
联觉的大脑网络激活*
2014-10-24 10:13李佳源赵伶俐
华东师范大学学报(教育科学版) 2014年2期
李佳源 赵伶俐
(1.西华师范大学管理学院,南充637009;2.西南大学教育学部,重庆400715)
联觉是一种感觉引起另一种感觉的心理现象(黄希庭,2007;Simner,2012),以往不少文献认为高尔顿(Galton,1880)最早发现联觉现象,但事实上Georg T.L.Sachs在1812年就对联觉(synaesthesia)现象进行了第一次描述(Simner,2012)。20世纪初,联觉成为心理学中一个令人感兴趣的问题,但当时的研究者未能成功地定义一个客观体系来描述此现象的特征,因此之后对这个问题的兴趣逐渐消退。然而伴随着认知革命的兴起和人类神经科学新的实验技术的迅速崛起,对联觉的兴趣作为科学研究的显性问题得到复兴。近年来,基础心理学、认知心理学竭力与神经科学研究前沿接轨,将行为实验与神经成像技术结合,致力于联觉的全脑研究。利用神经成像技术研究的常见联觉类型是颜色联觉,比如字母—颜色联觉、数字—颜色联觉等。本文以颜色联觉为基础,展现和分析有关联觉的全脑激活的新进展及其相关问题,揭示联觉研究中有待进一步研究的问题和方向。
一 大脑激活:从特定的大脑区域到大脑区域网络
大脑皮层的V4区一般被认为是处理颜色的专属区域,不少研究已经发现在联觉颜色的体验期间,V4区(颜色区域)被激活(Eagleman,2012)。然而除了V4区之外,目前也有研究发现联觉颜色体验期间被激活的区域超出了V4区(颜色区域),那么除了V4区外,到底还有哪些大脑区域与联觉颜色体验相关联呢?PET(正电子发射断层扫描)和fMRI(功能磁共振成像)是被使用来研究联觉脑机制的两种常用技术,它们具有很高的空间分辨率,能通过血氧量和局部脑血流变化指标来反映大脑皮层活动的情况。图1是目前研究者使用fMRI或PET来测量对联觉颜色作出反应的全脑激活情况,这些研究显示了大脑中不同区域的激活,在目前的实验报告中有五个区域的激活出现相对集中。
第一个区域是在枕颞叶皮层中的双侧激活,这在全脑分析中得到了充分显现(左:Laeng et al.2011;Nunn et al.,2002;Steven et al.,2006;右:Laeng et al.,2011;Rouw & Scholte,2007;Weiss et al.,2001)。在图1A中可以看到,增加激活的位置不仅仅局限于V4区,但出现激活位置不局限于V4区的情况不是由于实验研究中研究方法、实验装置或联觉者类型差异造成的。Hubbard等人(2005)使用皮层映射的研究方法来测量颜色联觉在大脑中的激活情况,皮层映射能够改进信号噪声比,能够提高对联觉颜色激活的敏感性。Laeng等人(2011)设计了一种更为巧妙的实验装置,他们提供给联觉者具有色彩的字母,并控制字母的字体颜色与它引起的联觉颜色之间的颜色距离,它能够通过操作结合体验(联觉的和字体的颜色)的性质,而不是主要地对比联觉颜色体验的出现和缺失,来更为敏感地测量被试的联觉体验。Novich等人(2011)用了接近四年的时间测试了更大量的联觉者群体(19133名联觉者),根据联觉特征得到了五种不同的联觉组,而这些不同的联觉组各自具有独立的表征,意味着可能有不同的神经基础。前两个实验在以往实验研究的基础上对研究方法和实验装置进行了改进,后一个实验则测量了更多类型的联觉者,然而他们的研究结果都显示了在V4区域,被真实颜色激活的,也能被联觉颜色激活。更为重要的是,他们的研究也发现联觉颜色体验激活的位置确实不局限于V4区,而是涉及了大脑枕颞叶皮层的更广泛的区域。
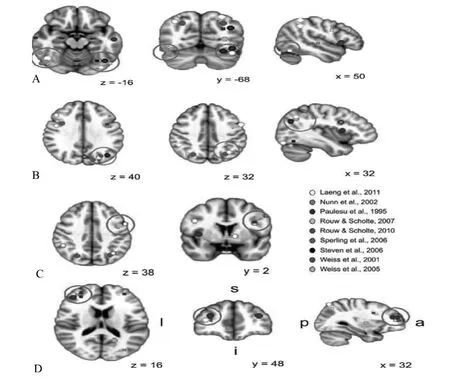
图1 联觉体验期间的全脑激活情况(Rouw et al.,2011)(图1显示了在言语—颜色联觉期间的大脑激活情况,在9个不同研究中获得的大脑激活区域坐标被给出,A、B、C、D分别代表了在9个研究中发现的被激活的大脑位置)
第二个区域是在顶叶皮层的集群激活,几乎完全位于后顶叶皮层。后顶叶皮层包括顶上小叶和顶下小叶,全脑研究发现在左顶上小叶(Laeng et al.,2011;Weiss et al.,2005)和右顶上小叶(Laeng etal.,2011;Paulesu et al.,1995;Weiss et al.,2005)中有对应于联觉颜色体验的增加激活;而共同激活的区域是顶下小叶,在顶下小叶中所有的激活位置要么在内沟附近,要么在角回中,即一些集群激活位于顶下小叶中较靠前和靠上的位置(Laeng et al.2011,Nunn et al.,2002,Weiss et al.,2005),其他的则位于顶下小叶中靠后和靠下的位置 (Laeng et al.,2011;Rouw&Scholte,2010;Steven et al.,2006)(见图1B)。顶下小叶这个区域的激活在随后的TMS(经颅磁刺激)研究中也被证实,但不清楚的是:为什么TMS研究仅仅发现了在右下顶叶区域联觉颜色体验的激活,然而fMRI或PET却发现主要是在左下顶叶区域。
后顶叶皮层在视觉特征捆绑中起到重要作用,而联觉将真实出现的刺激物与没有真实出现的伴随体验捆绑在一起,因此后顶叶皮层靠上的区域(顶上小叶)和后顶叶皮层靠下的区域(顶下小叶)可能与作为联觉的一个内在部分的捆绑有关,是联觉颜色体验期间被激活的一个重要备选区域。
第三个区域是双侧脑岛和岛盖(Nunn et al.,2002;Paulesu et al.,1995;Sperlinget al.,2006)。脑岛激活是与从一个特定的外在刺激到一个不同的内在刺激的转化过程相关,也可能是与常常伴随联觉体验的情绪特质有关,因为联觉者常常报道某种情绪属于一种联觉体验。如果这种解释正确,那么在不同类型的联觉体验中都能出现增加的脑岛激活。
第四个区域是中央前回。五个研究发现了在左侧中央前回的激活(Laeng et al.2011;Paulesu et al.,1995;Nunn et al.,2002;Rouw& Scholte,2010;Weiss et al.,2005),然而三个研究发现了在右侧中央前回的激活(Laeng et al.,2011;Paulesu et al.,1995;Rouw&Scholte,2010)(见图1C)。中央前回是处理感知外在世界以及在外在世界中行为的专属大脑区域,这个区域的增加激活说明了在联觉中更广泛的大脑区域网络被涉及(视觉皮层、在脑岛中和附近的激活,以及中央激活)。
第五个区域是额叶。9个研究中有3个研究发现了接近于右背外侧前额叶皮层位置的激活(Laeng et al.,2011;Paulesu et al.,1995;Sperling et al.,2006)(见图1D),这个区域的激活可能反映了联觉的一个重要方面,即认知控制加工。由于外在生成的感觉(刺激物)与内在生成的感觉(伴随体验)之间的不一致是联觉的一个内在部分,这就预示着在联觉的大脑区域网络中应该有与认知控制相关的某个区域,虽然右背外侧前额叶皮层是一个重要的备选区域,然而Weiss等人(2005)却发现在物理引起的字母颜色与联觉感知的颜色之间的不一致增加了在右梭形回和左背外侧前额叶皮层的激活,而另一个研究则提出一个任务来显示了一致性效应控制了在左顶叶的内沟和角回的激活。到目前为止,还无法解释为什么会出现这样的差异性。
从以上对联觉颜色的全脑激活情况的对比分析,我们可以越来越清晰地看到联觉体验不仅激活了一个特定区域(包括V4的视觉区域),而且激活了一个大脑区域网络(五个大脑区域),这个大脑区域网络与联觉的三个不同认知加工过程相关:感觉加工、(注意)捆绑加工和认知控制加工。虽然在这个大脑区域网络中,视觉区域和顶叶区域一直是研究者持续、重点关注的区域,但我们还无法知晓这两个区域在联觉中是否就起到关键性的作用。另外,习得的联觉研究已经表明了在联觉与丘脑之间有一种关联:一个丘脑病变导致习得的联觉(Beauchamp&Ro,2008)。但在前面没有提到联觉与丘脑之间的关联,这是因为丘脑的激活在神经影像中很少被报道,在联觉研究中一直没有仔细地分析丘脑。还有一个没有获得证据支持的是颞上皮层,这个多感觉大脑区域曾经被推测是通过回馈连接到单级感觉皮层来影响联觉的区域,但目前的研究中还没有证据能够支持在这个大脑区域与联觉体验之间具有关联。
为数不多的几个研究测量了其他类型联觉的大脑激活,从这几个研究我们也能得到类似的发现。第一是在对应于特定联觉体验的大脑感觉区域有增加的激活。一个习得的声音—触觉联觉者显示了在联觉期间第二体感皮层有增加的激活(Beauchamp&Ro,2008);而对气味—疼痛联觉者(某种气味增加了神经性疼痛的病人)的研究显示了在由气味引起的疼痛期间与疼痛有关的区域有增加的激活,以及在手臂区域的第一体感皮层有增加激活,即人感到神经性疼痛的区域。第二是像颜色联觉一样,也激活了一个大脑区域网络,而非激活了独立的特定感觉区域。在这些研究中大脑的几个区域被测量,而非仅仅特定的感觉激活的区域,并且都发现了联觉体验激活了这些区域(Jäncke&Langer,2011;Specht&Laeng,2011)。第三是联觉体验激活了关联性很强的中心区域,中心区域是一个与其他区域具有很强互联性的区域,而联觉体验激活一个中心区域就意味着其激活了一个大脑区域网络。Jäncke和Langer(2011)发现左侧顶叶皮层除了是相关的感觉大脑区域(听觉皮层)之外,还是一个很强的中心区域,他们不仅仅探究大脑的一个单独区域,而且还探究了大脑的几个连接区域。他们使用图理论分析显示在不同类型的功能中涉及到不同的联觉中心区域,这说明联觉的不同功能涉及不同的大脑区域网络。那么这些大脑区域网络是如何与联觉的不同功能相联系的呢?这是目前正在进行的工作。Jones等人(2011)发现情绪效价诱发嗅觉时,有脑岛激活,并且发现联觉的强度与内侧顶叶激活相关;Specht和Laeng(2011)发现了一个感觉网络,涉及后颞叶、枕下叶以及右额叶大脑区域;Amin等人(2011)发现一个人格化字母的联觉者有楔前叶激活,并且将这联系到联觉的一般方面:心理意象和自我参照加工能力。当前还不清楚这种结果的最好解释是什么,但这些令人感兴趣的发现支持了在联觉中的不同功能与大脑区域的不同网络相关联,将来的研究需要进一步证实联觉的这些功能以及涉及的大脑区域网络,并提供模型的解释。
上面讨论的所有发现都是通过使用fMRI或PET来测量联觉者在联觉体验期间的大脑激活而获得的,一个很自然的问题就是:为什么联觉者的这些大脑区域能被激活,而非联觉者却不能?一个具有竞争性的回答就是联觉者与非联觉者的大脑存在结构上的差异。
二 联觉:大脑结构的差异
自2007年以来,3个研究已经使用各向异性分数(FA)来测量白质束的一致性,并探讨在联觉者与非联觉者之间大脑结构上的差异(Hänggi et al.,2008;Jäncke et al.,2009;Rouw & Scholte,2007),而5个研究则根据灰质来探讨了这些差异(Hänggi et al.,2008;Jäncke et al.,2009;Weiss& Fink,2009;Rouw&Scholte,2010;Banissy et al.,2012)。虽然目前对联觉者与非联觉者之间大脑结构差异的研究有限,但从这些研究我们仍能得到一些有价值的发现。
首先,这些数据支持在刺激物与伴随体验这两个区域的交叉激活理论。联觉的交叉激活理论假设了刺激物和伴随体验的神经表征应当是位于密集的相互关联的大脑区域,这些区域通常会是相互毗邻连接的,但不必是同一个区域。Rouw和Scholte(2007)发现了在V4区临近的梭形回附近的增加连接,这种发现也被Jäncke等人(2009)部分地证实(在右梭形回,阈值p=.05.);Weiss和Fink(2009)则更进一步地显示了在V4区有增加的灰质;而Hänggi等人(2008)在一个声调或一个声调音程引发的联觉情况中也发现了增加的连接,以及在第一听觉皮层发现了增加的灰质和白质体积。
其次,联觉应与感觉区域的灰质密度增加有关。Hänggi等人(2008)和Jäncke等人(2009)发现了在V1和V2区增加的灰质,虽然Hänggi等人的研究测验的是音程—味觉和声调—颜色联觉,而Jäncke等人(2009)测验的是字形—颜色联觉;同时,Jäncke等人(2009)在测验字形—颜色联觉时还发现在第二体感皮层中有增加的灰质。Rouw和Scholte(2007,2010)也发现了在V1、听觉和体感皮层有增加的灰质,特别是在投影仪中与关联的联觉者比较时。Banissy等人(2012)则通过对字形和音程颜色联觉者的检测研究发现在左后梭形回中有增加的灰质,但在左梭形回和左MT/V5的前部区域有伴随减少。
最后,依据白质和灰质能够发现联觉者和非联觉者在不同的大脑区域有结构差异,这显示了联觉是与整个大脑的大范围结构差异相关,而非仅仅涉及交叉体验的加工区域。这些广泛的差异表明了不同的大脑特征可能与特定联觉类型相关,因此在解释联觉功能的差异时应当考虑这种结构上的差异,群体之间结构上的差异能够导致这些群体在fMRI激活中的差异。比如,如果联觉者的V4区普遍比控制组的更大,因为不同的基准,fMRI将会显示出联觉者比非联觉者的激活更多。
这些研究的结果意味着联觉者的大脑与非联觉者的大脑有结构上的差异——对应于刺激物和伴随信息的感觉大脑皮层存在结构上的差异,并且这种结构上的差异不仅仅局限在感觉大脑区域,在不对应特定联觉体验的感觉区域也存在着这种差异,这意味着大脑结构的差异不仅仅限于与联觉类型相关,这些大脑结构上的差异更可能是与遗传基因相关。
三 联觉的遗传基因
现代研究已经确认了联觉往往具有家族遗传性。Barnett等人(2008)征集了53个联觉者,42%的联觉者报道具有联觉的直系亲属,他们直接与这些直系亲属相联系,收集到17个家族所有成员的有关联觉状态的完全数据。他们通过研究发现同一个家族中会出现不同类型的联觉者,但家庭成员之间的体验有所不同,联觉关联和联觉者个体之间的差异会受其他因素的影响。
当前,两个研究对联觉的遗传基因机制进行了深入探讨。Asher等人(2009)为了搜寻与联觉有关的多态性,做了建立在43个多重家庭的全基因组连锁研究。在这个研究中,选择的联觉者是听觉(视觉联觉者,由于它没有进一步指定对于这些联觉者哪种特定的材料(声音、口语、书面语、单音素和字形)引起了颜色,这样就不可能构成一个完全同质的对象组。这个研究结果表明在这个对象组中联觉与多重的基因位点相关 (2q24,5q33,6p12,12p12),并且没有显示出联觉是与X-染色体有关系的。在这个研究中的最高负荷标志包含TBR1基因,这个基因在大脑皮层的发展中起重要作用,并且参与大脑调节。针对这个基因的基因敲除技术研究已经显示了大脑的解除调节导致在大脑薄板状组织中的失常,并且影响轴突的路径搜索。Tomson等人(2011)研究了颜色联觉的基因机制,他们对具有颜色序列联觉的5个家族中的48个个体的DNA进行了关联分析,发现颜色联觉与在16q12.2-23.1区域的一个基因相关,虽然他们在这次研究中还没法具体确定是哪一个基因,但是给出了6个可能的基因:GABARAPL2、NDRG4、PLLP、KATNB1、CIAPIN1、GNAO1。同样,在他们的研究中也没有基因证据支持联觉是与X-染色体有关系,这与Asher等人(2009)的结果一致。
有研究发现联觉者女性多于男性,从而把女性具有联觉优势的现象归于可能是X-染色体上的基因有关(Ward&Simner,2003)。然而从目前的基因研究来看,还没有证据发现这是与X-染色体基因相关,这就带来一个令人疑惑的问题:如果女性具有联觉优势的现象真的与X-染色体基因无关,那么这种现象又该如何解释呢?目前还没有发现对于女性具有联觉优势的现象有其他解释。同时,这些研究发现的那些决定联觉的可能基因都是在大脑中影响连接发展的基因,因此,我们相信联觉的基因基础可能将会在大脑中影响连接发展的那些基因中(至少部分)找到,但具体是哪一个基因,还需以后的基因研究进一步证实。
基因遗传在形成联觉中扮演重要角色,那么是否基因遗传就是形成联觉的仅有因素呢?大多数联觉(约88%)是被习得的语言单位所激发,或者涉及在童年中后期习得的认知建构,而联觉的基因遗传往往无法解释这些带有习得性的特征(Simner,2012)。一种潜在的可能性就是联觉在后期能够发展变化,联觉的出现除了基因遗传基础之外,还存在在合适的环境中发展与习得的可能。既然联觉是发展的,那么联觉是如何发展的呢?西姆纳等人(Simner et al,2013)对儿童时期的字体-颜色联觉发展进行了行为实验的研究,他们发现一些具有联觉的儿童随着年龄的增加联觉体验会消失,而另一些具有联觉的儿童的关联的范围则会增宽,新的联觉关联会出现。他们的研究仅仅针对的是儿童,对于少年、青年、中年以及老年联觉情况是否也具有这样的特征,联觉的一个整体发展状况是什么样的,目前还没有联觉研究对此提供出全面的证据。儿童时期能够发展出新的联觉关联,那么一个与此相关的问题就是:非联觉者能否通过训练习得联觉关联?这是目前一个具有争议性的问题,因为一些研究发现非联觉者经过配对关联训练后具有类似于真正联觉者的行为,而同时发现这些经过训练后的非联觉者与真正的联觉者又有一些差异(Meier&Rothen,2009;Rothen et al.,2011),那么非联觉者经过配对关联训练后能否成为真正的联觉者,还有待于以后进一步的实验证实。
四 小结与展望
通过对近几年联觉研究成果的总结与分析,我们可以得出如下结论:
(1)联觉体验不仅激活了特定区域(V4区域或顶叶区域),而且涉及一个大脑区域网络(6个区域),这个大脑区域网络是与联觉的三个不同加工过程有关:感觉加工、(注意)捆绑加工和认知控制加工。
(2)联觉具有基因遗传性,目前的基因研究已经确定了联觉的几个备选基因,比如TBR1,这几个备选基因都属于大脑中影响连接发展的那些基因。虽然目前还无法确定确切是哪个(哪些)基因,但我们相信决定联觉的基因可能在大脑中影响连接发展的那些基因中(至少部分)找到。
(3)不少类型的联觉诱发物和伴随物都涉及高层次的语言范畴认知和概念建构,这些高层次的语言范畴和概念是联觉者在具体的环境中习得的结果,因此基因遗传性不是决定联觉的唯一因素,联觉还具有发展的可能。
虽然联觉的脑机制研究已经取得初步的进展,但还仅仅处于一个“幼年期”,我们对联觉的脑机制所知只是“冰山一角”,还有不少问题需进一步揭示。首先,这些大脑区域网络(不同的大脑区域)如何与联觉的不同功能相关联?近期的发现(Jones et al.,2011;Amin et al.,2011)已经显示了联觉的不同功能是与不同的大脑区域有关的,将来的研究还需要进一步证实这些特定的区域。其次,联觉具有基因遗传性已经得到证实,但确切是哪个基因(或哪些基因)决定目前还无法确定,以后的基因研究还需进一步证实。最后,联觉是如何发展的以及联觉能否通过训练习得,还有待于将来的研究证实。
黄希庭.(2007).心理学导论.北京:人民教育出版社.293.
Amin,M.,Olu-Lafe,O.,Claessen,L.E.,Sagiv,N.(2011).Understanding grapheme personification:A social synaesthesia?Journal of Neuropsychology,5,255–282.
Asher,J.E.,Lamb,J.A.,Brocklebank,D.Monaco,A.P.(2009).A whole-genome scan and fine-mapping linkage study of auditoryvisual synesthesia reveals evidence of linkage to chromosomes 2q24,5q33,6p12,and 12p12.The American Journal of Human Genetics,84(2),279–285.
Banissy,M.J.,Stewart,L. ,Kanai,R.(2012).Grapheme-color and tone-color synesthesia isassociated with structural brain changes in visual regions implicated in color,form,and motion.Cognitive Neuroscience,3(1),29–35.
Barnett,K.J.,Finucane,C. ,Mitchel,K.J.(2008).Familial patterns and the origins of individual differences in synaesthesia.Cognition,106(2),871–893.
Beauchamp,M.S.,& Ro,T.(2008).Neural substrates of sound-touch synesthesia after a thalamic lesion.Journal of Neuroscience,28,13696–13702.
Eagleman,D.M.(2012).Synaesthesia in its protean guises.British Journal of Psychology,103,16–19.
Hänggi,J.,Beeli,G.,Oechslin,M.S.,& Jäncke,L.(2008).The multiple synaesthete ES – Neuroanatomical basis of interval-taste and tone-colour synaesthesia.Neuroimage,43(2),192–203.
Hubbard,E.M.,Arman,A.C.,Ramachandran,V.S.,& Boynton,G.M.(2005).Individual differences among grapheme-color synesthetes:Brain-behavior correlations.Neuron,45(6),975–985.
Jäncke,L.,Beeli,G.,Eulig,C.,& Hänggi,J.(2009).The neuroanatomy of grapheme-color synesthesia.European Journal of Neuroscience,29(6),1287–1293.
Jäncke,L.,& Langer,N.(2011).A strong parietal hub in the small-world network of coloured hearing synaesthetes during resting state EEG.Journal of Neuropsychology,5,178–202.
Jones,C.L.,Gray,M.A.,Minati,L.,Simner,J.,Critchley,H.D.,& Ward,J.(2011).The neural basis of illusory gustatory sensations:Two rare cases of lexical– gustatory synaesthesia.Journal of Neuropsychology,5,243–254.
Laeng,B.,Hugdahl,K.,& Specht,K.(2011).The neural correlate of colour distances revealed with competing synaesthetic and real colours.Cortex,47(3),320–331
Meier,B.,& Rothen,N.(2009).Training grapheme-colour associations produces a synaesthetic Stroop effect,but not a conditioned synaesthetic response.Neuropsychologia,47(4),1208–1211.
Novich,S.,Cheng,S.,& Eagleman,D.M.(2011).Is synaesthesia one condition or many?A large scale analysis reveals subgroups.Journal of Neuropsychology,5,353–371.
Nunn,J.A.,Gregory,L.J,Gray,J.A.(2002).Functional magnetic resonance imaging of synesthesia:Activation of V4/V8 by spoken words.Nature Neuroscience,5(4),371–375.
Paulesu,E.,Harrison,J.,Frith,C.D.(1995).The physiology of coloured hearing a PET activation study of colour-word synaesthesia.Brain,118(3),661–676.
Rothen,N.,Wantz,A.L.,Meier,B.(2011)Training synaesthesia.Perception,40,1248—1250.
Rouw,R.,Scholte,H.S.,& Colizoli,O.(2011)Brain areas involved in synaesthesia:A review.Journal of Neuropsychology,5,214 –242.
Rouw,R.,& Scholte,H.S.(2007).Increased structural connectivity in grapheme-color synesthesia.Nature Neuroscience,10(6),792–797.
Rouw,R.,& Scholte,H.S.(2010).Neural basis of individual differences in synesthetic experiences.Journal of Neuroscience,30(18),6205–6213.
Simner J.(2012).Defining synaesthesia.British Journal of Psychology,103,1–15
Simner,J.,& Bain,A.E.(2013).A longitudinal study of grapheme-color synaesthesia in childhood:6/7 years to 10/11 years.Frontiers in human neuroscience,7:603.
Specht,K.,& Laeng,B.(2011).An independent component analysis of fMRI data of grapheme– colour synaesthesia.Journal of Neuropsychology,5,203–213.
Sperling,J.M.,Prvulovic,D.,Linden,D.E.J.,Singer,W.,& Stirn,A.(2006).Neuronal correlates of colour-graphemic synaesthesia:A fMRI study.Cortex,42(2),295–303.
Steven,M.S.,Hansen,P.C.,& Blakemore,C.(2006).Activation of color-selective areas of the visual cortex in a blind synesthete.Cortex,42(2),304–308.
Tomson,S.N.,Avidan,N.,Eagleman,D.M.,(2011).The genetics of colored sequence synesthesia:Suggestive evidence of linkage to 16q and genetic heterogeneity for the condition.Behavioural Brain Research,223,48–52.
Ward,J.,& Simner,J.(2003).Lexical–gustatory synaesthesia:Linguistic and conceptual factors.Cognition,89,237–261.
Weiss,P.H.,Shah,J.N.,Toni,I.,Zilles,K.,& Fink,G.R.(2001).Associating colours with people:A case of chromatic-lexical synaesthesia.Cortex,37,750–753.
Weiss,P.H.,Zilles,K.,& Fink,G.R.(2005).When visual perception causes feeling:Enhanced cross-modal processing in graphemecolor synesthesia.NeuroImage,28(4),859–868.
Weiss,P.H.,& Fink,G.R.(2009).Grapheme-colour synaesthetes show increased grey matter volumes of parietal and fusiform cortex.Brain,132(1),65–70.
猜你喜欢
中国医学影像技术(2020年11期)2021-01-04
中国现代医药杂志(2020年3期)2020-05-08
现代青年·精英版(2019年9期)2019-09-25
中国生物医学工程学报(2019年6期)2019-07-16
中国医学影像技术(2019年3期)2019-03-25
三联生活周刊(2018年5期)2018-01-27
中国CT和MRI杂志(2016年11期)2017-01-18
中外医疗(2016年15期)2016-12-01
安徽农业科学(2015年10期)2015-04-24
心理科学进展(2015年1期)2015-01-23
