
牙髓干细胞向成牙本质细胞诱导分化过程中Oct4可变剪接体表达改变的研究
2015-04-20 06:52关丽娜孙雪飞张亚庆
牙体牙髓牙周病学杂志 2015年5期
贺 莹,关丽娜,孙雪飞,韩 冰,张亚庆,杨 帆
(军事口腔医学国家重点实验室,陕西省口腔医学重点实验室,第四军医大学口腔医院牙体牙髓病科,陕西西安710032)
牙髓干细胞向成牙本质细胞诱导分化过程中Oct4可变剪接体表达改变的研究
贺 莹,关丽娜,孙雪飞,韩 冰,张亚庆,杨 帆
(军事口腔医学国家重点实验室,陕西省口腔医学重点实验室,第四军医大学口腔医院牙体牙髓病科,陕西西安710032)
目的:研究多能性转录因子Oct4可变剪接体在人牙髓干细胞(hDPSCs)向成牙本质细胞定向分化过程中的表达改变。方法:矿化液诱导hDPSCs向成牙本质细胞分化,RT-PCR检测未经分化诱导、分化诱导7、14 d时Oct4可变剪接体(Oct4A,Oct4B)以及干性分子Sox2、Klf4和DSPP的表达改变;激光共聚焦显微镜(CLSM)检测Oct4A在分化过程中的表达和定位。结果:hDPSCs高表达CD146、CD105、CD90、CD29;CD45及CD34表达阴性。矿化液诱导细胞分化后牙本质涎磷蛋白(DSPP)的总蛋白和mRNA表达阳性。在hDPSCs未经分化诱导、分化诱导7、14 d时,Oct4A和Sox2、Klf4均有表达,分化诱导后表达量明显降低(P<0.05),14 d时hDPSCs表达均低于7 d(P<0.05);Oct4B在hDPSCs未经分化诱导、分化诱导7、14 d时表达均为阴性;TIP110与Oct4A在hDPSCs成牙本质分化过程中表达趋势相同。未经分化诱导hDPSCs剪接体Oct4A主要表达于细胞核,而分化诱导21 d后主要表达于细胞质。结论:hDPSCs向成牙本质细胞分化过程中伴随着其干性降低、剪接体Oct4A表达降低和核浆穿梭效应的发生,提示Oct4A在维持hDPSCs干性中发挥着一定作用。
牙髓干细胞;Oct4;成牙本质细胞分化;可变剪接体
[Chinese Journal of Conservative Dentistry,2015,25(5):282]
多能性转录因子Oct4(0ctamer binding transcription factor-4)是第一个被确定为调控干细胞多潜能性的核心转录因子,在维持胚胎干细胞未分化状态和体细胞重编程中发挥着重要的作用[1],已证实Oct4基因敲除小鼠缺失具有多向分化潜力的细胞群[2]。此外,在早期胚胎发育中Oct4也是决定细胞特定分化方向的重要因子。小鼠胚胎干细胞内抑制Oct4基因表达,细胞失去原有向内细胞团分化能力,分化形成滋养层[3]。进一步的研究发现抑制Oct4表达可促进中胚层发育,相反高表达则促进内胚层发育[4]。Oct4经选择性剪接可生成2种主要剪接异构体Oct4A和Oct4B,二者组成不同,功能也不尽相同。Oct4A mRNA序列包括447个碱基对,而Oct4B mRNA序列包括344个碱基。Oct4A是Oct4的原始形式,主要表达于未分化细胞的细胞核中,参与维持干细胞未分化状态。而Oct4BmRNA序列由344个碱基构成,表达于非多能性细胞[4-5],其作用尚不清楚。目前有关Oct4不同剪接体产生的机制尚未明确,有研究发现TIP110通过调控Oct4选择性剪接参与维持胚胎干细胞Oct4A高表达[6]。牙髓干细胞(dental pulp stem cells,DPSCs)属于成体间充质干细胞,具有多向分化和自我更新能力,位于牙髓组织血管旁的细胞龛内。当牙髓受到损伤时,DPSCs从龛内迁徙至受损部位分化为特定类型细胞参与牙髓损伤修复。DPSCs在未分化与分化状态的转变受到自身携带的遗传信息以及周围所处的微环境的调控[7],对比分化前后基因表达改变有助于筛选出维持细胞未分化状态和诱导其定向分化的调控因子。Oct4作为转录因子一方面参与维持细胞未分化状态,一方面抑制分化相关特定基因表达。研究发现在人牙髓干细胞(hDPSCs)内可检测到Oct4、Sox2、Klf4高表达,且表达随着DPSCs的衰老降低[8],说明Oct4在hDPSCs的干性维持中发挥重要作用。本实验拟采用实时荧光定量聚合酶链式反应(qPCR)和激光共聚焦显微镜检测牙髓干细胞向成牙本质细胞诱导分化前后Oct4主要可变剪接体(Oct4A和Oct4B)的表达水平和定位改变,为进一步深入研究Oct4在hDPSCs干性维持中的作用奠定基础。
1 材料和方法
1.1 材料和器材
α-MEM培养基、胎牛血清、胰蛋白酶(Hyclone,美国);双抗(Gibco,美国);Dispase酶(Roche,瑞士);CD29-PE、CD34-PE、CD45-PE、CD90-PE、CD105-PE、CD146-PE(Biolegend,美国);鼠抗人Oct4A单克隆抗体(Santa Cruz,Cat sc-5279,美国);山羊抗鼠Cy3荧光二抗(武汉博士德公司);Ⅰ型胶原酶、β-甘油磷酸钠、地塞米松、维生素C、离心机、小鼠抗β-actin抗体(Sigma,美国);小鼠抗人DSPP抗体(Santa Cruz,sc-73632,美国);Trizol Reagent、Oct4A、Sox2、Klf4、TIP110及β-actin引物(Invitrogen,美国);细胞孵箱(Heraeus,德国);倒置相差显微镜(Olympus,日本);低温高速离心机(Eppendorf,德国);cDNA合成试剂盒、SYBR Green试剂盒(Takara,日本)。
1.2 人牙髓细胞的分离培养和纯化
临床收集18~22岁新鲜拔除的健康完整的第三磨牙和因正畸需要拔除的牙齿,用带有双抗的PBS于超净台冲洗净后,置于无菌环境中劈开取出牙髓,用含有双抗的PBS反复冲洗3次,剪成1 mm ×1 mm×1 mm小块,移入含有3 g/L胶原酶和4 g/L Dispase酶(1∶1)离心管内,联合消化法37℃消化45 min,待组织块呈絮状加入等体积含双抗的100 mL/L胎牛血清的α-MEM培养液终止消化,1 000 r/min离心5 min,弃上清液,细胞培养液重悬2次后加入200 mL/L胎牛血清的α-MEM培养基,接种于35 mm的培养皿,37℃、50 mL/L CO2孵箱培养3 d,倒置显微镜下观察组织块周围是否有细胞爬出,采用有限稀释法挑选单克隆并传代扩增,取处于生长对数期的第4代细胞进行研究。
1.3 牙髓干细胞的鉴定
1.3.1 流式细胞仪鉴定干细胞表面标记物
2.5 g/L胰蛋白酶消化P4 hDPSCs,PBS清洗两遍,显微镜下计数,调整细胞浓度为1×109/L,分别用CD146、CD105、CD90、CD45、CD34和CD29标记抗体,室温避光孵育30 min,PBS清洗3遍,流式细胞仪进行检测。
1.3.2 矿化诱导培养
hDPSCs(1×105/孔)接种于35 mm培养皿,等细胞融合至80%时更换培养液为矿化诱导液(含有100 mL/L FBS的α-MEM培养基中加入终浓度为2 ng/mL地塞米松、10 mmol/Lβ-甘油磷酸钠、50 ng/mL维生素C),每3 d换液1次,诱导分化14 d终止矿化培养,茜素红进行染色。
1.3.3 成脂诱导培养
hDPSCs(1×105/孔)接种于35 mm培养皿,等细胞融合至80%时更换培养液为成脂诱导液(含有100 mL/L FBS的α-MEM培养基中加入终浓度为0.2 mmol/L吲哚美辛、0.5 mmol/L IBMX、10 mg/L胰岛素、1 mmol/L地塞米松),每3 d换液1次,21 d后终止培养,油红O染色。
1.4 牙本质涎磷蛋白(DSPP)的表达检测
Western blot:矿化诱导液连续培养hDPSCs 2周,分别提取矿化0、7、14 d时总蛋白,Western blot检测DSPP的表达,以β-actin作为内参。
1.5 实时荧光定量PCR(RT-PCR)
收集未经分化诱导和分化诱导7、14 d的牙髓干细胞,分别用Trizol试剂盒提取总RNA,分光光度计测量总RNA浓度,使用Takara逆转录试剂盒将1μg RNA反转录为cDNA模版,使用20μL体系进行PCR扩增反应,引物设计和合成由Invitrogen公司完成,引物序列见表1。PCR扩增40个循环,包括10 min预变性,95℃变性,55℃退火,72℃延伸及最后10 min延伸。采用2-△△Ct法进行数据分析。
1.6 免疫荧光染色
将P4代hDPSCs以1×103/孔的密度接种于24孔板内的盖玻片上,加入矿化诱导液,21 d后终止培养,PBS清洗5 min×3次,40 g/L多聚甲醛固定30 min,PBS清洗5 min×3次,1 g/L Triton X-100通透20 min,PBS清洗3次,山羊来源的血清封闭30 min,孵一抗(1∶150),4℃湿盒过夜,PBS清洗3次,避光孵二抗2 h,PBS清洗,DAPI染细胞核,激光共聚焦显微镜下观察拍照。

表1 RT-PCR引物序列
1.7 统计学分析
采用SPSS 17.0统计软件对实验结果进行单因素方差分析,两两比较采用t检验,检验水准α=0.05。
2 结果
2.1 人牙髓干细胞的分离培养和鉴定
镜下见组织块贴壁3 d后周围有梭形细胞爬出(图1),贴壁细胞胞体小,胞核大。换液继续培养至10 d左右细胞融合达80%时进行有限稀释法挑选单克隆,经单克隆细胞筛选扩增后的细胞经流式细胞仪检测表面标记物,结果显示,CD29、CD90、CD105、CD146呈阳性表达,CD34、CD45阴性表达(图2)。分化诱导14 d后,细胞呈复层生长,出现颗粒状结节,茜素红染色阳性(图3a),镜下见大量着色的矿化结节(图3b)。细胞成脂诱导21 d,油红O染色显示胞浆内出现散在脂滴(图3c)。

图1 组织块周围细胞爬出情况(倒置显微镜,×10)
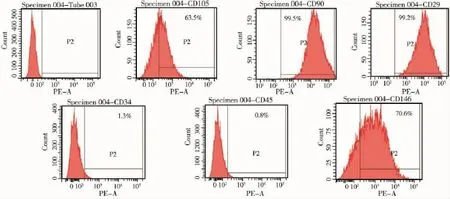
图2 流式细胞仪鉴定干细胞表面标记物
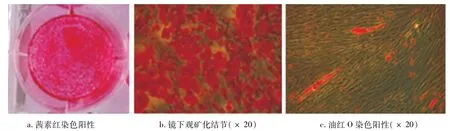
图3 hDPSCs分化能力鉴定
2.2 DPSCs、成牙本质细胞分化过程中Oct4可变剪接体及其调控分子TIP110、干性分子(Sox2和Klf4)表达改变
Western blot结果显示未经矿化诱导的人牙髓干细胞不表达DSPP,经矿化诱导7、14 d后,牙髓干细胞高表达DSPP,且随诱导时间增加表达增强(图4A)。qPCR结果显示Oct4A和干性分子(Sox2,Klf4)在牙髓干细胞未经分化诱导,分化诱导7、14 d均有表达,而Oct4B表达阴性,Oct4A、Sox2和Klf4随诱导时间增加表达量明显降低(P<0.05),诱导14 d表达低于7 d(P<0.05);Oct4可变剪接调控分子TIP110与Oct4A在牙髓干细胞成牙本质分化诱导过程中表达趋势相同,在牙髓干细胞未经分化诱导,分化诱导7、14 d均有表达,且随诱导时间增加表达量明显降低(P<0.05)(图4B)。
2.3 DPSCs成牙本质分化诱导前后Oct4A表达定位
激光共聚焦显微镜下观察发现未经分化诱导的hDPSC Oct4A主要位于细胞核,hDPSC成牙本质向诱导分化21 d时Oct4A表达主要位于细胞质(图5)。

图4 hDPSCs经矿化诱导分化前后相关基因的表达

图5 Oct4A免疫荧光染色结果(×60)
3 讨论
转录因子Oct4可与Sox2、Klf4协同共同维持干细胞多能状态,当Oct4表达受抑制,干细胞干性相关基因表达下调,细胞发生分化[9]。Oct4选择性剪接形成Oct4A、Oct4B两种主要可变剪接体,但研究显示只有剪接体Oct4A参与维持干细胞的全能性[10]。本研究着重观察DPSCs向成牙本质细胞分化过程中转录因子Oct4可变剪接体表达改变,以初步探究Oct4可变剪接体在DPSCs干性维持中作用。本结果表明DPSCs向成牙本质细胞分化过程中伴随着成牙本质细胞特异标记物DSPP表达增高,细胞其干性基因表达明显降低,同时在未经分化诱导DPSCs检测到Oct4A剪接体,且诱导分化后表达降低,并发生胞核向胞浆转位,但剪接体Oct4B在诱导DPSCs分化前后均不表达。
Oct4两种主要可变剪接体不同的表达分布提示二者功能不同。本研究发现,体外培养的DPSCs表达Oct4A,但不表达Oct4B,在诱导DPSCs成牙本质细胞分化过程中伴随干性分子Sox2和Klf4表达降低,Oct4A的表达也明显降低,表明Oct4可变剪接体中Oct4A是真正发挥调控DPSCs干性维持的分子。Wang等报道,Oct4B对应急状态的细胞具有保护作用[11]。有研究发现炎症牙髓组织中Oct4BmRNA水平显著高于正常牙髓组织[12],提示可能与牙髓组织的免疫应答有关。
基因mRNA水平上的可变剪接是指从一个mRNA前体中通过不同的剪接方式(选择不同的剪接位点组合)产生不同的mRNA剪接异构体的过程。研究显示90%的人类基因会在选择性剪接的作用下产生功能相同、不同甚至拮抗的多个同源异构体(isoform),从而增加蛋白质的多样性。已证实单个基因选择性剪接模式的异常即可引起细胞表性改变,发育异常和疾病。有研究提示Ⅱ型牙本质发育不全症(DGI-Ⅱ)的发生与牙本质涎磷蛋白选择性剪接位点基因点突变相关,提示维持正常选择性剪接模式是牙齿发育的重要因素[13]。Liu等报道TIP110调控Oct4的选择性剪接,过表达TIP110可上调Oct4A的表达量,抑制TIP110表达则不能形成剪接体Oct4A,但并不影响剪接体Oct4B的形成[6]。本研究也发现TIP110在DPSCs经成牙本质细胞诱导分化中的动态表达变化与Oct4A的表达一致,提示在DPSCs内TIP110可能参与调控Oct4可变剪接过程中Oct4A表达,但机制仍需进一步的实验证实。
转录因子的亚细胞定位是调控核蛋白活性的重要机制。文献报道,多种胞核分子信号通路参与调控核蛋白胞浆转位[14-15]。Oct4A作为转录因子主要位于未分化的细胞核内以维持干细胞的全能性[16]。本实验荧光染色结果显示在未经分化诱导DPSCs中Oct4A主要表达在胞核,但矿化诱导21 d后表达主要位于细胞质,发生胞核向胞质的转位,也进一步提示Oct4A在维持DPSCs未分化状态中具有重要作用。有研究发现特定激酶介导的磷酸化调控着转录因子核浆穿梭的过程。Ferro等发现在DPSCs中丝/苏氨酸蛋白激酶CK-II介导的Oct4A蛋白磷酸化程度减少是导致Oct4A发生胞核向胞浆转位的主要原因[9]。Spelat等发现Oct4A含有6个特定的ERK磷酸化结合位点,ERK通过磷酸化Ser111位点增强Oct4A从胞核到胞质转位[17]。
DPSCs干性维持是由多种基因、蛋白和外界因素共同决定的。本实验初步证实DPSCs内Oct4A通过其表达量改变和核浆穿梭效应参与调控维持细胞未分化状态,但其调控机制仍需进一步深入研究。
[1]Nichols J,Zevnik B,Anastassiadis K,etal.Formation of pluripotent stem cells in themammalian embryo depends on the POU transcription factor Oct4[J].Cell,1998,95(3):379-391.
[2]Reim G,Mizoguchi T,Stainier D Y,et al.The POU domain protein spg(pou2/Oct4)is essential for endoderm formation in cooperation with the HMG domain protein casanova[J].Dev cell,2004,6(1):91-101.
[3]Niwa H,Miyazaki J,Smith AG.Quantitative expression of Oct-3/4 defines differentiation,dedifferentiation or self-renewal of ES cells[J].Nat Genet,2000,24(4):372-376.
[4]Tsai SC,Chang DF,Hong CM,etal.Induced overexpression of OCT4A in human embryonic stem cells increases cloning efficiency[J].Am JPhysiol Cell Physiol,2014,306(12):1108-1118.
[5]Lee J,Kim HK,Rho JY,etal.The human OCT-4 isoforms differ in their ability to confer self-renewal[J].J Biol Chem,2006,281(44):33554-33565.
[6]Liu Y,Timani K,Ou X,etal.C-MYC controlled TIP110 protein expression regulates OCT4 mRNA splicing in human embryonic stem cells[J].Stem cells Dev,2012,22(5):689-694.
[7]Huang GT,Yamaza T,Shea LD,et al.Stem/progenitor cellmediated de novo regeneration of dental pulp with newly deposited continuous layer of dentin in an in vivo model[J].Tissue Eng Part A,2009,16(2):605-615.
[8]Ferro F,Spelat R,D'Aurizio F,et al.Dental pulp stem cells differentiation reveals new insights in Oct4A dynamics[J].PloS One,2012,7(7):e41774.
[9]Ding J,Xu H,Faiola F,et al.Oct4 linksmultiple epigenetic pathways to the pluripotency network[J].Cell Res,2012,22(1):155-167.
[10]Seo KW,Lee SR,Bhandari DR,et al.OCT4A contributes to the stemness and multi-potency of human umbilical cord bloodderived multipotent stem cells(hUCB-MSCs)[J].Biochem Biophys Res Commun,2009,384(1):120-125.
[11]Wang X,Zhao Y,Xiao Z,etal.Alternative translation ofOCT4 by an internal ribosome entry site and its novel function in stress response[J].Stem Cells,2009,27(6):1265-1275.
[12]韦曦,孔倩颖,黄奕俊,等.正常及炎症状态人牙髓中八聚体结合转录因子4B的表达[J].中华口腔医学研究杂志(电子版),2012,6(2):7-10.
[13]Wang SK,Chan HC,Makovey I,et al.Novel PAX9 and COL1A2 missensemutations causing tooth agenesis and OI/DGI without skeletal abnormalities[J].PloS One,2012,7(12):e51533.
[14]Radzisheuskaya A,Silva JC.Do all roads lead to Oct4?The emerging concepts of induced pluripotency[J].Trends Cell Biol,2014,24(5):275-284.
[15]Oka M,Moriyama T,Asally M,et al.Differential role for transcription factor Oct4 nucleocytoplasmic dynamics in somatic cell reprogramming and self-renewal of embryonic stem cells[J].J Biol Chem,2013,288(21):15085-10097
[16]Xu L,MassaguéJ.Nucleocytoplasmic shuttling of signal transducers[J].Nat Rev Mol Cell Biol,2004,5(3):209-219.
[17]Spelat R,Ferro F,Curcio F.Serine 111 phosphorylation regulates OCT4A protein subcellular distribution and degradation[J].JBiol Chem,2012,287(45):38279-38288.
Expressions of Oct4 isoforms during odontoblastic differentiation of human dental pulp stem cells
HE Ying,GUAN Li-na,SUN Xue-fei,HAN Bing,ZHANG Ya-qing,YANG Fan
(State Key Laboratory of Military Stomatology,Department of Operative Dentistry and Endodotics,School of Stomatology,The Fourth Military Medical University,Shaanxi Key Laboratory of Stomatology,Xi′an 710032,China)
AIM:To examine the expression of Oct4 isoforms during odontoblastic differentiation of human dental pulp stem cells(hDPSCs).METHODS:hDPSCswere cultured in odontogenic inductionmedium.Expression of Oct4 isoforms(Oct4A and Oct4B),stem cellsmarkers Sox2,Klf4 and DSPPwere detected by RT-PCR.The protein location of Oct4A in hDPSC was detected by confocal laser scanningmicroscope(CLSM).RESULTS:hDPSCs positively expressed CD146,CD105,CD90 and CD29,but negatively expressed CD45 and CD34.Oct4A isoform and Sox2 and Klf4 were downregulated after odontogenic induction(P<0.05),while Oct4B isoform was not detected before and after induction of hDPSCs.TIP110 showed the same dynamic changeswith Oct4A during odontoblastic differentiation.Moreover,Oct4A was translocated from nuclear to cytoplasm after odontogenic induction.CONCLUSION:The stemness of hDPSC are reduced during odontogenic induction.Downregulation and translocation of spliced variant Oct4A indicates that itmay play a role inmaintaining stemness of hDPSC.
dental pulp stem cell;Oct4;odontoblastic induction;alternative splicing variant
R780.2
A
1005-2593(2015)05-0282-06
10.15956/j.cnki.chin.j.conserv.dent.2015.05.005
2014-12-20;
:2015-03-30
国家自然科学基金资助项目(81070803,81170930)
陕西省国际合作课题(2011KW-40)
贺莹(1989-),女,汉族,陕西榆林人。硕士生(导师:张亚庆)
张亚庆,E-mail:Zhangyaqq@fmmu.edu.cn
杨 帆,E-mail:yangfan@fmmu.edu.cnn
猜你喜欢
康复(2022年27期)2022-11-18
中国典型病例大全(2022年11期)2022-05-13
中国典型病例大全(2022年10期)2022-05-10
锦州医科大学报(2022年2期)2022-05-07
现代临床医学(2021年3期)2021-07-16
现代口腔医学杂志(2021年1期)2021-03-12
方圆(2017年12期)2017-07-17
中国医药导报(2016年29期)2016-12-27
家庭医学(2015年8期)2015-09-10
华西口腔医学杂志(2013年5期)2013-11-11
