
SAMP6小鼠骨髓间充质干细胞线粒体呼吸链复合体功能研究
2015-04-20 06:52胡成虎张少锋
牙体牙髓牙周病学杂志 2015年5期
杨 燚,胡成虎,张少锋,金 岩
(军事口腔医学国家重点实验室,陕西省口腔医学重点实验室,第四军医大学口腔医院:*修复科,**组织工程研究中心,陕西西安710032)
SAMP6小鼠骨髓间充质干细胞线粒体呼吸链复合体功能研究
杨 燚*,**,胡成虎**,张少锋*,金 岩**
(军事口腔医学国家重点实验室,陕西省口腔医学重点实验室,第四军医大学口腔医院:*修复科,**组织工程研究中心,陕西西安710032)
目的:研究自发性老年性骨质疏松模型SAMP6小鼠骨髓间充质干细胞(BMMSCs)的衰老状态、分化功能及线粒体呼吸链复合体活性。方法:分离培养SAMP6及其对照品系SAMR1小鼠BMMSCs,分别以实时定量PCR(RT-PCR)、Western Blot及细胞染色的方法,检测衰老、分化相关指标,并分别检测线粒体呼吸链复合体活性及复合体相关基因的表达。结果:与对照品系SAMR1小鼠相比,SAMP6小鼠BMMSCs的衰老相关基因、蛋白水平及β-gal染色阳性率显著升高(P<0.05);成骨分化相关基因、蛋白水平及茜素红染色阳性率显著降低(P<0.05);线粒体呼吸链复合体Ⅰ和Ⅲ活性显著降低,且呼吸链复合体相关基因表达亦降低(P<0.05)。结论:SAMP6小鼠BMMSCs存在细胞衰老及成骨分化功能障碍,其线粒体呼吸链复合体功能显著下降。
SAMP6小鼠;骨髓间充质干细胞(BMMSCs);线粒体呼吸链复合体
[Chinese Journal of Conservative Dentistry,2015,25(5):263]
骨髓间充质干细胞(bone marrow mesenchymal stem cells,BMMSCs)作为组织工程最重要的种子细胞,在口腔医学领域有着良好的应用前景。BMMSCs具有成骨、成脂、成软骨、成肌等多向分化潜能[1],且由于取材方便、易于分离培养、免疫原性低等优点,在干细胞治疗领域被视为最具有临床应用潜力的研究对象。目前研究中BMMSCs已经在包括心血管系统疾病、运动系统疾病、神经损伤乃至整形美容等多个领域展现良好的应用前景[2]。
自发性老年性骨质疏松模型小鼠(senescence accelerated mouse strain prone 6,SAMP6)以其早期出现骨密度降低、脆性增加、骨形成缺陷及全身性骨量减少等特性成为目前研究骨衰老及病理状态下BMMSCs性质的理想模型[3-4]。尽管越来越多的证据表明BMMSCs功能缺陷在骨质疏松发病过程起关键作用[5],目前尚缺乏对SAMP6及其对照品系SAMR1小鼠BMMSCs衰老状态及分化能力的系统比较,也鲜有对其功能缺陷机制的探索。
线粒体功能缺陷被认为是细胞衰老的重要标志之一[6],这其中又尤以呼吸链复合体功能的下降为重要因素。线粒体呼吸链是细胞活性氧(reactive oxygen species,ROS)的产生场所,而呼吸链复合体功能的下降直接导致ROS的产生增加,引发一系列级联反应最终导致细胞衰老及功能缺陷[7]。恢复线粒体功能也因此可能成为治疗年龄相关性疾病的新手段[8-9]。
本研究旨在以SAMP6骨早衰小鼠为模型,对其BMMSCs衰老状态及成骨分化能力进行检测鉴定,并以对线粒体呼吸链复合体功能的研究为入口,探讨造成BMMSCs功能下降,并最终引发骨形成缺陷的内在机制,为基于恢复干细胞线粒体功能的治疗方法提供依据,同时为进一步拓宽干细胞治疗的应用前景提供新的思路。
1 材料和方法
1.1 主要材料、试剂和仪器
SAMP6及SAMR1小鼠(第四军医大学实验动物中心提供);α-MEM培养液、链霉素、青霉素、L-谷氨酰胺(Gibco,美国);胎牛血清(杭州四季青);维生素C、β-磷酸甘油钠、地塞米松(Sigma,美国);兔抗P53、兔抗P21抗体(CST,美国);兔抗RUNX2、兔抗ALP、羊抗SP7抗体(Abcam,美国);鼠抗β-actin抗体、抗兔二抗、抗鼠二抗、抗羊二抗(上海康为世纪);Trizol Reagent(Life Technologies,美国);RNA逆转录试剂盒、SYBR-ⅡGreen荧光定量试剂(TAKARA,日本);相关基因引物(上海生工);PCR扩增仪(BioMetra,美国);Western及IP用细胞裂解液、BCA蛋白定量试剂盒、细胞β-gal原位染色试剂盒、线粒体呼吸链复合体(Ⅰ~Ⅳ)活性检测试剂盒(上海碧云天);PVDF膜(DUPONT,美国);电泳仪、转膜仪(BIO-RAD,美国);ECL发光试剂(上海七海生物);Tanon图像扫描分析系统(上海天能);YJ-875型超净工作台(苏州净化设备厂);倒置相差显微镜及照相系统(Olympus,日本);CO2恒温孵箱(Forma,美国)。
1.2 小鼠BMMSCs的分离、培养与成骨分化诱导及表型鉴定
1.2.1 小鼠BMMSCs的分离、培养与成骨分化诱导
分别取4~6个月龄的SAMP6及SAMR1小鼠,脱颈处死后于750 mL/L乙醇中浸泡消毒10~15 min;超净工作台内解剖、分离小鼠股骨、胫骨,剪去两端骨骺后,以1 mL注射器、α-MEM培养液(含200 mL/L胎牛血清、100 mg/mL链霉素及青霉素)快速冲出骨髓组织于9 cm培养皿中,小心吹打至无肉眼可见组织团块后,置于37℃、饱和湿度、50 mL/L CO2培养箱中培养;第3天换液以去除悬浮细胞,而后每3 d换液;待细胞贴壁生长至90%时,弃去培养液,PBS清洗后,以2.5 g/L胰酶、37℃消化3~5 min后,加入等体积α-MEM培养液终止消化,细胞悬液于800 r/min离心5 min后弃去上清、计数,以2.5×104/cm2密度进行接种传代。对于成骨分化诱导,待P1代BMMSCs生长至90%以上时,换用含50 mg/mL维生素C,3 g/L β-磷酸甘油钠及4 mg/L地塞米松的完全培养液,每3 d换液。
1.2.2 小鼠BMMSCs的表型鉴定
分别取P3代SAMP6及SAMR1小鼠BMMSCs,消化、离心后用PBS重悬,使细胞密度为3×106/mL,并以每管100μL分装至EP管中;按照抗体使用说明的推荐比例,分别加入适宜浓度的抗Sca-1抗体、抗CD29抗体、抗CD90抗体、抗CD34抗体及抗CD45抗体,并将每组不加抗体的空白管作为对照;4℃避光孵育40 min后,离心并弃去上清,PBS清洗细胞两遍,以200μL PBS重悬细胞,流式细胞仪检测相应抗体标记细胞阳性率。
1.3 SAMP6和SAMR1小鼠BMMSCs衰老相关指标的检测
1.3.1 RT-PCR检测SAMP6和SAMR1小鼠BMMSCs衰老相关基因表达水平
分别取生长至90%的P1代SAMP6和SAMR1小鼠BMMSCs,以Trizol试剂提取总RNA,以RNA逆转录试剂盒反应合成cDNA;以cDNA为模板、GAPDH为内参照,RT-PCR仪分别检测两组细胞P53及P21的mRNA表达水平。反应条件严格遵照产品说明,反应体系如表1,相关引物序列见表2。

表1 RT-PCR反应体系

表2 细胞衰老相关基因引物及其序列
1.3.2 Werstern Blot检测细胞衰老相关蛋白水平
分别取生长至90%的P1代SAMP6和SAMR1小鼠BMMSCs,以含蛋白酶抑制剂的蛋白裂解液裂解细胞,冰上超声裂解提取细胞总蛋白,以BCA蛋白定量试剂盒进行蛋白定量,每组取等量蛋白样本进行10%十二烷基硫酸钠-聚丙烯酰胺凝胶电泳分离蛋白,湿转法转于PVDF膜,50 g/L牛血清白蛋白封闭液室温封闭2 h;分别以抗P53、抗P21及抗β-actin抗体以产品说明推荐浓度4℃孵育过夜;以辣根过氧化物酶标记的兔、鼠二抗常温孵育2 h后,ECL发光试剂显色,Tanon图像分析系统进行分析和记录。
1.3.3 衰老相关β-半乳糖苷酶(β-gal)染色
分别取P1代SAMP6和SAMR1小鼠BMMSCs,按照产品说明,吸除细胞培养液,PBS洗涤1次,加入1 mLβ-gal染色固定液,室温固定15min;吸除固定液,PBS洗涤细胞3次后,每孔加入1 mL染色工作液,于无CO2孵箱中37℃避光孵育过夜,PBS清洗去除染色液终止染色,倒置相差显微镜观察并拍照。
1.4 SAMP6和SAMR1小鼠BMMSCs成骨分化能力检测
1.4.1 RT-PCR检测SAMP6和SAMR1小鼠BMMSCs成骨分化相关基因表达水平
分别取成骨诱导7 d的P1代SAMP6和 SAMR1小鼠BMMSCs,以RT-PCR方法检测成骨分化相关基因ALP、RUNX2、OCN及COL-1的mRNA表达水平。检测方法与反应体系同前,所用引物序列见表3。

表3 成骨分化相关基因引物及其序列
1.4.2 Werstern Blot检测细胞成骨分化相关蛋白水平
分别取成骨诱导7 d的P1代SAMP6和SAMR1小鼠BMMSCs,以Werstern Blot方法检测成骨分化相关基因ALP、RUNX2及SP7的蛋白表达水平。检测方法同前。
1.4.3 茜素红染色
称取14.41 g茜素红溶解于100 mL去离子水中,充分搅拌混匀后,调整pH值为4.1,过滤备用;分别取成骨诱导14 d的P1代SAMP6和SAMR1小鼠BMMSCs,PBS冲洗3次,100 g/L多聚甲醛固定40 min;PBS清洗2遍,用上述茜素红染液染色15 min,PBS洗净染色液,倒置相差显微镜观察、拍照并使用ImageJ 1.43u软件进行半定量分析,得到茜素红染色阳性面积率(%)并进行统计学比较。
1.5 SAMP6和SAMR1小鼠BMMSCs线粒体呼吸链复合体功能的检测
1.5.1 线粒体呼吸链复合体活性检测
以复合体Ⅰ活性检测为例,以蛋白裂解液分别提取P1代两组BMMSCs总蛋白样本并定量后,调整蛋白浓度为250μg/100μL;设定分光光度仪为波长340 nm,间隔1 min,读数3次;按照试剂盒产品说明分别测定各组样本的背景对照、样本总活性测定及样本非特异性活性,并计算样本特异性活性测定值。复合体Ⅱ~Ⅳ活性检测方法与Ⅰ类似。
1.5.2 线粒体呼吸链复合体相关基因表达水平检测
分别取P1代的SAMP6和SAMR1小鼠BMMSCs,RT-PCR方法检测呼吸链复合体相关基因ATP5a、NDUFS8、ND1、Cytb及COX1的mRNA表达水平。检测方法与反应体系同前,所用引物序列见表4。

表4 线粒体呼吸链复合体相关基因引物及其序列
1.6 统计学分析
2 结果
2.1 SAMP6和SAMR1小鼠BMMSCs表型鉴定
流式细胞仪对两组细胞的表面标记物鉴定结果显示,P3代SAMP6小鼠BMMSCs的Sca-1阳性率为90.5%、CD29为98.0%、CD90为97.2%;CD34阳性率为1.2%、CD45为0.9%(图1)。P3代SAMR1小鼠BMMSCs的Sca-1阳性率为84.1%、CD29为94.8%、CD90为96.7%;CD34为1.0%、CD45为0.8%(图2)。

图1 流式细胞仪检测SAMP6小鼠BMMSCs表面标记物

图2 流式细胞仪检测SAMR1小鼠BMMSCs表面标记物
2.2 SAMP6和SAMR1小鼠BMMSCs衰老相关指标的检测
2.2.1 两组细胞衰老相关因子蛋白表达水平检测
Western Blot检测结果显示,SAMP6组
BMMSCs的衰老相关因子P53及P21蛋白表达水平明显高于SAMR1组(图3)。

图3 两组BMMSCs衰老相关因子蛋白水平比较
2.2.2 两组细胞衰老相关因子mRNA表达水平检测
RT-PCR检测结果显示,SAMP6组BMMSCs的衰老相关因子P53及P21的mRNA表达水平均显著高于SAMR1组(P<0.05)(图4)。

图4 两组BMMSCs衰老相关因子mRNA表达水平比较(*与SAMR1相比,P<0.05)
2.2.3 两组细胞衰老相关β-gal染色结果
β-gal染色结果显示,SAMP6组BMMSCs染色阳性率明显高于SAMR1组(图5)。
2.3 SAMP6和SAMR1小鼠BMMSCs成骨能力的检测
2.3.1 两组细胞成骨分化相关蛋白水平的检测
Western Blot检测结果显示,SAMP6组BMMSCs的成骨分化相关ALP、RUNX2及SP7水平明显低于SAMR1组(图6)。
2.3.2 两组细胞成骨分化相关基因mRNA表达水平的检测
RT-PCR检测结果显示,SAMP6组BMMSCs的成骨分化相关基因ALP、RUNX2、OCN及COL-1的mRNA表达水平均显著低于SAMR1组(P<0.05)(图7)。

图5 两组BMMSCs衰老相关β-gal染色比较(×100)

图6 两组BMMSCs成骨分化相关基因蛋白表达水平比较
2.3.3 两组细胞茜素红染色结果比较

图7 两组BMMSCs成骨分化相关基因mRNA表达水平比较
茜素红染色可见,SAMP6组细胞染色阳性率明显低于SAMR1组,半定量分析示SAMP6组细胞茜素红染色阳性面积显著低于SAMR1组(P<0.05)(图8~9)。

图8 两组BMMSCs茜素红染色
2.4 SAMP6和SAMR1小鼠BMMSCs线粒体呼吸链复合体功能检测
2.4.1 两组细胞线粒体呼吸链复合体Ⅰ~Ⅳ活性的比较
对两组细胞线粒体呼吸链复合体活性的检测可见,SAMP6组BMMSCs的复合体Ⅰ及复合体Ⅲ活性显著低于SAMR1组(P<0.05),而对于复合体Ⅱ及复合体Ⅳ两组活性无明显差异(图10)。
2.4.2 两组细胞呼吸链复合体相关基因mRNA表达水平检测
RT-PCR检测结果显示,SAMP6组BMMSCs的呼吸链复合体相关基因ATP5a、NDUFS8、ND1、Cytb及COX1的mRNA表达水平均显著低于SAMR1组(P<0.05)(图11)。

图9 两组BMMSCs半定量分析(*P<0.05)
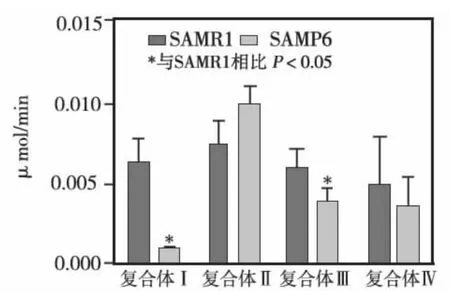
图10 两组BMMSCs 4种线粒体呼吸链复合体活性比较

图11 两组BMMSCs呼吸链复合体相关基因mRNA表达水平比较
3 讨论
自Friedenstein等首次(1976)以全骨髓贴壁法成功分离以来[10],BMMSCs以其良好的自我更新和多向分化潜能,较低的免疫原性及简便的取材方式而被视为理想的组织工程种子细胞受到广泛关注。不仅如此,基于BMMSCs的干细胞疗法也在多种疾病和损伤模型中展现出良好的应用前景。在口腔医学领域,BMMSCs在治疗牙和牙槽骨再生,以及颌面部缺损修复等方面的研究中占据重要地位[11],然而BMMSCs治疗要实现临床应用仍存在种种困难。由于BMMSCs在骨髓中存在比例低,取材后必须经过体外扩增方能获得足够的细胞数量,这一过程是否会对BMMSCs治疗活性产生影响尚不得而知。另一方面,自体干细胞移植疗法难以保证患者自身BMMSCs的质量,有学者认为在老年患者应用自体BMMSCs治疗心梗的过程中,由于患者BMMSCs处于衰老状态导致最终治疗效果欠佳,并以此作为解释BMMSCs治疗在临床试验中的效果不及在动物实验中出色的原因[12]。这些问题使得对BMMSCs的质量控制成为干细胞治疗应用中的新课题,也对病理状态下BMMSCs特性的研究提出了新的要求。
近年很多研究表明BMMSCs的衰老与分化异常与骨形成缺陷和骨质疏松发病之间存在密切联系。自发性老年性骨质疏松模型小鼠SAMP6在3~4个月龄即出现各种骨质疏松表型[13],这一性质使其成为研究病理状态下BMMSCs特性的良好模型。在本实验中,我们对SAMP6及其对照品系SAMR1小鼠的BMMSCs进行分离培养并进行细胞衰老和成骨分化能力的比较。其中,SAMP6组在细胞衰老相关因子p53及其下游p21的基因表达及蛋白水平上均显著高于SAMR1组,同时,衰老相关β-gal染色结果也说明了SAMP6组BMMSCs的衰老状态。另一方面,我们从成骨相关基因的表达、蛋白水平检测及茜素红染色等层次证实SAMP6组BMMSCs存在成骨分化能力缺陷。这些结果为BMMSCs参与骨质疏松发病过程的观点提供了依据,同时也提示恢复BMMSCs功能状态可能对于提高其治疗效果具有重要意义。
线粒体功能缺陷被认为是导致细胞衰老的始发因素之一,干细胞衰老和分化缺陷往往伴随着ROS升高、线粒体膜电位降低、线粒体形态学改变等一系列变化。而作为线粒体能量代谢工厂的核心,位于线粒体膜结构上的呼吸链则是造成这些功能变化的根源之一[14]。线粒体呼吸链由4种呼吸链复合体(Ⅰ~Ⅳ)及ATP合酶(也称复合体Ⅴ)共同协调完成电子传递及ATP合成过程。本实验通过对SAMP6和SAMR1小鼠BMMCSs呼吸链复合体功能的比较,发现SAMP6小鼠BMMCSs的呼吸链复合体Ⅰ及Ⅲ的活性显著低于SAMR1组。同时,我们检测了两组细胞中几种代表性的呼吸链复合体相关基因的表达状态,其中包括核编码基因NDUFS8和ATP5a,以及线粒体编码基因ND1、Cytb和COX1。SAMP6小鼠BMMSCs中这几种基因的表达均显著低于SAMR1组,提示其可能存在整体转录水平上的呼吸链复合体基因表达异常,这一异常在核编码及线粒体编码的基因中均存在,以此导致呼吸链复合体相关蛋白合成出现障碍,从而直接导致其活性的下降,引发线粒体能量合成功能障碍及ROS升高,最终导致BMMCSs衰老及分化功能缺陷。
线粒体呼吸链相关基因的表达需要多种因子协同参与共同维持。本实验表明衰老BMMSCs中存在线粒体呼吸链复合体相关基因的表达异常,但其具体发生机制仍需探索。例如,过氧化物酶体增殖物激活受体γ辅助激活因子1α(peroxisome proliferator-activated receptorγco-activator 1α,PGC-1α)通过协调其下游NRF1、2及ERRα等多种线粒体呼吸链复合体相关基因转录因子的表达调控复合体相关基因表达[15];又如核编码的线粒体转录因子A(mitochondria transcriptional factor A,TFAM)直接调控线粒体编码的复合体相关基因的表达[16]。以上这些因子的异常变化均可导致线粒体呼吸链相关基因表达的异常,从而导致复合体活性下降,因此也可能成为药物干预调控线粒体功能的靶点。本实验为改善线粒体因呼吸链异常导致的功能障碍,从而为提高BMMSCs的功能和治疗效果提供了新的思路。
[1]Pittenger MF,Mackay AM,Beck SC,etal.Multilineage potential ofadulthumanmesenchymal stem cells[J].Science,1999,284(5411):143-147.
[2] 王彤.骨髓间充质干细胞临床研究进展[M].北京:人民卫生出版社,2009.
[3]Matsushita M,Tsuboyama T,KasaiR,etal.Age-related changes in bonemass in the senescence-accelerated mouse(SAM).SAM-R/3 and SAM-P/6 as new murinemodels for senile osteoporosis[J].Am JPathol,1986,125(2):276-283.
[4]Silva MJ,BrodtMB,Ettner SL.Long bones from the senescence accelerated mouse SAMP6 have increased size but reduced whole-bone strength and resistance to fracture[J].J Bone Miner Res,2002,17(9):1597-1603.
[5]Chan GK,Duque G.Age-related bone loss:Old bone,new facts[J].Gerontology,2002,48(2):62-71.
[6]López-Otín,Blasco MA,Partridge L,et al.The hallmarks of aging[J].Cell,2013,153(6):1194-1217.
[7]Guarente L.Mitochondria-a nexus for aging,calorie restriction,and sirtuins[J].Cell,2008,132(2):171-176.
[8]Scheibye-Knudsen M,Fang EF,Croteau DL,et al.Protecting themitochondrial powerhouse[J].Trends Cell Biol,2015,25(3):158-170.
[9]Price NL,Gomes AP,Ling AJ,et al.SIRT1 is required for AMPK activation and the beneficial effects of resveratrol onmitochondrial function[J].Cell Metab,2012,15(5):675-690.
[10]Friedenstein AJ,Gorskaja JF,Kulagina NN.Fibroblast precursors in normal and irradiated mouse hematopoietic organs[J].Exp Hematol,1976,4(5):267-274.
[11] 陈发明,金岩,施松涛,等.转化医学:十年回顾与展望[J].实用口腔医学杂志,2011,27(1):1-7.
[12]Liang H,Hou H,YiW,et al.Increased expression of pigment epithelium-derived factor in aged mesenchymal stem cells impairs their therapeutic efficacy for attenuatingmyocardial infarction injury[J].Eur Heart J,2013,34(22):1681-1690.
[13]Jilka RL,Weinstein RS,Takahashi K,et al.Linkage of decreased bonemass with impaired osteoblastogenesis in amurine model of accelerated senescence[J].J Clin Invest,1996,97(7):1732-1740.
[14]Ziegler DV,Wiley CD,Velarde MC.Mitochondrial effectors of cellular senescence:beyond the free radical theory of aging[J].Aging Cell,2015,14(1):1-7.
[15]Anderson R,Prolla T.PGC-1αin aging and anti-aging interventions[J].Biochim BiophysActa,2009,1790(10):1059-1066.
[16]Finly LW,Haigis MC.The coordination of nuclear and mitochondrial communication during aging and calorie restriction[J].Ageing Res Rev,2009,8(3):173-188.
Function ofm itochondrial respiration chain comp lex in bone marrow mesenchymal stem cells of SAMP6 m ice
YANG Yi,HU Cheng-Hu,ZHANG Shao-Feng,JIN Yan
(State Key Laboratory of Military Stomatology,Department of prosthodontics,School of Stomatology,The Fourth Military Medical University,Shaanxi Key Laboratory of Stomatology,Xi′an 710032,China)
AIM:To detect senescence condition,osteogenic differentiation ability and the activity of mitochondrial respiration chain complex of bone marrow mesenchymal stem cells(BMMSCs)from senescence accelerated mouse strain prone 6(SAMP6).METHODS:BMMSCs of SAMP6 and its control strain SAMR1 mice were isolated and cultured.The cell senescence and osteogenic differentiation ability were detected by RT-PCR,Western Blot and cell staining respectively.The function ofmitochondrial respiration complex was examined by activity assay and related gene expression assay.RESULTS:Compared with control strain SAMR1,BMMSCs of SAMP6 performed increase of cellular senescence related genes and protein level,increase ofβ-gal positive cell ratio(P<0.05),decline of osteogenic function(P<0.05),and low activity of respiration chain complex.CONCLUSION:BMMSCs from SAMP6 perform senescence and osteogenic differentiation dysfunction with significant decline ofmitochondrial respiration chain complex.
SAMP6 mice;bonemarrow mesenchymal stem cells(BMMSCs);mitochondrial respiration chain
R780.2
A
1005-2593(2015)05-0263-07
10.15956/j.cnki.chin.j.conserv.dent.2015.05.002
2015-02-13;
:2015-03-24
国家自然科学基金资助项目(31301062)
杨燚(1989-),男,汉族,陕西西安人。博士生(导师:张少锋、金岩)
张少锋,E-mail:sfzhang@fmmu.edu.cn金 岩,E-mail:yanjin@fmmu.edu.cn
猜你喜欢
医学研究生学报(2022年5期)2022-12-07
中日友好医院学报(2022年1期)2022-11-28
临床肺科杂志(2022年3期)2022-11-26
农业工程学报(2022年13期)2022-10-09
中华实用诊断与治疗杂志(2022年1期)2022-08-31
大气科学学报(2022年2期)2022-05-12
实用口腔医学杂志(2022年2期)2022-05-05
聊城大学学报(自然科学版)(2022年3期)2022-02-14
作物学报(2022年3期)2022-01-22
中国卒中杂志(2021年7期)2021-11-29
