
水温和体重对白斑红点鲑临界游泳速度和游动耗氧率的影响
2015-02-28 08:28张永泉尹家胜徐革峰席庆凯马波白庆利
水生生物学报 2015年4期
张永泉尹家胜徐革峰席庆凯马 波白庆利
(1. 中国水产科学研究院黑龙江水产研究所, 哈尔滨 150070; 2. 东北农业大学动物科技学院, 哈尔滨 150030)
水温和体重对白斑红点鲑临界游泳速度和游动耗氧率的影响
张永泉1尹家胜1徐革峰2席庆凯1马 波1白庆利1
(1. 中国水产科学研究院黑龙江水产研究所, 哈尔滨 150070; 2. 东北农业大学动物科技学院, 哈尔滨 150030)
为了揭示水温和体重对白斑红点鲑(Salvelinus leucomaenis)临界游泳速度和游动耗氧率的影响, 利用试验生态学方法测定了不同体重(1龄组、2龄组和3龄组)的白斑红点鲑在4、8、12、16、20和24℃共6个水温的临界游泳速度和游泳耗氧率。结果表明: 水温和体重对临界游泳速度的单独效应均显著(P<0.05),但水温和体重交互作用效应却不显著(P>0.05), 在相同水温下白斑红点鲑临界游泳速度均随着年龄的增长(体重增加)而增加。水温4℃时3个年龄组白斑红点鲑临界游泳速度均最低, 分别为(21.6±1.06)、(22.93±0.61) 和(30.27±1.29) cm/s, 随着水温的升高临界游泳速度均不断增加, 当水温升高到16℃时临界游泳速度达到最大值, 分别为(39.6±0.80)、(46.80±0.80)和(53.73±1.22) cm/s, 此后随着水温进一步升高到 20℃, 临界游泳速度虽出现略微降低, 但经统计分析 16℃和 20℃时临界游泳速度无显著性差异(P>0.05), 当水温达到 24℃时却出现明显降低。水温和流速以及二者的交互作用对游泳耗氧率的影响均达到显著水平(P<0.05), 白斑红点鲑在适宜的相同水温和流速时体重越大其游泳耗氧率越低, 整体观察 3个年龄组白斑红点鲑鲑游泳耗氧率均随着水温和流速的增高而增高, 但当水温和流速升高到一定值游泳耗氧率却出现降低。研究得出体重较大的个体在相同水温下抵抗水流的游泳能力较强, 3个年龄组白斑红点鲑适宜的最高水温不应超过20℃, 最高流速依次不应超过32、40和48 cm/s。
白斑红点鲑; 水温; 流速; 耗氧率; 临界游泳速度
白斑红点鲑(Salvelinus leucomaenis)隶属鲑科红点鲑属, 主要分布在太平洋西北部, 即白令海, 鄂霍次克海和日本海, 向南到海参崴, 仅有小部进入黑龙江河口, 我国红点鲑属鱼类仅有 2种, 即花羔红点鲑(Salvelinus malm)和白斑红点鲑[1]。白斑红点鲑鱼体呈纺锤形, 体被圆鳞且细小, 身体背部呈褐色, 体侧布满白色或灰色小斑点, 故名白斑红点鲑。由于近年来的环境污染和过渡捕捞, 该鱼的资源已基本枯竭, 多年未见有捕捞到野生鱼的报道。中国水产科学研究院黑龙江水产研究所于1996年12月从日本东京水产大学引进发眼卵1.0万粒, 此后, 国内陆续出现了白斑红点鲑的相关报道。本研究团队经过多年研究, 目前已成功实现了白斑红点鲑的全人工繁殖和苗种培育[2, 3], 并已在全国 10多省市自治区推广养殖。
国外早在1933年就对鱼类的耗氧率和运动能力方面进行了研究[4], 我国则是在20世纪80年代随着渔业的蓬勃发展才陆续开展相关研究[5, 6]。呼吸和游动是鱼类最基本的生命活动, 因此也是生物能量学研究的重要内容之一, 其不仅能反映鱼类的生理健康状态, 也可反映外部环境条件对鱼类生理活动的影响程度[6, 7]。鱼类呼吸和游动能力主要受体重、摄食水平等生物因素和水温、盐度、水流等非生物因素的影响[5]。鱼类作为生活在水体中的变温动物, 与哺乳类和禽类等其他脊椎动物的呼吸代谢和运动行为存在很大差异, 鱼类呼吸代谢和运动行为对水温和水流的变化极为敏感。近年国内外学者对生物和非生物因素影响下的欧洲鲈(Dicentrarchus labrax)[8]、圆口铜鱼(Coreius guichcnoti)[9]、罗非鱼(Oreochromis niloticus)[10]、巨须裂腹鱼(Schizothorax macropogon)[11]、南方鲇(Silurus meridionalis)[12]和细鳞鲑(Brachymystax lenok)[13]等鱼类代谢和游动行为进行了大量的研究, 但有关我国白斑红点鲑的相关研究尚未见报道,本文利用进口的鱼类生态学装置, 测定了3个年龄组的白斑红点鲑在不同水温和流速作用下的临界游泳速度和游泳耗氧率变化, 旨在了解水温、流速和体重对白斑红点鲑呼吸和游泳能力的影响规律, 以期为白斑红点鲑自然资源的复壮和人工养殖提供理论依据和数据参考。
1 材料与方法
1.1 试验动物
试验动物来自中国水产科学研究院黑龙江水产研究所渤海冷水鱼试验站, 挑选健康规格相近, 鱼体完整且游动正常的1龄(90尾)、2龄(60尾)和3龄(54尾)的白斑红点鲑, 1龄组体重为(46.67±3.24) g, 体长为(12.5±1.42) cm, 2龄组体重为(185.4±5.83) g, 体长为(25.8±2.86) cm, 3龄组体重为(320±10.43) g, 体长为(31.73±4.39) cm。
1.2 试验方法
试验前首先进行预试验确定水温范围, 分别选取1龄、2龄和3龄的白斑红点鲑放入控温循环水族箱,每日10: 00和15: 00投喂2次, 每次投喂前清理粪便和残饵, 每日换水1/3。起始水温为4℃, 每个水温下暂养20d后, 依次升高4℃, 每个水温下注意观察鱼类的摄食和游动。根据预试验得出: 水温4℃白斑红点鲑很少进食, 且生长十分缓慢, 但并未出现死亡,随着水温升高, 鱼类摄食和生长情况出现好转, 但水温达24℃时, 白斑红点鲑进食活动减少, 鱼体明显消瘦, 持续7d以上开始出现死亡, 且死亡量逐渐增加; 当水温升高到28℃时, 24h内全部死亡。因此本研究将试验水温范围设定为4—24℃, 共设置4、8、12、16、20和24℃六个水温梯度, 每个水平的测定设3个重复组。
该试验于2012年7月采用丹麦进口鱼类生态测量仪完成(图1), 该仪器可任意调节水温和流速。在进行不同水温试验前, 将控温水族箱的水温提前调节至试验水温, 将鱼放入控温水族箱中暂养24h, 随机取出白斑红点鲑放入生态测量仪中, 让试验鱼适应2h以降低环境更替带来的压力, 期间水流速度控制在8 cm/s, 在每个试验水温条件下, 采用逐步提速法进行, 起始测定流速设为8 cm/s, 每次流速增量为8 cm/s。

图1 鱼类生态测定仪器Fig. 1 The measuring instruments of fish ecology
1.3 试验参数和数据的测量
为了研究水温和体重对白斑红点鲑临界游泳速度的影响, 分别在每个水温条件下进行3个年龄组白斑红点鲑最大有氧持续游泳速度, 即临界游泳速度(Critical swimming speed, Uc)的测定, 在测定上述Uc的同时测定其实时游泳耗氧率(2OM ), 游泳耗氧率的测定截止到鱼类力竭流速的耗氧率, 即截止到“V+ V”流速下的耗氧率, 耗氧率为丹麦进口鱼类生态测量仪全自动测量。参考徐革锋等[13]和张伟等[14]研究方法, 对白斑红点鲑Uc采取以下公式进行计算: Uc= V + (t/T) V。式中, Uc为临界游泳速度(cm/s); T为在各速度梯度下设定的持续游泳时间间隔20min; V为完成设定时间间隔20min游泳所具有的最大速度(cm/s), 即鱼力竭前一档速度, 力竭的判断依据为试验鱼拒绝游泳, 并停留在生态测量仪工作区下游遮挡网处大于20s; V 是各速度梯度的速度增量, t为试验鱼在“V+ V”速度下的持续时间。
1.4 数据的处理
数据采用 Excel和 SPSS 13.0软件进行统计分析,首先利用[Analyz]→[General Linear Mode]→[Univariate]模块进行双因素方差分析, 确定单因素和双因素的交互效应是否显著。单因素的效应显著, 对组间作多重比较, 所有试验结果均以平均值±标准差(Mean± SD)表示, P<0.05 表示差异显著, P>0.05 表示差异不显著。
2 结果
2.1 水温和体重对白斑红点鲑临界游泳速度的影响
对水温和体重与临界游泳速度进行双因素方差分析的效应性检验, R2=0.871, 水温和体重与临界游泳速度的单独效应均显著(P<0.05), 但水温和体重交互作用效应却不显著(P>0.05)(表1)。由图2可见, 不同年龄组(体重不同)白斑红点鲑的临界游泳速度随水温变化明显, 整体观察相同水温下临界游泳速度均随着年龄的增长而增加。1龄组白斑红点鲑临界游泳速度最低出现在水温4℃情况下, 为(21.6±1.06) cm/s,水温8℃时临界游泳速度[(22.67±0.67) cm/s]虽略高于4℃, 但二者间却无显著性差异(P>0.05), 随着水温升高临界游泳速度不断增加, 当水温升高到16℃时, 临界游泳速度达到最大值[(39.6±0.80) cm/s], 此后随着水温进一步升高, 临界游泳速度却出现降低,水温20℃时降低较少[(37.87±0.24) cm/s], 经统计分析16℃与20℃时临界游泳速度无显著性差异(P>0.05), 但水温24℃却呈现显著性降低。2龄组和3龄组白斑红点鲑临界游泳速度随水温的变化特性基本相同, 最低值均出现在水温4℃的情况, 分别为(22.93±0.61)和(30.27±1.29) cm/s, 随水温的升高临界游泳速度均出现显著性增长(P<0.05), 当水温达到16℃临界游泳速度同样达到最大值, 分别为(46.80±0.80)和(53.73±1.22) cm/s, 当水温升高到20℃2龄组与3龄组的临界游泳速度略有降低, 但同一年龄组16℃和20℃水温下的临界游泳速度, 却无显著性差异(P>0.05), 此后随着水温的进一步升高到24℃,两个年龄组的临界游泳速度均出现了显著性降低(P<0.05)。
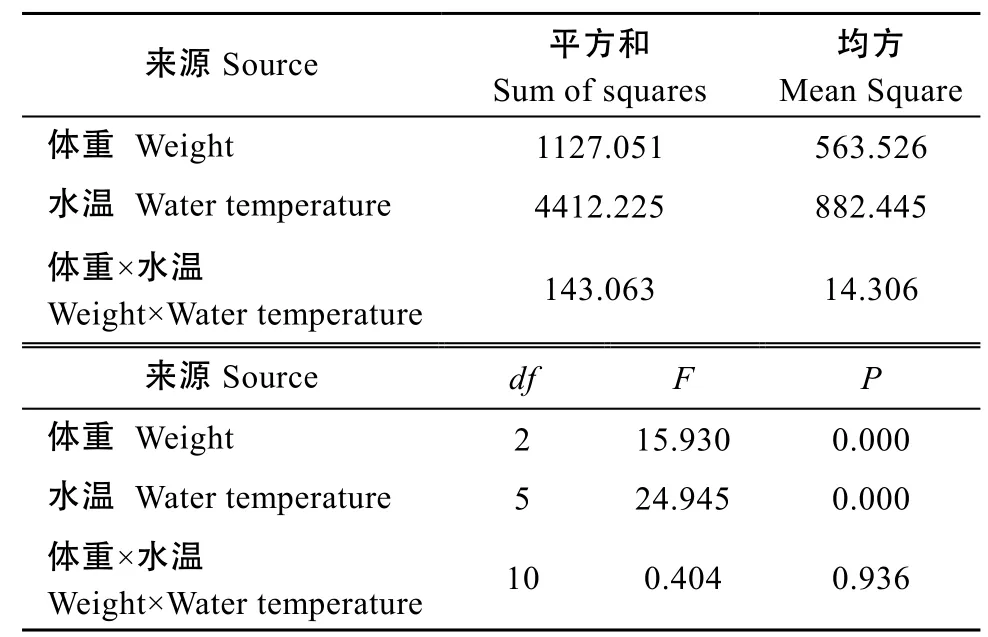
表1 水温和体重与临界游泳速度的双因素方差分析Tab. 1 Two-factor variance analysis of the water temperature, the body weight, and the critical swimming speed
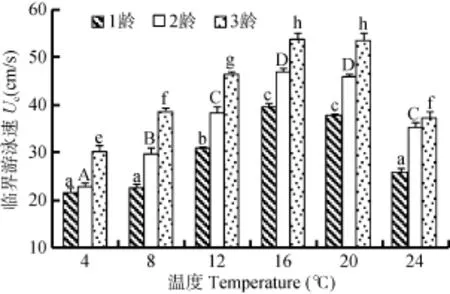
图2 水温对白斑红点鲑临界游泳速度的影响Fig. 2 Effects of the water temperature on the critical swimming speed of Salvelinus leucomaenis
2.2 水温和流速对不同体重白斑红点鲑游泳耗氧率的影响
在测定临界游泳速度的同时测定其实时游泳耗氧率(2OM ), 分别对3个年龄组的水温和流速与游泳耗氧率进行双因素方差分析的检验表明, 3个年龄组统计分析的R2分别为0.900、0.964和0.961, 水温、流速、水温和流速交互作用效应均显著(P<0.05)(表2)。3个年龄组白斑红点鲑游泳耗氧率最大值均随着水温升高而增高, 白斑红点鲑游泳耗氧率随着水温和流速的增加而增高, 但当水温和流速升高到一定值游泳耗氧率却出现降低的现象, 且白斑红点鲑在适宜的相同条件下体重越大其耗氧率却越低(图3)。在水温4℃水体中白斑红点鲑1龄组和2龄组不同流速下的游泳耗氧率比较接近, 但明显高于3龄组, 1龄组、2龄组和3龄组均是流速8 cm/s时游泳耗氧率最低, 分别为(28.37±3.50)、(28.61±1.91)和(15.13±2.26) mg/(g·h), 此后随着水流速的增加均出现显著性增长(P<0.05);在水温8℃水体中1龄、2龄和3龄组游泳耗氧率的最低值均出现在流速为8 cm/s时, 此后虽随着水流速的增加均出现了增长, 但增长幅度却出现明显差异。1龄组当流速由8 cm/s增至16 cm/s时游泳耗氧率增加幅度最大, 16 cm/s升至24 cm/s时增幅却明显降低, 但16 cm/s和24 cm/s时的游泳耗氧率存在显著性差异(P<0.05)。2龄组的游泳耗氧率当流速8 cm/s增至16 cm/s时出现较大幅度的增长, 此后虽然随着流速的增加出现增长, 但幅度明显变缓。3龄组当流速由8 cm/s增至24 cm/s时游泳耗氧率增幅度相对较大,当流速24 cm/s增至40 cm/s时增幅度相对较低, 经统计2龄组24 cm/s与32 cm/s和3龄组32 cm/s与40 cm/s都存在显著性差异(P<0.05); 在水温12℃水体中1龄组流速由8 cm/s增至24 cm/s时随流速的增加游泳耗氧率增幅较大, 虽24 cm/s升至32 cm/s时游泳耗氧率增幅出现明显降低, 经统计24 cm/s和32 cm/s时的游泳耗氧率差异显著(P<0.05)。2龄组和3龄组的变化规律十分相似, 两个龄组均是当流速由8 cm/s增至16 cm/s时游泳耗氧率增加幅度相对较小, 此后增幅开始变大, 2龄组当水流速由32 cm/s升至40 cm/s时增幅出现明显降低, 3龄组水流速40 cm/s升至48 cm/s时同样出现游泳耗氧率增加幅度降低的现象, 经统计2龄组32 cm/s与40 cm/s和3龄组40 cm/s 与48 cm/s时的游泳耗氧率均无显著性差异(P>0.05);在水温16℃水体中1龄组流速8 cm/s至32 cm/s时游泳耗氧率增加较快, 但当流速32—40 cm/s时游泳耗氧率却无显著性增加(P>0.05), 仅增加3.42 mg/(g·h)。2龄组在流速32—40 cm/s时游泳耗氧率出现了快速增长, 此后40—48 cm/s时增长不显著(P>0.05)。3龄组流速8—40 cm/s时增加幅度相对较小, 当40—48 cm/s时出现快速增加期, 此后增长幅度明显降低;在水温20℃水体中1龄组在流速8—32 cm/s时游泳耗氧率急剧增长, 当达到32 cm/s时达到最大值(350.45± 14.39) mg/(g·h), 此后随着水流速的增高游泳耗氧率出现显著性降低(P<0.05)。2龄组和3龄组在流速8 cm/s时游泳耗氧率十分接近, 此后随着流速的增加游泳耗氧率迅速增加, 2龄组达到40 cm/s和3龄组到48 cm/s时游泳耗氧率达到最大值, 此后游泳耗氧率显著性降低(P<0.05); 在水温24℃水体中1龄组当水流速达到24 cm/s时游泳耗氧率达到最大值(423.59 ±24.04) mg/(g·h), 当流速升高到32 cm/s时游泳耗氧率降低到最低值(349.10±19.42) mg/(g·h)。2龄组和3龄组随着流速增加游泳耗氧率不断增加, 当达到32 cm/s时游泳耗氧率分别达到最大值(481.35±15.40)和(453.56±24.57) mg/(g·h), 此后随着流速增加游泳耗氧率明显降低。
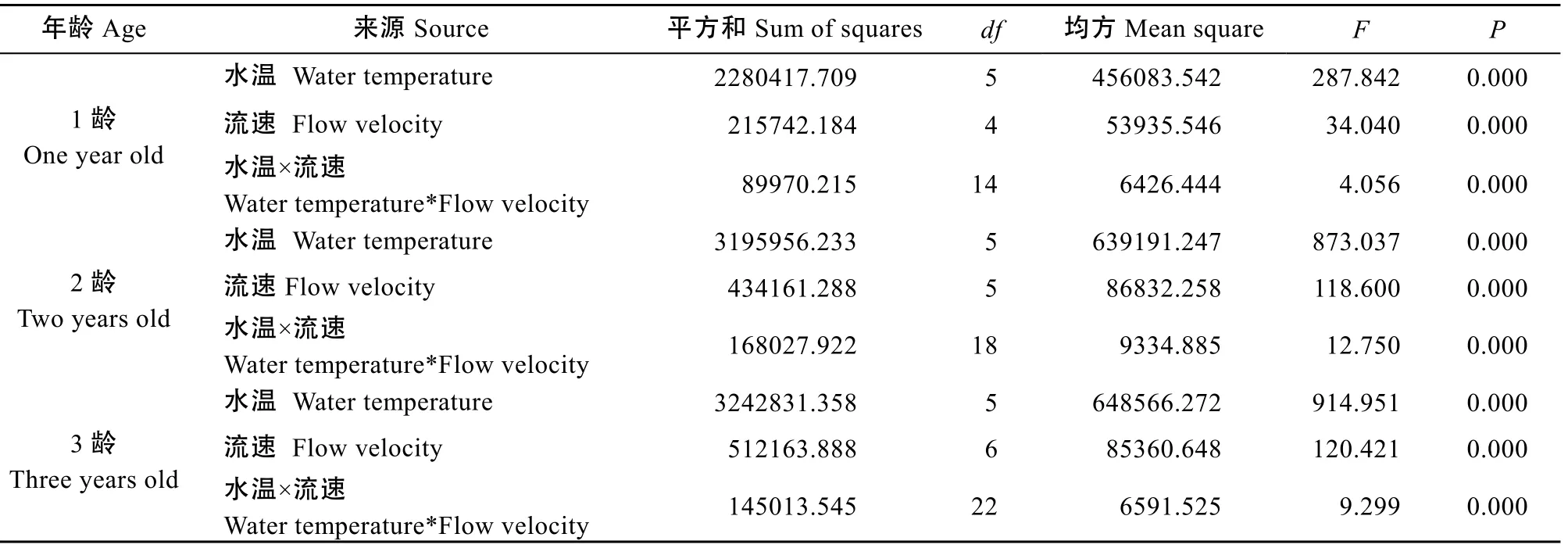
表2 不同年龄组水温和流速与游泳耗氧率的双因素方差分析Tab. 2 Two-factor variance analysis of the water temperature, the flow velocity and the oxygen consumption rate in different age groups

图3 在不同水温下流速对3个年龄白斑红点鲑游动耗氧量的影响Fig. 3 Effects of the water velocity on the oxygen consumption of Salvelinus leucomaenis at three different ages at different water temperatures
3 讨论
3.1 水温、流速和体重与白斑红点鲑游泳能力的关系
游泳运动为鱼类捕食、逃逸、洄游和繁殖等提供必要的保障, 是其重要的生存活动方式[15]。水温和水流作为鱼类生活环境的非生物性因子, 能够刺激鱼类的感觉器官, 使其产生相应的活动方式及反应机制[11]。1958年Brett等[16]首次提出临界游泳速度,即鱼类最大有氧持续游泳速度, 多年来国内学者针对鱼种类、体重、摄食水平等生物因素和水温、盐度、水流等非生物因素的影响进行了深入的研究,并将临界游泳速度作为了衡量鱼类变速游泳能力的重要评价指标之一[17—19]。本文研究了水温和体重与临界游泳速度的关系, 研究发现同等水温下白斑红点鲑临界游泳速度均随着年龄的增长而增加, 即体重较大的个体在相同水温下抵抗水流的游动能力较强。3个年龄组白斑红点鲑临界游泳速度的最低值均出现在低温区, 随着水温的升高临界游泳速度均在增加, 当水温升高到16℃时临界游泳速度达到最大值, 此后随着水温进一步升高到20℃, 临界游泳速度虽出现略微降低, 但经统计分析16℃和20℃时临界游泳速度无显著性差异(P>0.05), 当水温达到24℃时却出现明显降低。Boutilier等[20]和徐革锋等[13]通过对七鳃鳗(Petromyzon marinus)和细鳞鲑的研究认为能量底物(Energy stores)被快速消耗殆尽以及乳酸等代谢产物的累积过度是导致临界游泳速度随水温变化的主要因素。本研究分析得出白斑红点鲑临界游泳速度最低值出现在低温区, 应该是低温限制能量底物的动员、活化和利用, 使得能量转化率降低, 鱼体能量底物浓度供给不足, 进而导致临界游泳速度较低。随着水温的升高能量转化率不断提高,能量底物浓度逐渐增加, 鱼类抵抗水流的游动能力增强, 其临界游泳速度也在增加。在高温24℃时随着流速不断增加, 力竭性运动导致有氧呼吸提供能量已不能满足机体需要, 无氧代谢过早启动, 致使乳酸等代谢产物迅速积累过多抑制了有氧代谢效率,因此出现临界游泳速度降低的现象。白斑红点鲑在16℃和20℃能量底物供给和抑制作用基本达到动态平衡, 因此, 此水温的临界游泳速度最高, 因此认为白斑红点鲑在水温16℃和20℃水体中的运动能力相对较强, 且体重较大的个体在相同水温下抵抗水流的运动能力强于体重较小的个体。
3.2 水温、流速和体重与白斑红点鲑呼吸的影响
鱼类机体生理功能的完成会受到氧气的摄入、运输、底物利用和代谢废物的清除等代谢能力方面的影响[14]。鱼类的耗氧率是重要的呼吸代谢反应参数, 通过分析水温和流速等因素与耗氧率的相互关系可以了解鱼类的代谢规律[21]。本研究在测定临界游泳速度的同时测定其实时游泳耗氧率, 即在特定水温下所能承受最大流速过程中的耗氧率。研究发现白斑红点鲑在适宜的相同条件下体重越大其耗氧率却越低, 这与皱纹盘鲍(Haliotis discus hannai)[22]、中华绒螯蟹(Eriocheir sinensis)[23]和管角螺(Hemifusus tuba)[24]等不同物种, 以及与圆口铜鱼(Coreius guichcnoti)[9]和罗非鱼[10]等同一物种的研究一致。白斑红点鲑游泳耗氧率最大值随着水温的升高而增高。在低温组的4℃和8℃时, 其游泳耗氧率均随着流速的增长出现不同幅度的增长, 但与高温组相比整体游泳耗氧率却偏低, 这可能与水温较低影响了有氧呼吸相关酶类活性和血液循环运输氧气能力导致[13, 23], 详细机理有待下一步更深入研究。在12℃和16℃其游泳耗氧率虽然随着流速增加不断增加, 但鱼力竭和鱼力竭前一档流速时其游泳耗氧率增加幅度明显降低, 甚至在20℃和24℃时随着流速不断增加, 当达到一定极限后出现了游泳耗氧率降低的现象, 且在24℃水体中游泳耗氧率降低更加明显。鱼类的这种代谢调整可能与其长期适应变化的环境条件有关[13, 19]。鱼类在一定水温下, 为保证游泳的持续性过早地启动无氧代谢, 由于水温较高使其机体内肌酸迅速积累, 且二氧化碳未能及时清除, 而阻碍了有氧代谢的进行导致了游泳耗氧率降低[25], 这与宋波澜等[26]对红鳍银鲫(Barbodes schwanenfeldi)、蔡露等[27]对齐口裂腹鱼(Schizothorax prenanti)和徐革锋等[13]对细鳞鲑的研究结果基本一致。因此, 本研究认为1龄、2龄和3龄组白斑红点鲑适宜的最高水温不应超过20℃, 1龄组最高流速不应超过32 cm/s, 2龄组最高流速不应超过40 cm/s, 3龄组最高流速不应超过48 cm/s。
[1] Ren M L. Ichthyofauna of the Heilongjiang River [J]. Chinese Journal of Fisheries, 1994, 7(1): 1—14 [任慕莲. 黑龙江鱼类区系. 水产学杂志, 1994, 7(1): 1—14]
[2] Zhang Y Q, Yin J S, Du J, et al. The artificial propagation of Salvelinus leucomaenis [J]. Journal of Anhui Agricultural University, 2011, 38(6): 902—906 [张永泉, 尹家胜, 杜佳, 等. 白斑红点鲑人工繁殖. 安徽农业大学学报, 2011, 38(6): 902—906]
[3] Zhang Y Q, Yin J S, Wang B Q, et al. Effects of stocking density on growth, survival and behavior of juvenile Siberian charr Salvelinus leucomaeni [J]. Journal of Dalian Fisheries University, 2009, 24(6): 520—524 [张永泉, 尹家胜, 王丙乾, 等. 养殖密度对白点鲑幼鱼生长、存活以及行为的影响. 大连水产学院学报, 2009, 24(6): 520—524]
[4] Grav J. Studies in animal locomotion I. The movement of fish with special reference to the eel [J]. Journal of Experimental Biology, 1933, 10: 88—104
[5] Shen X M, Zhao Q L. The effects of water temperature and do on the respiratory rates of immature Fugu obscure [J]. Chinese Journal of Ecology, 2001, 20(4): 13—15 [沈旭明,赵清良. 温度、溶解氧对暗纹东方鱼屯幼鱼呼吸频率的影响. 生态学杂志, 2001, 20(4) :13—15]
[6] Feng X, Chen P M, Qing C X. Advancement of the study on respiratory metabolism of fishery resources organism [J]. Guangdong Agricultural Sciences, 2012, (4): 92—95 [冯雪,陈丕茂, 秦传新. 渔业资源生物呼吸代谢研究进展. 广州农业科学, 2012, (4): 92—95]
[7] Yuan X, Li L P, Tu Z Y, et al. The effect of temperature on fatigue induced changes in the physiology and swimming ability of juvenile Aristichthys nobilis (Bighead carp) [J]. Acta Hydrobiologica Sinica, 2014, 38(3): 505—509 [袁喜,李丽萍, 涂志英, 等. 温度对鳙幼鱼疲劳引起的生理变化和游泳能力的影响研究. 水生生物学报, 2014, 38(3): 505—509]
[8] Claireaux G, Lagardère J P. Influence of temperature, oxygen and salinity on the metabolism of the European sea bass [J]. Journal of Sea Research, 1999, 42(2): 157—168
[9] Sun B Z, Huang H, Cao W X, et al. Studies on the oxygen consumption rate and asphyxiant point of Megalobrama pellegrini and Coreius guichcnoti [J]. Acta Hydrobiologica Sinica, 2010, 34(1): 89—93 [孙宝柱, 黄浩, 曹文宣, 等.厚颌鲂和圆口铜鱼耗氧率与窒息点的测定. 水生生物学报, 2010, 34(1): 89—93]
[10] Lei S J. Effect of salinity and body weight on the oxygen consumption of Oreochromis niloticus × O. mossambicus [J]. Chinese Journal of Applied Ecology, 2002, 13(6): 739—742[雷思佳. 盐度与体重对台湾红罗非鱼耗氧率的影响. 应用生态学报, 2002, 13(6): 739—742]
[11] Tu Z Y, Yuan X, Wang C F, et al. Swimming capability and activity metabolism of subadult schizothorax macropogon [J]. Acta Hydrobiologica Sinica, 2012, 36(4): 682—688 [涂志英,袁喜, 王从锋, 等. 亚成体巨须裂腹鱼游泳能力及活动代谢研究. 水生生物学报, 2012, 36(4): 682—688]
[12] Zeng L Q, Zhang Y G, Fu S J, et al. Effects of acute temperature change on resting oxygen consumption rate and critical swimming speed in juvenile southern catfish (Silurus meridionalis Chen) [J]. Acta Hydrobiologica Sinica, 2011, 35(2): 276—282 [曾令清, 张耀光, 付世建, 等. 双向急性变温对南方鲇幼鱼静止耗氧率和临界游泳速度的影响.水生生物学报, 2011, 35(2): 276—282]
[13] Xu G F, Wang Y Y, Han Y, et al. Effect of locomotion and feeding on metabolic mode of juvenile lenok, Brachymystax lenok (Pallas) under different water temperature conditions [J]. Chinese Journal of Applied Ecology, 2014, 25(4): 1156—1162 [徐革锋, 王裕玉, 韩英, 等. 不同温度条件下运动和摄食对细鳞鲑幼鱼代谢模式的影响. 应用生态学报, 2014, 25(4): 1156—1162]
[14] Zhang W, Cao Z D, Fu S J. Effect of dissolved oxygen level on metabolic mode in juvenile crucian carp [J]. Acta Ecologica Sinica, 2012, 32(18): 5806—5812 [张伟, 曹振东,付世建. 溶氧水平对鲫鱼代谢模式的影响. 生态学报, 2012, 32(18): 5806—5812]
[15] Wang P, Gui F K, Wu C W. Classification of fish swimming speed [J]. Journal of Fishery Sciences of China, 2010, 17(5): 1137—1146 [王萍, 桂福坤, 吴常文. 鱼类游泳速度分类方法的探讨. 中国水产科学, 2010, 17(5): 1137—1146]
[16] Brett J R, Hollands M, Alderdice D R. The effect of temperature on the cruising speed of young sockeye and
coho salmon [J]. Journal of the Fisheries Research Board of Canada, 1958, 15(4): 587—605
[17] Rebecca F, Jeffrey M L, Domine L C, et al. Critical swimming speeds of late-stage coral reef fish larvae: variation within species, among species and between locations [J]. Marine Biology, 2005, 147(5): 1201—1212
[18] Mateus C S, Quintella B R, Almeida P R. The critical swimming speed of Iberian barbel Barbus bocagei in relation to size and sex [J]. Journal of Fish Biology, 2008, 73(7): 1783—1789
[19] Farrell A P. Comparisons of swimming performance in rainbow trout using constant acceleration and critical swimming speed tests [J]. Journal of Fish Biology, 2008, 72(3): 693—710
[20] Boutilier R G, Ferguson R A, Henry R P, et al. Exhaustive exercise in the sea lamprey (Petromyzon marinus): Relationship between anaerobic metabolism and intracellular acid-base balance [J]. Journal of Experimental Biology, 1993, 178: 7l—88
[21] Yuan X, Tu Z Y, Han J C, et al. Effects of flow rate on swimming behavior and energy consumption of Schizothorax chongi [J]. Acta Hydrobiologica Sinica, 2012, 36(2): 270—275 [袁喜, 涂志英, 韩京成, 等. 流速对细鳞裂腹鱼游泳行为及能量消耗影响的研究. 水生生物学报, 2012, 36(2): 270—275]
[22] Bi Y B, Jiang S, Liu H Y, et al. Effect of temperature and weight on oxygen consumption rate andammonia excretion rate of Haliotis discus hannai [J]. Chinese Journal of Applied and Environmental Biology, 2000, 6(5): 444—446 [毕远溥,蒋双, 刘海映, 等. 温度、体重对皱纹盘鲍耗氧量和排氨量的影响. 应用与环境生物学报, 2000, 6(5): 444—446]
[23] Wen X B, Chen L Q, Ai C X, et al. Effects of temperature and body weight on fasting metabolism of the juvenile Eriocheir sinensis [J]. Acta Hydrobiologica Sinica, 2003, 27(4): 357—361 [温小波, 陈立侨, 艾春香, 等. 温度和体重对中华绒螯蟹幼蟹饥饿代谢的影响. 水生生物学报, 2003, 27(4): 357—361]
[24] Luo J, Liu C W, Li F, et al. Suffocation point and diurnal metabolism pattern of Hemifusus tuba (Gmelin) [J]. Chinese Journal of Applied Ecology, 2008, 19(9): 2092—2096 [罗杰,刘楚吾, 李锋, 等. 管角螺窒息点及昼夜代谢规律. 应用生态学报, 2008, 19(9): 2092—2096]
[25] Lee C G, Devlin R H, Farrell A P. Swimming performance, oxygen consumption and excess post-exercise oxygen consumption in adult transgenic and ocean-ranched coho salmon [J]. Journal of Fish Biology, 2003, 62: 753—766
[26] Song B L, Lin X T, Wang W J, et al. Effects of water velocities on rheotaxis behaviour and oxygen consumption rate of tinfoil barbs Barbodes schwanenfeldi [J]. Acta Zoologica Sinica, 2008, 54(4): 686—694 [宋波澜, 林小涛,王伟军, 等. 不同流速下红鳍银鲫趋流行为与耗氧率的变化. 动物学报, 2008, 54(4): 686—694]
[27] Cai L, Liu G Y, Rachel Taupier, et al. The recovery of swimming performance of juvenile Schizothorax prenanti after fatigue [J]. Acta Hydrobiologica Sinica, 2013, 37(6): 993—998 [蔡露, 刘国勇, Rachel Taupier, 等. 齐口裂腹鱼幼鱼疲劳后游泳特性恢复状况研究. 水生生物学报, 2013, 37(6): 993—998]
EFFECTS OF THE WATER TEMPERATURE AND THE WEIGHT ON THE CRITICAL SWIMMING SPEED AND OXYGEN CONSUMPTION RATE OF SALVELINUS LEUCOMAENIS
ZHANG Yong-Quan1, YIN Jia-Sheng1, XU Ge-Feng2, XI Qing-Kai1, MA Bo1and BAI Qing-Li1
(1. Heilongjiang River Fisheries Research Institute of Chinese Academy of Fishery Sciences, Harbin 150070, China; 2. College of Animal Science and Technology, Northeast Agricultural University, Harbin 150030, China)
In this study, we applied ecological methods to measure the critical swimming speed and the oxygen consumption rate of three age groups of Salvelinus leucomaenis (one-year old, two-year old and three-year old) at different water temperatures (4, 8, 12, 16, 20, 24 ) and℃ the flow velocity. The results showed that both the water temperature and the body weight had significant effects on the critical swimming speed (P<0.05), however, the interaction between the two factors did not (P>0.05). At the same water temperature, the critical swimming speed of Salvelinus leucomaenis increased along with the aging/increase in weight. At 4 , ℃ the critical swimming speed was the lowest, which were (21.6±1.06) cm/s, (22.93±0.61) cm/s and (30.27±1.29) cm/s for the three groups respectively. The critical swimming speed increased along with the rise of the water temperature in all groups, and reached the maximum at 16 , ℃ which were (39.6±0.80) cm/s, (46.80±0.80) cm/s and (53.73±1.22) cm/s for the three groups respectively. Then the speed slightly decreased as the water temperature rose to 20℃. However, there was no statistical difference in the speed at 16 and ℃ at 20℃ (P>0.05). When the water temperature further rose to 24 , ℃ the critical swimming speed dropped significantly. The water temperature, the flow velocity and the interaction between these two factors displayed significant effects on the oxygen consumption rate (P<0.05). Under the same suitable conditions, the oxygen consumption rate of Salvelinus leucomaenis significantly decreased as the weight increased. Overall, the oxygen consumption rates of the three groups were all increased with the rise of the water temperature and the flow rate, but the swimming oxygen consumption appeared to decrease when the water temperature and the flow rate reached a certain value. This study demonstrated that heavier individuals had higher swimming ability against the water flow given the same water temperature. The suitable water temperature for the three groups of Salvelinus leucomaenis should not exceed 20℃, and the maximum flow rate should be no higher than 32 cm/s, 40 cm/s and 48 cm/s for the three groups respectively.
Salvelinus leucomaenis; Water temperature; Flow velocity; Oxygen consumption rate; Critical swimming speed
S965.1
A
1000-3207(2015)04-0661-08
10.7541/2015.88
2014-08-18;
2015-02-12
国家科技支撑计划(2012BAD25B10); 公益性行业科研专项(201003055)资助
张永泉(1982—), 男, 蒙古族; 黑龙江哈尔滨人; 助理研究员; 硕士; 主要从事鱼类繁殖生物学和生态学研究。E-mail: atai0805@163.com
白庆利, 研究员, E-mail: bql2002sc@hotmail.com
猜你喜欢
当代水产(2021年9期)2021-12-02
基层中医药(2021年8期)2021-11-02
山东工艺美术学院学报(2020年5期)2020-12-10
水产科学(2018年3期)2018-06-04
山东工艺美术学院学报(2017年4期)2017-11-01
妈妈宝宝(2017年3期)2017-02-21
中国民族医药杂志(2016年1期)2016-05-09
工业设计(2016年11期)2016-04-16
工业设计(2016年8期)2016-04-16
河北渔业(2015年7期)2015-07-21
