C, Laroche S. Adult hippocampal neurogenesis, synaptic plasticity and memory: facts and hypotheses[J]. Rev Neurosci, 2007, 18(2):93-114.
[8]Roozendaal B, McEwen BS, Chattarji S. Stress, memory and the amygdale [J]. Nat Rev Neurosci, 2009, 10(6):423-433.
[9]李亚,孔宏,宋倩,等.慢性应激对大鼠空间学习记忆及海马和前脑皮层突触体膜流动性的影响[J].心理学报, 2010, 42(2):235-240.
[10]李亚,陈亚静,史建勋,张冠雄. 慢性应激对小鼠学习记忆功能影响及其突触作用[J].中国公共卫生,2012.28(12):1602-1604.
Effects of exhausted exercise on the prefrontal cortex and hippocampus function in mice and its mechanism
CAI Cheng-fa1,LI Ya2
(1. Shandong Research Center of Sports Sciences, Jinan 250102, Shandong, China. 2.College of Life Sciences, Qufu Normal University, Qufu 273165, Shandong, China)
Objective:To study the effect of exhausted exercise on the prefrontal cortex (PFC) and hippocampus (HP) function and its mechanism.Methods:The repeatedly (4 weeks) exhausted exercise model mice were applied. The spatial learning-memory abilities of mice were tested, using Morris water maze task. The changes of synaptosomal membrane fluidity and [Ca2+] i of the PFC and HP in exhausted exercise mice brain were examined, at 0 h (immediately),12 h,24 h, respectively.Results:After repeatedly exhausted exercise, the escape latency was significantly increased and the swimming time of first quadrants were significantly decreased at 12 h, 24 h in exhaustive exercise group mice(Plt;0.01,Plt;0.05). Compared with control mice, the ability of spatial learning and memory of the exhaustive exercise group mice were significantly reduced. The membrane fluidity of synaptosomes in PFC and HP of exhaustive exercise group mice were significantly decreased at 0 h and 12 h (Plt;0.05,Plt;0.01), compared with control group mice. The synaptosomal[Ca2+]i in PFC and HP were significantly increased at 0 h, 12 h and 24 h(Plt;0.05,Plt;0.01) in exhaustive exercise group mice. The [Ca2+]i in PFC and HP at 1 week were significant reduced than the exhaustive exercise 0 h group mice(Plt;0.05), respectively.Conclusion:The generation and recovery of exercise-induced central fatigue and the damage of spatial learning and memory function in mice after exhausted exercise which may be nearly related to the changes of membrane fluidity and [Ca2+]i of synaptosomes.
exhausted exercise; membrane fluidity; free calcium concentrations; spatial learning-memory; hippocampus; prefrontal cortex
2014-05-06
山东省优秀中青年科学家科研奖励基金项目(编号:BS2012SW021),山东省自然科学基金(编号:ZR2009DL009)。
蔡成法(1974- ),男,博士,助理研究员,研究方向运动生理生化。
A
1009-9840(2014)05-0080-03
猜你喜欢
中华实验眼科杂志(2022年6期)2022-11-15
实用手外科杂志(2022年2期)2022-08-31
作文周刊·小学二年级版(2022年20期)2022-05-05
科学导报·学术(2020年29期)2020-10-21
美与时代·美术学刊(2019年9期)2019-11-29
首都体育学院学报(2019年5期)2019-10-18
中国当代医药(2017年17期)2017-07-25
支部建设(2016年18期)2016-11-28
科技知识动漫(2016年6期)2016-06-24
外语教学理论与实践(2014年2期)2014-06-21
